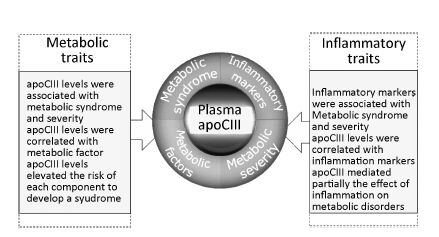
Objective Assessment of the comprehensive relationship among apolipoprotein CIII (apoCIII) levels, inflammation, and metabolic disorders is rare.Methods A total of 1455 consecutive patients not treated with lipid-lowering drugs and undergoing coronary angiography were enrolled in this cross-sectional study. A mediation analysis was used to detect the underlying role of apoCIII in the association of inflammation with metabolic syndrome (MetS).Results Patients with MetS showed higher levels of apoCIII[95.1 (73.1-131.4) vs. 81.7 (58.6-112.4) μg/mL, P<0.001] and inflammatory markers[high sensitivity C-reactive protein, 1.7 (0.8-3.4) vs. 1.1 (0.5-2.2) mg/L; white blood cell count, (6.48±1.68) vs. (6.11±1.67)×109/L]. The levels of apoCIII and inflammatory markers increased with the number of metabolic risk components (all P<0.001). Furthermore, apoCIII levels were associated with virtually all individual MetS risk factors and inflammatory markers (all P<0.05). Importantly, the prevalence of MetS in each metabolic disorder rose as apoCIII levels increased (all P<0.05). Mediation analysis showed that apoCIII partially mediated the effect of inflammation on MetS independently from triglycerides.Conclusion Plasma apoCIII levels were significantly associated with the development and severity of MetS, and a role of apoCIII in the effect of inflammation on the development of MetS was identified.
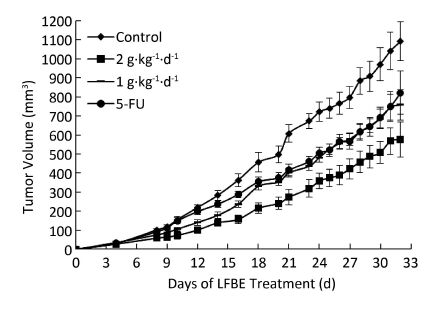
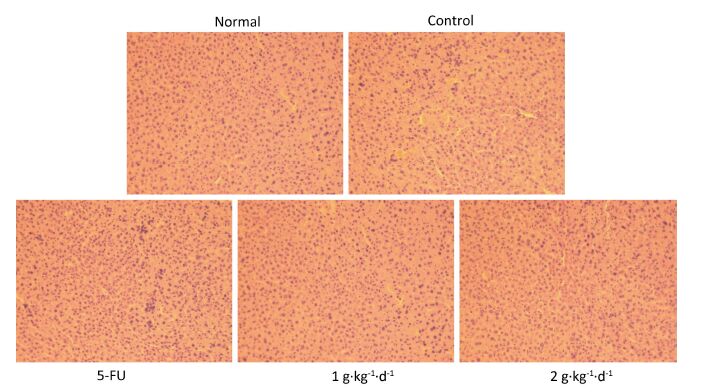
Objective A subcutaneous transplantation tumor model of human HT-29 cells was established in nude mice to study the anticarcinogenic activities and apoptosis-regulatory mechanistic effect of aqueous extract of fermented barley with Lactobacillus plantarum dy-1 (LFBE).Methods HT-29 cells were transplanted via subcutaneous injection of 1×107 cells into the right flank of each nude mouse. Then, nude mice were treated for 30 days with LFBE (high-dose 2 g·kg-1·d-1; low-dose 1 g·kg-1·d-1) and for 7 days with 5-fluorouracil (5-FU, 25 g·kg-1·d-1) by gavage and intraperitoneal injection, respectively.Results Tumor volume and weight decreased significantly in both groups of nude mice treated with LFBE. In addition, the cell apoptosis rate of the LFBE group was significantly higher than that of the control group and 5-FU groups as measured by the TUNEL assay. Moreover, the real-time fluorescent quantitative PCR and Western blot methods further confirmed these apoptosis-enhancing and growth-inhibiting effects. The involvement of LFBE in inducing apoptosis was confirmed by the expression of Bax, Bcl-2, caspase-3, and cyclinD1.Conclusion The results showed that LFBE could induce subcutaneous transplantation tumor apoptosis in nude mice and could be used as a natural nutrient supplement or chemopreventive agent in the treatment of human colon cancer.
