-
Aeromonas, which belongs to Aeromonadaceae, is a Gram-negative facultative anaerobe positive for oxidase and catalase. Aeromonas spp. are widely distributed in aquatic environments[1], and more than 28 Aeromonas spp. have been identified[2]. At least 19 of these species are associated with human diseases, and they can cause widespread infection[3]. The common clinical manifestations of Aeromonas are diarrhea, localized soft tissue infection, and bacteremia[4]. Bacteremia primarily occurs in patients with underlying diseases, such as hepatobiliary diseases, cancer, and diabetes[5].
Phenotypically identifying Aeromonas species is considered reasonably difficult because of the limitations of commercially available identification systems for Aeromonas, e.g., API20E, API20NE, Vitek, and BioMerienx[6]. Therefore, molecular methods are widely used to identify Aeromonas spp. correctly. The nucleotide sequences of housekeeping genes, such as gyrB, cpn60, and rpoB, have been regarded as reliable markers that can be used to identify Aeromonas species[7-9]. Some reports have shown that strains initially identified as A. hydrophila via phenotypic methods can be recognized as A. dhakensis through cpn60 or gyrB sequencing[8,10]. A. dhakensis was initially described as an A. hydrophila subspecies in 2002 and as A. aquariorum; in 2013, it was described as a novel species, namely, Aeromonas dhakensis[11]. A. dhakensis has been shown to be a pathogenic bacterium linked to numerous human diseases[5]. Previous studies on A. aquariorum recognized the virulence of A. dhakensis, and subsequent evaluations have shown that this strain carries several important virulence genes[6]. This strain also has the most potent cytotoxic activities against human blood cell lines[10]. However, the prevalence of A. dhakensis infection is likely underestimated because phenotypic identification systems often misidentify A. dhakensis as A. hydrophila.
Aeromonas sp. pathogenesis involves a series of virulence factors[12], including hemolytic toxins, such as aerolysin-related cytotoxic enterotoxin (act)[13], heat-labile cytotoxic enterotoxin (alt), hemolysin (hlyA), heat-stable cytotonic toxins (ast)[14], and aerolysin (aerA)[15]. In addition, the type III secretion system (TTSS), lateral flagella (laf), polar flagellum (fla)[16], elastase (ela), and lipase (lip) contribute to Aeromonas pathogenicity[17]. With these virulence factors, bacteria can colonize, invade, and overcome their hosts’ immune response, leading to infection and disease development.
Diarrhea caused by Aeromonas is self-limiting, and it is effectively treated with the oral or intravenous antibiotic infusion. However, patients with severe diarrhea or extraintestinal infection should receive antibacterial treatments[18]. Aeromonas is resistant to ampicillin, but it is susceptible to third-generation cephalosporins and aminoglycosides that exhibit potent antibacterial effects against clinical Aeromonas isolates[19]. However, the extensive use of antibiotics in aquaculture and clinical treatment has caused an increase in antimicrobial resistance[20]. The development of resistance in Aeromonas spp. against several different classes of antibiotics threatens human health because resistant isolates can be transmitted from the aquatic environment to humans via the food chain or direct contact[21]. Therefore, the antimicrobial resistance of Aeromonas should be monitored to guide treatments.
In this study, the diversity and distribution of Aeromonas species isolated from healthy individuals and clinical patients in Ma’anshan, Anhui Province, China, were evaluated. Their genetic diversity, pathogenicity, and antimicrobial resistance profiles were compared.
-
A total of 57 Aeromonas isolates (38 from clinical patients and 19 from healthy individuals) were obtained from Ma’anshan, Anhui Province, China, between 2018 and 2019 (Table 1). They were identified using an automatic bacteriological analyzer (Vitek 2 Compact, BioMèrieux) and cultured on Luria–Bertani (LB) broth or brain heart infusion agar plates overnight at 37 °C.
Species Total no. strains (%) No. clinical patients (%) No. healthy people (%) A. caviae 21 (36.8) 18 (47.3) 3 (15.8) A. veronii 19 (33.3) 10 (26.3) 9 (47.4) A. dhakensis 6 (10.5) 5 (13.2) 1 (5.3) A. jandaei 6 (10.5) 1 (2.6) 5 (26.3) A. hydrophila 2 (3.5) 1 (2.6) 1 (5.3) A. media 2 (3.5) 2 (5.3) 0 A. enteropelogenes 1 (1.8) 1 (2.6) 0 Total 57 38 19 Table 1. Distribution of Aeromonas spp. in isolates collected from clinical patients and healthy individuals
-
The Aeromonas MLST scheme (
http://pubmlst.org/Aeromonas/ ), which relies on the amplification and sequencing of six housekeeping genes, namely, gyrB, groL, gltA, metG, ppsA, and recA, was applied to identify the subtype of each Aeromonas isolate[22]. The total chromosomal DNA from the Aeromonas samples was prepared using a DNA purification kit (Promega, Madison, USA) in accordance with the manufacturer’s instructions. PCR was performed using previously described primers and protocols[23], and the sequences of the six loci were compared with those hosted on the Aeromonas MLST database and STs. New alleles and STs were submitted to the Aeromonas MLST database for naming.With the limitations related to molecular identification involving 16S rRNA sequencing[24], the sequences of the housekeeping genes gyrB and cpn60 were included in our species identification assays. Concatenated two-gene phylogenetic trees were constructed and compared with representative species as previously described[25]. A phylogenetic tree was also constructed using the neighbor-joining method in Clustal-W[24] with Bootstrap values calculated using 1,000 replicates.
