
2017 Vol. 30, No. 12





2017, 30(12): 863-874.
doi: 10.3967/bes2017.117
2017, 30(12): 875-886.
doi: 10.3967/bes2017.118
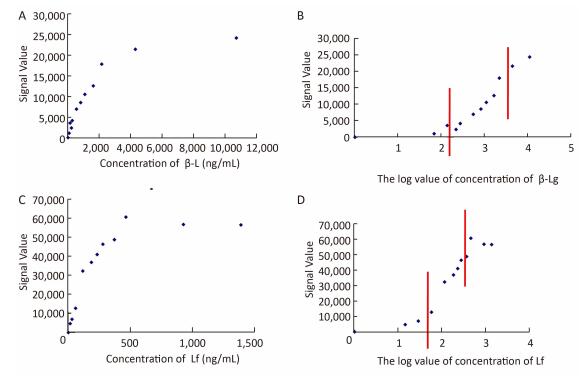



2017, 30(12): 898-906.
doi: 10.3967/bes2017.120

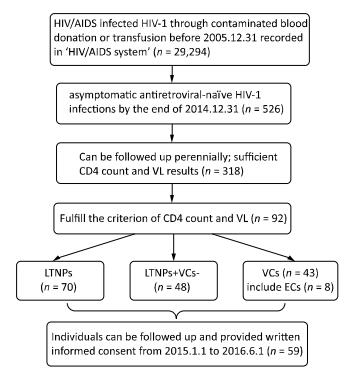
2017, 30(12): 907-912.
doi: 10.3967/bes2017.121