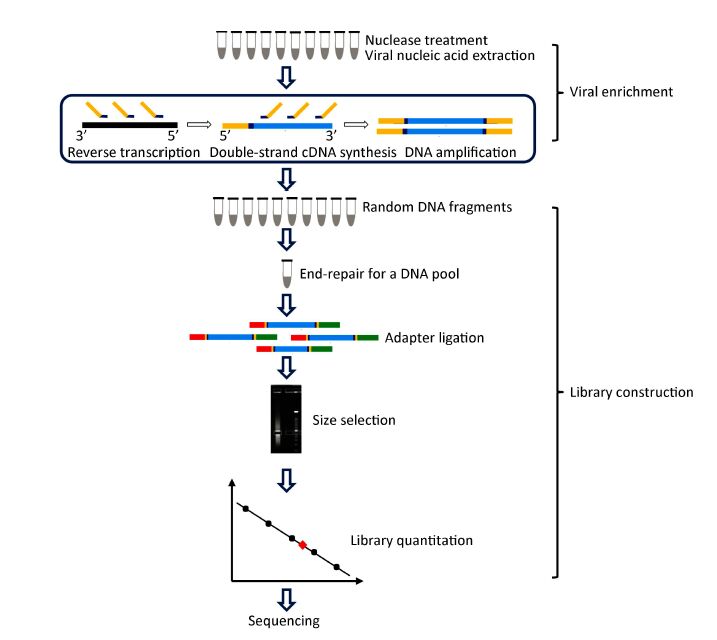
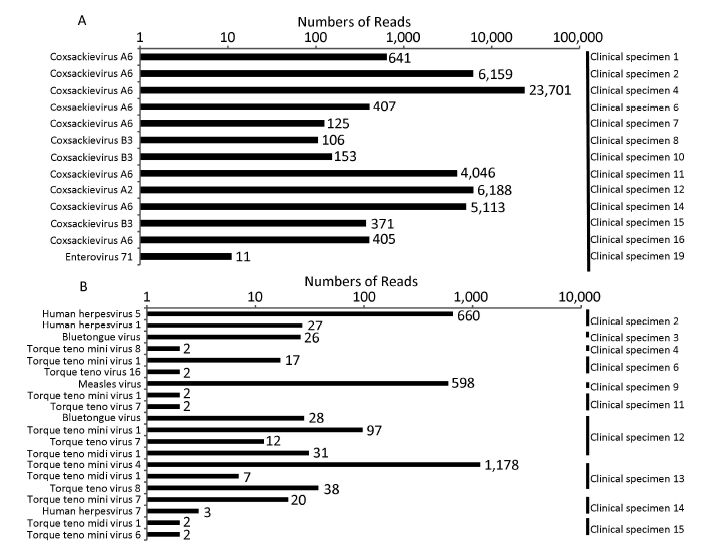
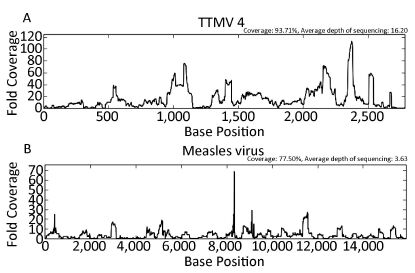
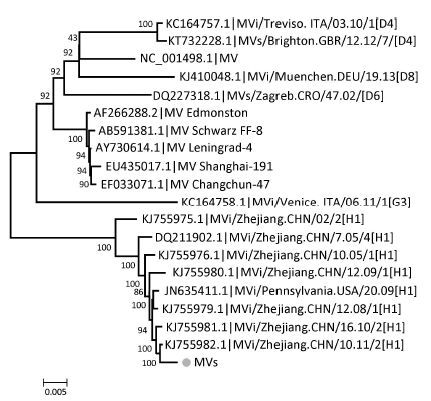
Objective To provide a feasible and cost-effective next-generation sequencing (NGS) method for accurate identification of viral pathogens in clinical specimens, because enormous limitations impede the clinical use of common NGS, such as high cost, complicated procedures, tremendous data analysis, and high background noise in clinical samples.Methods Viruses from cell culture materials or clinical specimens were identified following an improved NGS procedure:reduction of background noise by sample preprocessing, viral enrichment by barcoded oligonucleotide (random hexamer or non-ribosomal hexanucleotide) primer-based amplification, fragmentation-free library construction and sequencing of one-tube mixtures, as well as rapid data analysis using an in-house pipeline.Results NGS data demonstrated that both barcoded primer sets were useful to simultaneously capture multiple viral pathogens in cell culture materials or clinical specimens and verified that hexanucleotide primers captured as many viral sequences as hexamers did. Moreover, direct testing of clinical specimens using this improved hexanucleotide primer-based NGS approach provided further detailed genotypes of enteroviruses causing hand, foot, and mouth disease (HFMD) and identified other potential viruses or differentiated misdiagnosis events.Conclusion The improved barcoded oligonucleotide primer-based NGS approach is simplified, time saving, cost effective, and appropriate for direct identification of viral pathogens in clinical practice.