-
The degree of beta-hemolysis was assessed in isolates cultured in LB agar containing 5% (vol/vol) sheep blood agar (KeMaJia, Shanghai, China). Qualitative assays were performed with LB agar containing 2% (wt/vol) skim milk (KeMaJia, Shanghai, China) to investigate the exoprotease activity. The presence of clear zones surrounding the streaks indicated positive reactions in both tests[26].
-
An assay was designed to evaluate the presence of the genes encoding exotoxins or secretion system components, structural components, and extracellular enzymes associated with Aeromonas virulence. In this assay, aerolysin (aerA), hemolysin (hlyA), heat-labile enterotoxin (act), heat-stable cytotoxin (ast), heat-labile cytotoxin (alt), type III secretion system components (ascV), ADP-ribosyl transferase toxin (aexT), lipase (lip), elastase (ela), and flagellin (fla) were detected using previously described primers and protocols[25]. PCR was performed in a 50 μL reaction mixture containing 25 μL of Taq PCR MasterMix (Takara Bio, Inc., Japan), 1 μL of the relevant primers (10 µmol/L), 21 μL of ddH2O, and 2 μL of DNA template under the following cycling conditions: pre-denaturation at 95 °C for 5 min; 30 cycles of denaturation at 95 °C for 30 s, annealing at 55–60 °C for 30 s, and extension at 72 °C for 1 min; and a final cycle at 72 °C for 5 min. Sequencing was conducted to confirm the positive PCR products.
-
The antimicrobial susceptibility of each Aeromonas isolate was evaluated using the microbroth dilution method in accordance with the current guidelines of the Clinical and Laboratory Standards Institute. The minimum inhibitory concentrations of 12 antibiotics were measured: amoxicillin/clavulanate, cefepime, ceftriaxone, ceftazidime, imipenem, aztreonam, gentamycin, tetracycline, ciprofloxacin, trimethoprim/sulfamethoxazole (SXT), chloramphenicol, and colistin. Escherichia coli ATCC 25922 was the strain used for the quality control strain of all the susceptibility tests.
-