To characterize long-term nonprogressors (LTNPs) and viremia controllers (VCs), infected with HIV-1 through contaminated blood donation or transfusion between 1992 and 1996 in Henan, China. LTNPs and VCs were defined by CD4+T lymphocyte (CD4) count and viral load (VL). Of 29, 294 patients infected with HIV-1 via contaminated blood donation or transfusion that had conducted for more than 20 years, 92 were LTNPs/VCs. There were 70 LTNPs (0.24%), 43 VCs (0.15%), and 48 LTNPs+VCs-(0.16%). VCs had a significantly lower CD4 nadir, compared to LTNPs and LTNPs+VCs-, and no significant differences for the highest VL and HIV-1 DNA. Cases P4 and P5 were LTNPs, while their VL reached approximately 4.3 log copies/mL. P6 was a VC, but with CD4 < 500 cells/μL constantly. Data from the LTNPs/VCs cohort provided valuable information, future research is needed.
To evaluate the HIV pandemic in Chongqing, the pooled PCR, IgG-capture BED enzyme immunoassay (BED-CEIA), and cohort observations were used to estimate the HIV incidences among men who have sex with men (MSM). 617 MSM subjects completed the survey at a voluntary counseling and testing (VCT) site. The observed HIV incidence was 12.5 per 100 P-Ys (95% CI = 9.1-15.7). The annual acute HIV infection (AHI) incidence estimated by pooled PCR was 14.0% (95% CI = 10.9-17.1). The HIV-1 annual incidence estimated based on the BED-CEIA was 12.0% (95% CI = 7.5-16.5). The HIV incidences estimated by these three approaches were consistent and complementary. The HIV incidence rates were alarmingly high with an uptrend among the urban MSM of Chongqing.
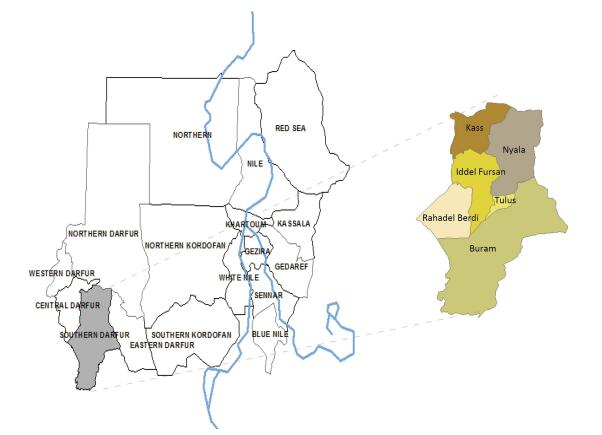
2017, 30(12): 917-921.
doi: 10.3967/bes2017.123
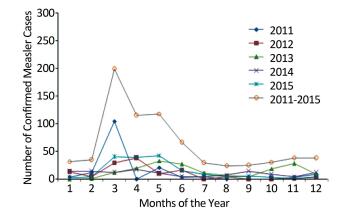
Case-based surveillance measles data was defined according to World Health Organization (WHO) guidelines. A total of 511 measles cases were studied from 2011 to 2015 in Southern Darfur State, Sudan and 58.1% of cases were confirmed from Nyala city. About 43.4% of cases were males, 56.6% of cases were female, and 47.7% were children under five years old. Similarity, within February to June, the cases increased by 8.0% in children vaccinated through measles campaign, and 5.3% in children that used child vaccination card and 78.7% in unvaccinated one. The epidemiologically linked (EPI-Linked) measles cases declined from 2011 to 2015; consequently, Measles still remain to be a significant challenge in south Darfur state, Sudan.
Kidney stones are a common urinary system condition that can progress to kidney disease. Previous studies on the association between tea consumption and kidney stones are inconsistent. A cross-sectional study to investigate the association between tea consumption and kidney stones was conducted from 2013 to 2014 and recruited 9, 078 northern Chinese adults. A total of 8, 807 participants were included in the final analysis. Participants' prevalence of kidney stones was 1.07%, 1.73%, and 2.25% based on their tea consumption frequency of never, occasionally, and often groups, respectively. Compared with the 'never' group, the odds ratios (95% confidence intervals) for the occurrence of kidney stones were 1.57 (1.00-2.46) and 1.65 (1.06-2.57) in the 'occasionally' and 'often' groups, respectively. After adjusting for sex, age, and other potential confounding factors, tea consumption still significantly increased the risk of kidney stones. Tea consumption is independently associated with an increased risk of kidney stones in the investigated population, suggesting that a decrease in the consumption of tea may be a preventive strategy for kidney stones.
2017, 30(12): 927-931.
doi: 10.3967/bes2017.125
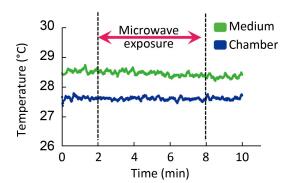
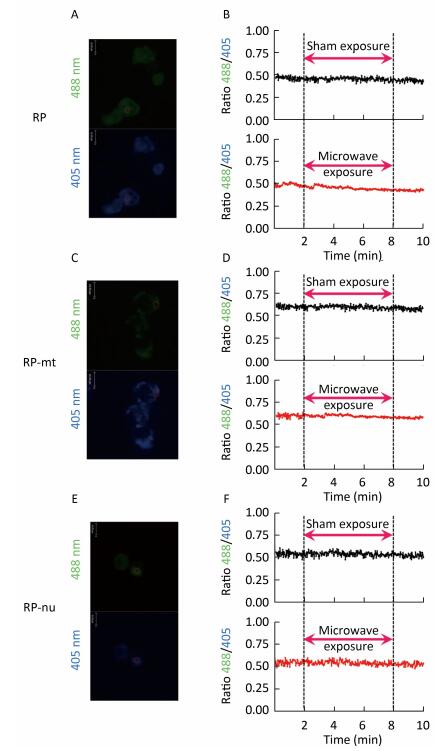
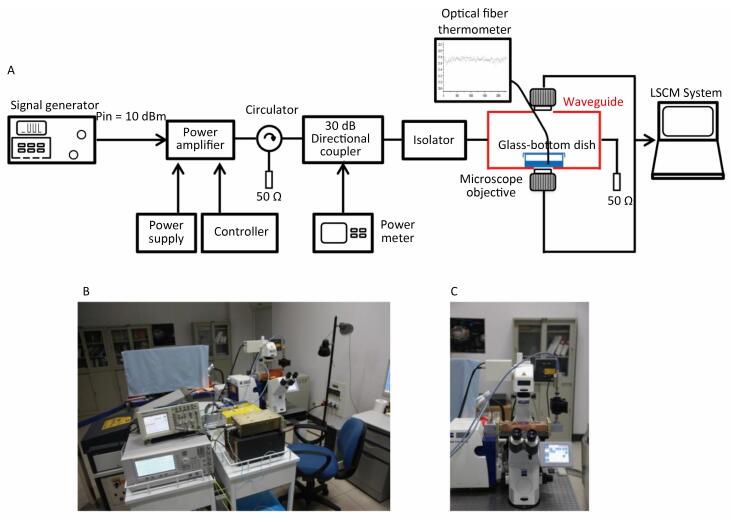
Little information is available about the effects of exposure to pulsed microwaves on neuronal Ca2+ signaling under non-thermal conditions. In this study, rat pheochromocytoma (PC12) cells were exposed to pulsed microwaves for 6 min at a specific absorption rate (SAR) of 4 W/kg to assess possible real-time effects. During microwave exposure, free calcium dynamics in the cytosol, mitochondria, and nucleus of cells were monitored by time-lapse microfluorimetry using a genetically encoded calcium indicator (ratiometric-pericam, ratiometric-pericam-mt, and ratiometric-pericam-nu). We established a waveguide-based real-time microwave exposure system under accurately controlled environmental and dosimetric conditions and found no significant changes in the cytosolic, mitochondrial, or nuclear calcium levels in PC12 cells. These findings suggest that no dynamic changes occurred in [Ca2+]c, [Ca2+]m, or [Ca2+]n of PC12 cells at the non-thermal level.