Objective To understand the relationships between CDH13 (T-cadherin) genetic polymorphisms, adiponectin levels and ischemic stroke, and possible interactions between CDH13 polymorphisms and other risk factors.Methods We recruited 342 Chinese ischemic stroke sib pairs. We genotyped rs4783244 and rs7193788 on CDH13 using time-of-flight mass spectrometry genotyping technology and measured total and high-molecular weight (HMW) adiponectin levels. We investigated associations between SNPs and ischemic stroke, and interactions between SNPs and other risk factors using multi-level mixed-effects regression model.Results In individuals without ischemic stroke, CDH13 rs4783244 was associated with total adiponectin levels (per T:Coef=-0.257, P=0.001). CDH13 rs7193788 was associated with total adiponectin levels (per A:Coef=-0.221, P=0.001) and HMW adiponectin levels (per A:Coef=-0.163, P=0.003). rs7193788 was significantly associated with ischemic stroke (GA/AA vs. GG:OR=1.55, 95% CI:1.07 to 2.24, P=0.020) after Bonferroni correction (α=0.025). There was an interaction between rs7193788 and diabetes (P=0.036). Compared to diabetes-free individuals with rs7193788 GG genotype, diabetes patients with rs7193788 GA/AA genotypes had higher risks for ischemic stroke (OR=2.64, 95% CI:1.58-4.40, P<0.001).ConclusionCDH13 genetic polymorphisms are associated with adiponectin levels and ischemic stroke. An interaction is found between CDH13 SNP and diabetes for ischemic stroke.
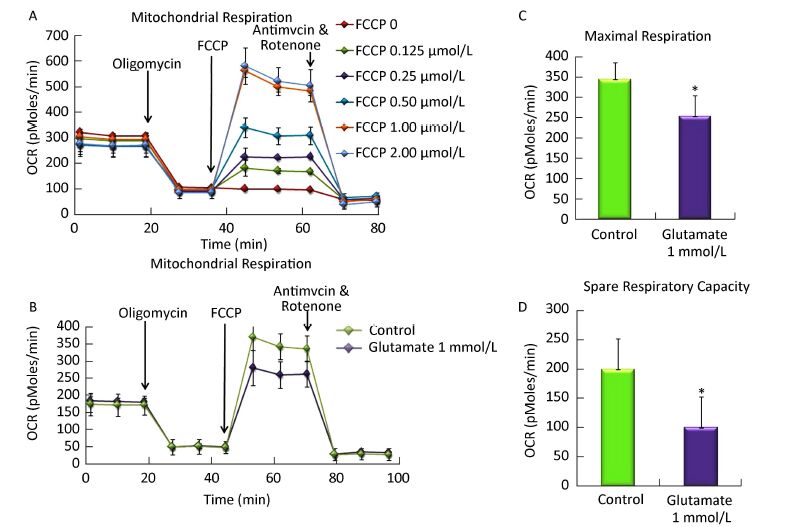
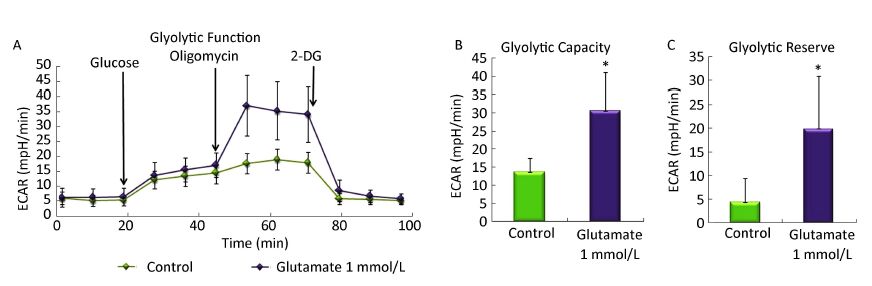
Objective To study the effect of glutamate on metabolism, shifts in glycolysis and lactate release in rat astrocytes.Methods After 10 days, secondary cultured astrocytes were treated with 1 mmol/L glutamate for 1 h, and the oxygen consumption rates (OCR) and extra cellular acidification rate (ECAR) was analyzed using a Seahorse XF 24 Extracellular Flux Analyzer. Cell viability was then evaluated by MTT assay. Moreover, changes in extracellular lactate concentration induced by glutamate were tested with a lactate detection kit.Results Compared with the control group, treatment with 1 mmol/L glutamate decreased the astrocytes' maximal respiration and spare respiratory capacity but increased their glycolytic capacity and glycolytic reserve. Further analysis found that 1-h treatment with different concentrations of glutamate (0.1-1 mmol/L) increased lactate release from astrocytes, however the cell viability was not affected by the glutamate treatment.Conclusion The current study provided direct evidence that exogenous glutamate treatment impaired the mitochondrial respiration capacity of astrocytes and enhanced aerobic glycolysis, which could be involved in glutamate injury or protection mechanisms in response to neurological disorders.


































 Quick Links
Quick Links