The strains were screened for the presence of various antimicrobial resistance genes, including those for tetracycline resistance (tetA, tetB, and tetE), extended-spectrum b-lactamase (ESBL) resistance (blaTEM, blaSHV, and blaCTX)[27], aminoglycoside resistance (armA, aphAI-IAB, aac(6ʹ)-Ib, and aac(3)-IIa)[28], sulfonamide resistance (sul1 and sul2;)[29], mobile colistin resistance (mcr-1, mcr-2, mcr-3, and mcr-4), and PMQR (qnrA, qnrB, and qnrS)[27], by using previously described primers and protocols[25]. Sequencing was conducted to confirm the positive PCR products.
-
The 57 Aeromonas isolates were divided into 55 STs. Of these isolates, 21 were novel (ST722-ST742), indicating that the degree of genetic diversity was high (Figure 1). No dominant STs were found in either the patient or healthy groups.

Figure 1. Phylogenetic relationships were established using the concatenated sequences of the six genes included in this study. The source, species, virulence genes, and antibiotic resistance are shown on the right. A phylogenetic tree was determined using a neighbor-joining algorithm. ST: sequence type.
-
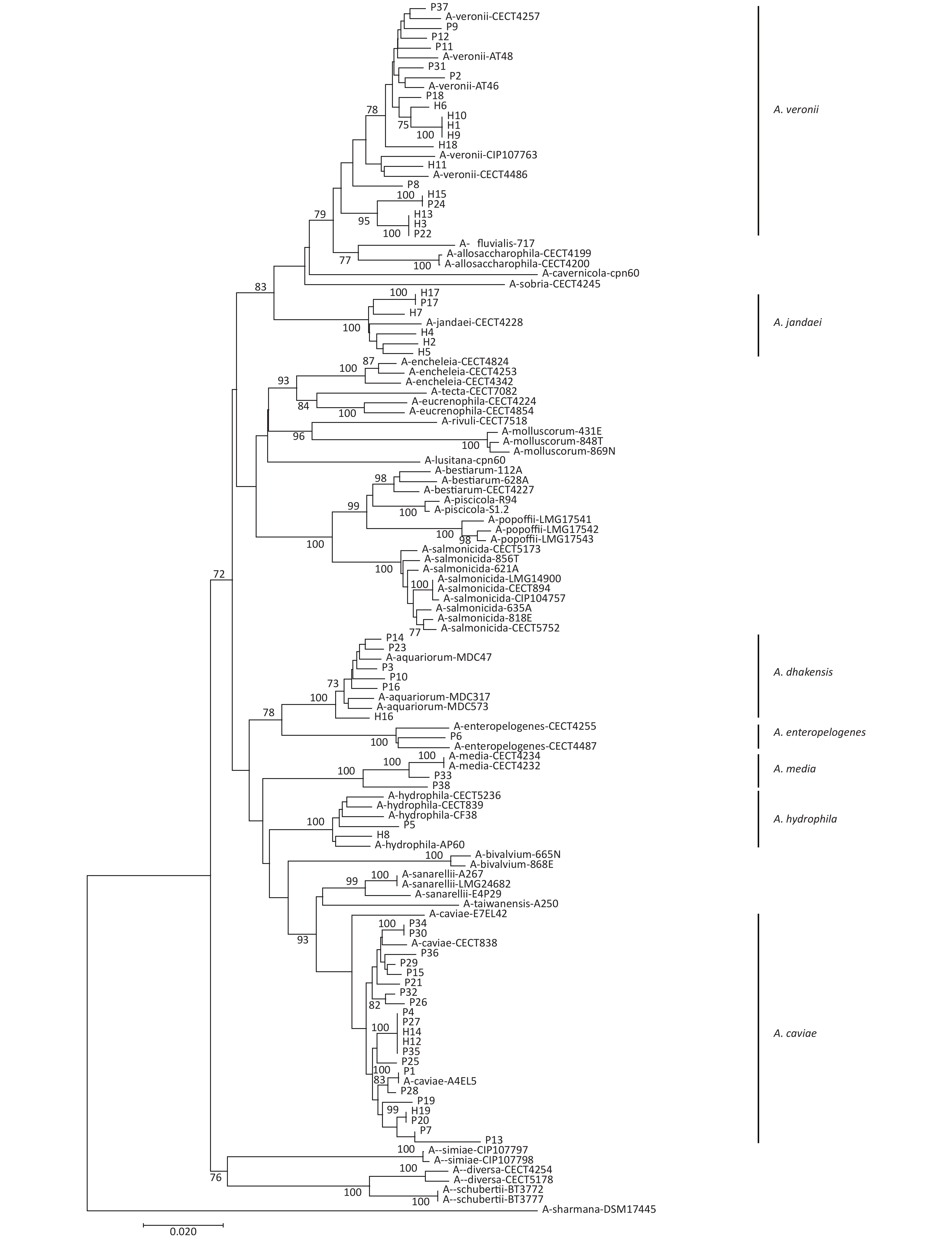
gyrB and cpn60 were sequenced to evaluate the phylogeny of the 57 Aeromonas isolates identified in this study (Figure 2). This analysis revealed that these isolates could be divided into seven different species. The most common species were A. caviae (36.8%), A. veronii (33.3%), A. dhakensis (10.5%), and A. jandaei (10.5%). Notably, the distribution of the Aeromonas isolates from the patients and the healthy individuals varied (Table 1). A. caviae (47.3%) and A. veronii (26.3%) were the most prevalent in the patient samples, whereas A. veronii (47.4%) and A. jandaei (26.3%) were the most common species in the healthy samples. In addition, the abundance of A. dhakensis was higher in the patients than in the healthy individuals.
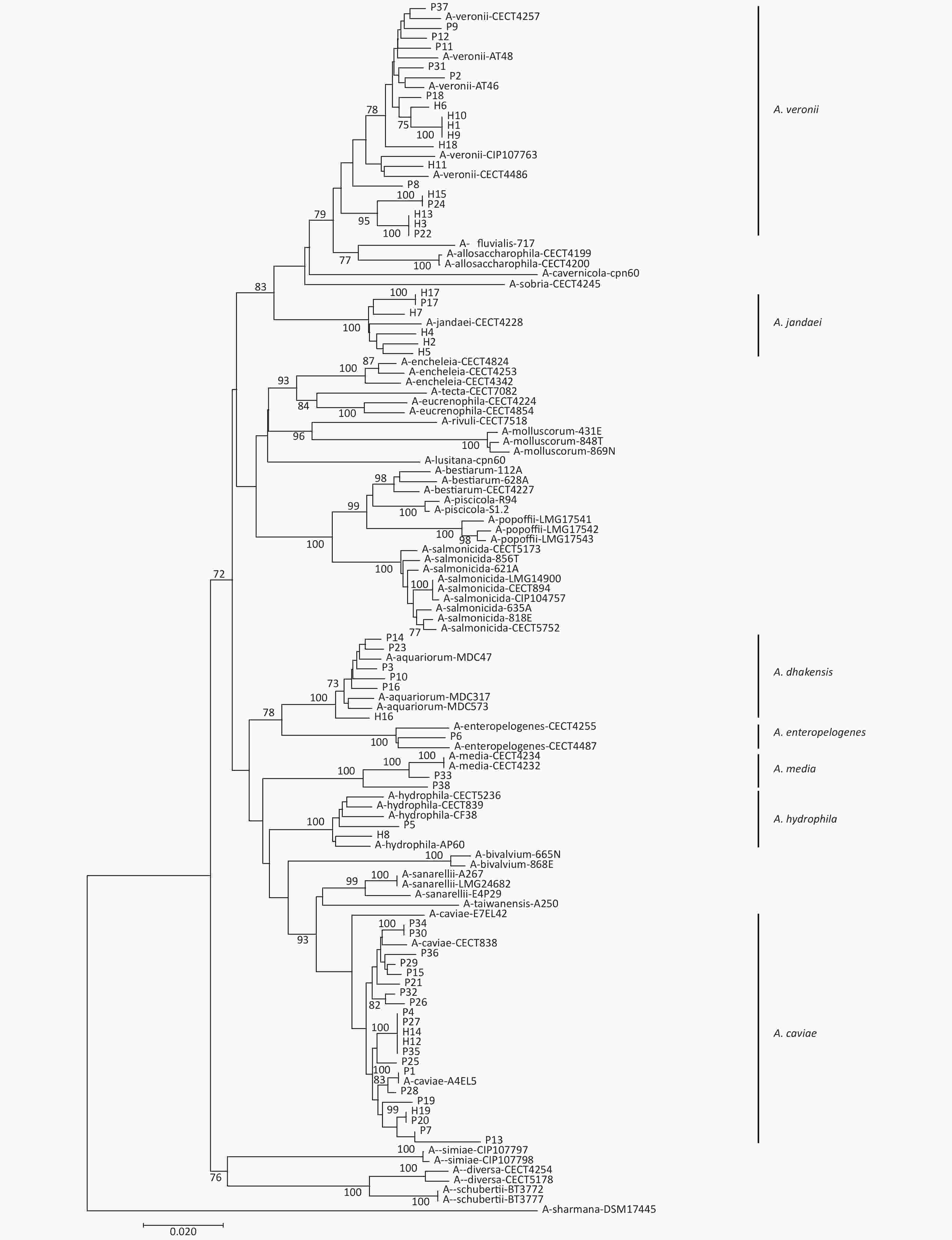
Figure 2. Neighbor-joining phylogenetic tree was constructed using the concatenated sequences of gyrB and cpn60 to reveal the relationships between 57 Aeromonas isolates from clinical patients and healthy individuals from Ma’anshan, Anhui Province, China. Isolates were designated as either P or H to indicate the strains isolated from clinical patients or healthy individuals, respectively.
-