2017, 30(12): 932-937.
doi: 10.3967/bes2017.126
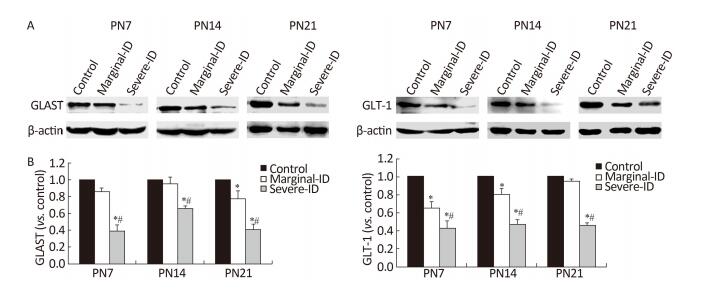
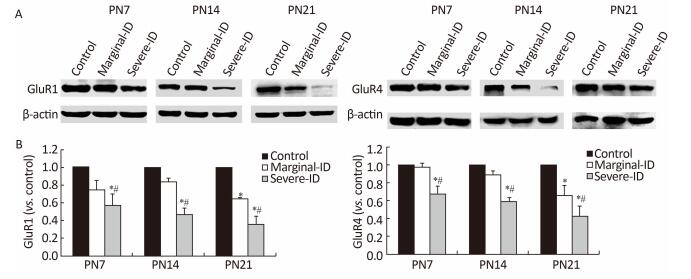
Iodine deficiency (ID) during early pregnancy has an adverse effect on children's psychomotor and motor function but the mechanism has not been clarified. Therefore, our aim was to study the effect of maternal marginal ID on cerebellar neurodevelopment and the underlying mechanism. After obtaining marginal ID rats, we examined interactions between Bergmann glia cells (BGs) and Purkinje cells (PCs) using immunofluorescence and expression of the glutamate transporter and receptor by western blot. Our results showed that marginal ID reduced the number of contacted points between BGs and PCs, and disturbed expression of the glutamate transporter and receptor. Our results support the hypothesis that marginal ID inhibits interactions of BGs-PCs, which may be involved in abnormal regulation of the glutamate transporter and receptor.
2017, 30(12): 938-942.
doi: 10.3967/bes2017.127


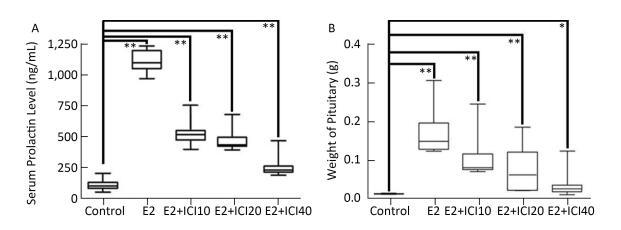
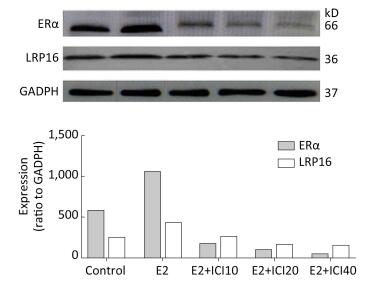
Prolactinoma is an estrogen-related tumor and leukemia-related protein 16 (LRP16) is correlated with the progression of estrogen-related tumors, but the regulatory mechanism between LRP16 and prolactinoma remain unclear. This study demonstrates a variation in LRP16 with estrogen receptor α (ERα) in prolactinoma models and the up and downregulation effects of LRP16 on prolactin secretion of pituitary adenomas cells (GH3 cells). In our study, 50 male SD rats (30-day-old) were randomly divided into five groups of 10 rats each. After 120 days of treatment, the rats were sacrificed, and the expression of LRP16 and ERα were examined by Western blot and immunohistochemistry to explore the changes in ERα, LRP16, and prolactin. After siRNA transfection of the respective genes, the GH3 cells were cultured, and their secretory function as well as the expression of ERα mRNA and prolactin were analyzed by enzyme-linked immunosorbent assay and real-time-polymerase chain reaction analysis. The results show that secretion of prolactin by GH3 cells can be affected by up and downregulating LRP16 expression, which may provide a novel medical therapy in clinical trials.













 Quick Links
Quick Links