A total of 28 (49.1%) isolates demonstrated beta-hemolysis, and 42 (73.7%) isolates possessed a proteolytic activity (Table 2). A. media isolates lacked hemolytic and proteolytic activities. A. caviae isolates (19.0% and 66.7%, respectively) had less hemolytic and exoprotease activities than those of A. dhakensis isolates (83.3% and 100%, respectively) or A. hydrophila isolates (50% and 100%, respectively). Of the strains isolated from healthy individuals, only 36.8% processed any beta-hemolytic activity, whereas 63.2% exhibited some proteolytic activities. However, 55.2% of the clinical isolates exhibited a hemolytic activity, and 84.2% had a proteolytic activity (Table 2).
Species and source No. (%) of isolates Total Healthy individual Clinical patient A. veronii 19 (100) 9 (47.5) 10 (52.6) Hemolysis 13 (68.4) 4 (44.4) 9 (90.0) Proteolysis 15 (78.9) 6 (66.7) 9 (90.0) A. jandaei 6 (100) 5 (83.3) 1 (16.7) Hemolysis 4 (66.7) 3 (60.0) 1 (100) Proteolysis 4 (66.7) 3 (60.0) 1 (100) A. dhakensis 6 (100) 1 (16.7) 5 (83.3) Hemolysis 5 (83.3) 0 (0) 5 (100) Proteolysis 6 (100) 1 (100) 5 (100) A. caviae 21(100) 3 (14.4) 18 (85.7) Hemolysis 4 (19.0) 0 (0) 4 (22.2) Proteolysis 14 (66.7) 1 (33.3) 13 (72.2) A. enteropelogenes 1 (100) 0 (0) 1 (100) Hemolysis 1 (100) 0 (0) 1 (100) Proteolysis 1 (100) 0 (0) 1 (100) A. media 2 (100) 0 (0) 2 (100) Hemolysis 0 (0) 0 (0) 0 (0) Proteolysis 0 (0) 0 (0) 0 (0) A. hydrophila 2 (100) 1 (50) 1 (50.0) Hemolysis 1 (50) 0 (0) 1 (100) Proteolysis 2 (100) 1 (100) 1 (100) Total 57 (100) 19 (33.3) 38 (66.7) Hemolysis 28 (49.1) 7 (36.8) 21 (55.2) Proteolysis 42 (73.7) 12 (63.2) 32 (84.2) Table 2. Prevalence of hemolytic and proteolytic activity in Aeromonas isolates
-
The distribution of the virulence genes in these Aeromonas isolates is summarized in Table 3. In particular, fla was the most common virulence gene identified in 87.7% of the 57 isolates, and ela was the second-most prevalent virulence gene in these isolates (n = 41, 71.9%). Laf, ast, and aerA were identified in less than 10% of the isolates. The prevalence of lip differed significantly between patient- and healthy-derived strains (P < 0.05, Fisher's exact test). Only lip, alt, and aerA were more prevalent in patient-derived isolates than in healthy-derived isolates.
Gene Total no. strains (%) No. clinical patients (%) No. healthy individuals (%) fla 50 (87.7) 33 (86.8) 17 (89.5) ela 41 (71.9) 27 (71.1) 14 (73.7) lip 34 (59.6) 28 (73.6) 6 (31.6) act 25 (43.9) 16 (42.1) 9 (50.0) ascV 18 (31.6) 11 (28.9) 7 (36.8) alt 19 (33.3) 16 (42.1) 3 (15.8) aexT 16 (28.1) 10 (26.3) 6 (31.6) hlyA 9 (15.8) 6 (15.8) 3 (15.8) laf 5 (8.8) 2 (5.3) 3 (15.8) ast 2 (3.5) 1 (2.6) 1 (5.3) aerA 2 (3.5) 2 (5.3) 0 (0.0) Total 57 (100) 38 (100) 19 (100) Table 3. Distribution of virulence-associated genes in Aeromonas strains isolated from clinical patients and healthy individuals
In the four most common Aeromonas species, the detection rates of ela, lip, alt, hlyA, and aerA in A. dhakensis isolates were significantly higher than those in A. caviae, A. veronii, and A. jandaei (P < 0.05, Fisher’s exact test; Table 4). Although the total number of A. jandaei isolates was low, few of them were positive for any of the evaluated virulence genes. The extracellular protease genes ela and lip were abundant in A. caviae and A. dhakensis, and the hemolytic gene act was abundant in A. veronii. The enterotoxin gene alt and hemolytic genes hlyA and aerA were most commonly identified in A. dhakensis. Their rates of occurrence in other species identified in this study were relatively low.
Gene No. A. caviae strains (%) No. A. veronii strains (%) No. A. dhakensis strains (%) No. A. jandaei strains (%) fla 17 (80.9) 18 (94.7) 6 (100) 4 (66.7) ela 21 (100) 5 (26.3) 6 (100) 5 (83.3) lip 21 (100) 2 (10.5) 6 (100) 0 (0) act 5 (23.8) 17 (89.4) 2 (33.3) 0 (0) ascV 3 (14.3) 12 (63.2) 1 (16.7) 1 (16.7) alt 8 (38.1) 0 (0) 6 (100) 0 (0) aexT 3 (14.3) 12 (63.2) 1 (16.7) 0 (0) hlyA 0 (0) 0 (0) 6 (100) 0 (0) laf 1 (4.8) 2 (10.5) 1 (16.7) 0 (0) ast 0 (0) 0 (0) 0 (0) 0 (0) aerA 0 (0) 0 (0) 1 (16.7) 0 (0) Total 21 (100) 19 (100) 6 (100) 6 (100) Table 4. Distribution of virulence genes in the four most common Aeromonas spp.
-
The antimicrobial susceptibility rates of 12 common antimicrobials in the Aeromonas isolates identified in this study are presented in Table 5. Almost all the Aeromonas strains had high amoxicillin/clavulanic acid (96.4%) resistance; however, most of the isolates (≥ 90%) were susceptible to cefepime, imipenem, chloramphenicol, gentamicin, and tetracycline. In general, clinical A. dhakensis, A. hydrophila, A. jandaei, and A. caviae isolates were more resistant than their healthy individual counterparts to the commonly prescribed antibiotics, including ceftriaxone, ceftazidime, ciprofloxacin, and amoxicillin/clavulanic acid. The healthy sample isolates had an increased rate of resistance to colistin and imipenem only (Supplementary Table S1 available in www.besjournal.com). A. enteropelogenes and A. media, identified only in the patient samples, were susceptible to most of the tested antibiotics except amoxicillin/clavulanic acid, aztreonam, ceftriaxone, and ceftazidime (Supplementary Table S1).
Antibiotics No and % of resistant isolates Total no. strains (%) No. clinical patients (%) No. healthy individuals (%) Penicillins Amoxicillin/clavulanic acid 55 (96.4) 36 (94.7) 19 (100) Caphems Cefepime 4 (7.0) 4 (10.5) 0 (0) Ceftazidime 12 (21.1) 11 (28.9) 1 (5.3) Ceftriaxone 13 (22.8) 12 (31.6) 1 (5.3) Carbapenems Imipenem 2 (3.5) 0 (0.0) 2 (10.5) Monobactams Aztreonam 7 (12.3) 6 (15.8) 1 (5.3) Aminoglycosides Gentamicin 5 (8.8) 4 (10.5) 1 (5.3) Tetracyclines Tetracycline 5 (8.8) 5 (13.2) 0 (0.0) Quinolones Ciprofloxacin 8 (14.0) 7 (18.4) 1 (5.3) Folate pathway inhibitors Trimethoprim-sulfamethoxazole 13 (22.8) 11 (28.9) 2 (10.5) Phenicols Chloramphenicol 5 (8.8) 5 (13.2) 0 (0.0) Polymyxins Colistin 15 (26.3) 9 (23.7) 6 (31.6) Table 5. Prevalence of resistance to different antibiotics
Drug No and % of resistant isolates A. veronii A. jandaei A. dhakensis A. enteropelogenes A. media A. hydrophila A. caviae Clinical
(n = 10)Healthy
(n = 9)Clinical
(n = 1)Healthy
(n = 5)Clinical
(n = 5)Healthy
(n = 1)Clinical
(n = 1)Healthy
(n = 0)Clinical
(n = 2)Healthy
(n = 0)Clinical
(n = 1)Healthy
(n = 1)Clinical
(n = 17)Healthy
(n = 3)IPM 0 (0.0) 1 (11.1) 0 (0.0) 1 (20) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) CHL 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 1 (100) 0 (0.0) 4 (23.5) 0 (0.0) FEP 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 1 (100) 0 (0.0) 3 (17.6) 0 (0.0) CRO 1 (10) 1 (11.1) 0 (0.0) 0 (0.0) 1 (20) 0 (0.0) 0 (0.0) 1 (50) 1 (100) 0 (0.0) 8 (47.1) 0 (0.0) CAZ 0 (0.0) 0 (0.0) 1 (100) 0 (0.0) 2 (40) 0 (0.0) 0 (0.0) 1 (50) 1 (100) 0 (0.0) 6 (35.3) 0 (0.0) CT 3 (30) 1 (11.1) 1 (100) 4 (80) 5 (100) 1 (100) 0 (0.0) 0 (0.0) 1 (100) 1 (100) 0 (0.0) 0 (0.0) GEN 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 1 (20) 0 (0.0) 0 (0.0) 0 (0.0) 1 (100) 0 (0.0) 2 (11.8) 1 (33.3) TET 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 5 (29.4) 0 (0.0) CIP 1 (10) 1 (11.1) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 1 (100) 0 (0.0) 5 (29.4) 0 SXT 0 (0.0) 1 (11.1) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 1 (100) 0 (0.0) 9 (52.9) 1 (33.3) AMC 10 (100) 9 (100) 1 (100) 0 (0.0) 5 (100) 0 (0.0) 1 (100) 2 (100) 1 (100) 0 (0.0) 15 (88.2) 0 (0.0) ATM 1 (10) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 0 (0.0) 2 (100) 0 (0.0) 0 (0.0) 3 (17.6) 1 (33.3) Note. *IPM, imipenem; CHL, chloramphenicol; FEP, cefepime; CRO, ceftriaxone; CAZ, ceftazidime; CT, colistin; GEN, gentamycin; TET, tetracycline; CIP, ciprofloxacin; SXT, trimethoprim/sulfamethoxazole; AMC, amoxicillin/clavulanate; ATM, aztreonam. Table S1. Antimicrobial resistance of Aeromonas isolates from clinical patient and healthy people*
Of the 57 strains identified in this study, 15 (26.3%) were found to have multidrug resistance (MDR), displaying resistance to at least 3 of the antibiotics tested in this assay. The MDR rates of the healthy-individual-derived strains (5.3%) were significantly lower than those of the patient isolates (36.8%). Therefore, these MDR phenotypes were likely acquired after treatment and hospitalization.
-
The tetracycline resistance genes tetA and tetE were found in 4 (6.9%) isolates and 1 (1.7%) isolate, respectively. The PMQR gene qnrS was present in 7 (12.1%) isolates, and the aminoglycoside resistance genes aac (6ʹ)-Ib and armA were detected in 3 (5.2%) isolates and 1 (1.7%) isolate, respectively. The ESBL genes blaCTX and bla-TEM were observed in 2 (3.4%) isolates and 1 (1.7%) isolate, and the sulfonamide genes sul1 and sul2 were found in 7 (12.1%) isolates and 1 (1.7%) isolate, respectively (Supplementary Table S2 available in www.besjournal.com). The mobile colistin resistance gene mcr-3 was detected in 1 (1.7%) isolate. The following genes were not detected in any isolate: the ESBL gene blaSHV; the aminoglycoside resistance genes aphAI-IAB and aac(3)-IIa; the tetracycline resistance gene tetB; the colistin resistance genes mcr-1, mcr-2, and mcr-4; and the PMQR genes qnrA and qnrB.
Gene No and % of resistant genes A. veronii A. jandaei A. dhakensis A. enteropelogenes A. media A. hydrophila A. caviae Clinical
(n = 10)Healthy
(n = 9)Clinical
(n = 1)Healthy
(n = 5)Clinical
(n = 5)Healthy
(n = 1)Clinical
(n = 1)Healthy
(n = 0)Clinical
(n = 2)Healthy
(n = 0)Clinical
(n = 1)Healthy
(n = 1)Clinical
(n = 17)Healthy
(n = 3)tetA 0 (0) 1 (11.1) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 3 (17.6) 0 (0) tetE 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (50) 0 (0) 0 (0) 0 (0) 0 (0) qnrS 1 1 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 5 (29.4) 0 (0) aac-6 0 (0) 1 (11.1) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (100) 0 (0) 1 (5.9) 0 (0) bla-TEM 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (5.9) 0 (0) bla-CTX 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 2 (11.8) 0 (0) armA 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (100) 0 (0) 0 (0) 0 (0) sul1 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (100) 0 (0) 6 (35.3) 0 (0) sul2 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 1 (5.9) 0 (0) mcr-3 0 (0) 0 (0) 1 (100) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) Table S2. Prevalence of antimicrobial resistance genes of Aeromonas isolates from clinical patient and healthy people
The detection rate of resistance genes in the clinical strains was higher than that in their healthy individual counterparts. The resistance genes of A. caviae were higher than that of any of the other species identified in this study (Supplementary Table S2).
-
Aeromonas spp. are ubiquitous in most aquatic environments, and certain species have been reported to be an important cause of acute diarrhea in children and adults[2]. Aeromonas strains were found in 7.6% of patients with diarrhea and 3.8% of a healthy population[30,31]. These rates may reflect actual differences or may be influenced by seasonal or environmental factors.
In this study, 57 Aeromonas isolates were described in detail in terms of species distribution, ST type, pathogenicity, and antimicrobial susceptibility to evaluate the differences in the epidemiology and etiology of Aeromonas strains isolated from clinical patients and healthy individual samples. The results of these analyses were then used to evaluate the significance of Aeromonas infections in clinical settings in China. They could be considered as a basis for developing the clinical protocols of treatment and surveillance.
These 57 isolates were separated into 55 STs. Of these isolates, 21 were novel, indicating that the degree of genetic diversity within these sample populations was high. Aeromonas is often isolated from patients with diarrhea. However, the role of Aeromonas spp. as potential human enteric pathogens remains unclear, possibly because of the lack of systematic analysis of these strains and the inherent difficulty in classifying them.
In this study, Aeromonas strains were identified to the species level via gyrB–cpn60 concatemer sequencing, and their species distribution was compared between the clinical patients and healthy individuals. The distribution of Aeromonas species isolated from the clinical patients and healthy individuals varied. Among them, A. caviae (47.3%) and A. veronii (26.3%) were the most prevalent clinical isolates, and A. veronii (47.4%) and A. jandaei (26.3%) were the most common isolates from the healthy individual samples. Wu et al.[32] reported that the most prevalent species in clinical patients are A. caviae and A. dhakensis, which account for 29.8%; they also found that A. veronii is the second-most prevalent species, which account for 23.4%. In the present study, the isolates were identified as follows: 21 (36.8%) as A. caviae, 19 (33.3%) as A. veronii, and 6 (10.5%) as A. dhakensis. Studies in other countries have shown a slightly different distribution; in particular, the most prevalent clinical strain in India is A. hydrophila, while the most prevalent one in France, Italy, and the Netherlands is A. caviae[30]. In addition, most A. veronii strains are isolated from human samples, whereas most A. hydrophila strains are obtained from water samples. These differences may also depend on sampling season and geographical location. A. dhakensis, the third-most prevalent species in our study, is a novel Aeromonas species[33] that has been identified as a potential pathogen in humans[5]. A. dhakensis isolates have been collected from stool, blood, wound, and other extraintestinal samples from humans globally; skin and soft tissue infections (SSTIs) caused by A. dhakensis have been reported in Taiwan, China[6]. In a set of 80 Aeromonas wound isolates, 37 (46.3%) samples contain A. dhakensis, which can cause severe SSTIs[34]. Chang et al.[8] demonstrated that A. dhakensis is the most frequently isolated aeromonad in Australia, accounting for 30.7%. In clinical specimens, A. dhakensis is the most prevalent in wounds but is less frequently isolated from fecal strains and blood samples. Salas et al.[35] believed that A. dhakensis is the second-most prevalent species in clinical isolates, accounting for 25.5%. Zhou et al.[31] revealed that the four most prevalent species of Aeromonas in clinical isolates are A. caviae (41.7%), A. veronii (31.3%), A. dhakensis (13.9%), and A. hydrophila (5.2%). Senderovich et al.[36] showed that several Aeromonas species are recovered from Israeli patients suffering from diarrhea; among the isolates, A. caviae (65.0%) and A. veronii (29.0%) are the most prevalent species. Therefore, previous studies supported our findings on the most prevalent strains in our samples.
The pathogenic mechanism of Aeromonas spp. is complex and multifactorial. In this study, the pathogenic potential was evaluated using beta-hemolytic and proteolytic activity assays, and several virulence genes were detected. A total of 28 (49.1%) isolates showed beta-hemolysis activities, and 42 (73.7%) had some proteolytic activities. Consistent with previous findings[32], our results indicated that A. dhakensis and A. hydrophila isolates were more likely to exhibit hemolytic and proteolytic activities than A. caviae isolates. The beta-hemolytic and proteolytic activities of the clinical isolates differed from those of the healthy individual isolates. In particular, 36.8% of the healthy sample strains demonstrated any beta-hemolytic activity, and 63.2% of these strains exhibited a proteolytic activity. However, 55.2% and 84.2% of the clinical isolates had hemolytic and proteolytic activities, respectively. Therefore, the strains from clinical patients were more likely to exhibit beta-hemolytic and proteolytic activities than their healthy sample counterparts (Table 2).
The distribution of the virulence-associated genes in these isolates was determined. The results revealed that fla, ela, and lip were identified in most of the strains, i.e., 50 (87.7%), 41 (71.9%), and 34 (59.6%) of the 57 isolates, respectively. In addition, fla was present in most species, while ela and lip were more prevalent in A. dhakensis and A. caviae isolates than in other isolates (Table 4). The enterotoxin and hemolysin genes act, aerA, alt, and ast were detected in 25 (43.9%), 2 (3.5%), 19 (33.3%), and 2 (3.5%) of the 57 isolates, respectively. act was observed in 89.4% of A. veronii isolates, and aerA was found in 16.7% of A. dhakensis isolates. alt was detected in 100% of A. dhakensis and 38.1% of A. caviae isolates. Conversely, ast was not present in any of the A. dhakensis, A. caviae, and A. veronii strains (Table 4). The TTSS genes ascV and aexT were present in 18 (31.6%) and 16 (28.1%) of the 57 isolates, respectively, and ascV and aexT were both present in 63.2% of A. veronii strains and 16.7% of A. dhakensis strains. The detection rate of lip was significantly higher in the clinical isolates than in the healthy individual isolates. By comparison, ascV and laf were detected more frequently in the strains from the healthy samples than from the clinical samples. Enterotoxins and hemolysins are important virulence factors in Aeromonas spp., and many studies have shown that the number of toxin genes harbored by any isolate is positively correlated with its potential virulence[37,38]. Among the four most common Aeromonas strains, A. dhakensis had the highest prevalence of virulence genes. This finding was also consistent with previous reports, which described that A. dhakensis strains harbor high cytotoxicity and liquid toxicity[34]. In our study, fla, ela, lip, alt, and hlyA were detected in all the A. dhakensis isolates, but aerA and hlyA were detected in A. dhakensis isolates only. The abundance of these virulence genes also varies among studies. For example, ast was not detected in A. dhakensis isolates in our study; however, some researchers have reported that 50.0%–83.3% ast is found in A. dhakensis isolates from wound and blood infections[6,10].
Consistent with previous findings[39], our results suggested that 96.4% of the Aeromonas strains exhibited resistance against amoxicillin/clavulanic acid. The resistance rates to most antibiotics were higher in the clinical isolates than in the healthy individual isolates, but the resistance to colistin and imipenem was higher in the strains identified in the healthy individual samples than in the clinical samples. All the A. dhakensis isolates exhibited resistance to colistin. This result suggested that A. dhakensis should be the focus of future research because it harbors a large number of virulence genes, high rates of drug resistance, and a high degree of beta-hemolytic and proteolytic activities. Some scholars indicated that Aeromonas has a high MDR, which is 38.7%, because of the different sources of strains[40]. In our study, 15 strains (26.3%) exhibited MDR to 12 antimicrobial agents, and this value was consistent with 28.7% as previously reported[31]. The MDR rates of the healthy individual-derived strains were significantly lower than those of the clinical isolates. Therefore, the extensive use of antimicrobial agents increases the selective pressure on nosocomial infectious bacterial strains to develop resistance. This selective pressure should be addressed in future studies.
Resistance to SXT and quinolone, which are antimicrobials used to treat Aeromonas infection, has been widely described. Deng et al.[40] reported that at least 18.86% of Aeromonas isolates collected from cultured freshwater animals are resistant to SXT; they also detected sul1 and qnrS in 18.9% and 4.7% of these samples, respectively. Our study obtained an SXT resistance rate of 22.8%, and 12.1% of these strains were positive for both sul1 and qnrS. The detection rate of antimicrobial resistance genes in the clinical isolates was higher than that in the strains identified in healthy individuals. This observation was consistent with the higher antimicrobial resistance rate in clinical isolates. This increased abundance of antimicrobial resistance genes in A. caviae was also consistent with their resistance phenotype, which may explain the increased prevalence of A. caviae isolates in clinical samples. However, this finding should be evaluated in future studies.
-
We analyzed 57 Aeromonas strains isolated from patients and healthy individuals in Ma’anshan City, Anhui Province, China. These samples exhibited a high degree of genetic diversity with the 57 isolates that produced 55 independent STs. The constructed phylogenetic tree with concatenated gyrb-cpn60 sequences divided the 57 isolates into 7 species. Beta-hemolysis, proteolytic activity, and virulence gene analyses revealed that these properties were more common in the clinical isolates than in the isolates from the healthy individuals. Furthermore, A. dhakensis harbored the highest number of virulence genes. Our results suggested that 26.3% of the strains were MDR (≥ 3) positive, and the MDR status was significantly more common in the clinical isolates than in the isolates from the healthy individuals. PMQR, tetA, tetE, ESBL, aminoglycoside resistance, sulfanilamide, and mcr-3 genes were detected in some of the isolates. Their prevalence increased in the clinical isolates. Therefore, the genetic diversity, antimicrobial resistance, and pathogenicity of Aeromonas strains isolated from clinical patients significantly differed from those of the isolates from healthy individuals.
-
The authors have declared that no competing interests exist.
Comparative Study of the Genetic Diversity, Antimicrobial Resistance, and Pathogenicity of Aeromonas Isolates from Clinical Patients and Healthy Individuals
doi: 10.3967/bes2021.062
Funds:
This work was supported by Major Project of the thirteenth Five Year Special for infectious diseases of China [2018ZX10101002]
- Received Date: 2021-01-13
- Accepted Date: 2021-04-07
-
Key words:
- Aeromonas /
- Genetic diversity /
- Multidrug resistance /
- Virulence gene /
- Antimicrobial resistance gene
Abstract:
| Citation: | MENG Shuang, DU Xiao Li, WANG Yong Lu, QU Feng Tian, XIE Gui Lin, ZHOU Hai Jian, HU Jin Rui, QIN Zheng, WANG Yue, KAN Biao, CUI Zhi Gang. Comparative Study of the Genetic Diversity, Antimicrobial Resistance, and Pathogenicity of Aeromonas Isolates from Clinical Patients and Healthy Individuals[J]. Biomedical and Environmental Sciences, 2021, 34(6): 454-464. doi: 10.3967/bes2021.062

|

 Quick Links
Quick Links
 DownLoad:
DownLoad: