Objective To analyze the prevalence and years lived with disability (YLD) from dental caries among children and adolescents and the time trends over the past two decades in Sichuan province, the largest province in west China.Methods Based on the Global Burden of Disease Study 2015 (GBD2015), which systematically assessed the epidemiological characteristics of major diseases and their transitions by country and region from 1990 to 2015, we extracted the estimated results for China. We then used the Bayesian meta-regression method to estimate the sex-and age-specific prevalences and YLDs from dental caries among children and adolescents under 15 years old in Sichuan province and compared them with global and national indicators for the same period.Results In 2015, there were almost 6 million cases of dental caries in children and adolescents (aged < 15 years) in Sichuan province, accounting for 6% of the total cases in China. For children under 5 years, the prevalence of deciduous caries was 55.9%, and the YLDs value was 10.8 per 100, 000, while it was 24.3% and 5.1 per 100, 000 respectively among 5-to 14-year-olds; for those aged 5 to 14 years, the prevalence of permanent caries was 21.5%, and the YLDs value was 11.5 per 100, 000. From 1990 to 2015, the prevalence of dental caries for children under 5 years increased substantially, by 16.2%, and the YLDs increased by 8.7%. Among those aged 5 to 14 years, the prevalence increased and the YLDs decreased.Conclusion Dental caries remains a huge health burden in Western China. In contrast to the global and national data, the trend has increased rapidly over the past 25 years in this region. This work provides suggestions for the prevention and control for oral health in China with the policy of two-child.
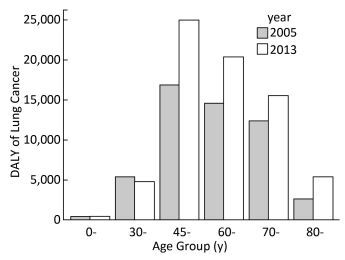
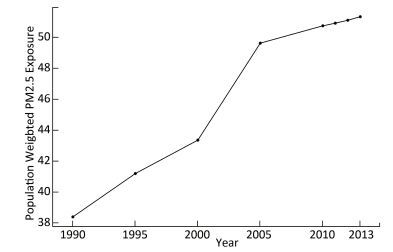
Objective To estimate the lung cancer burden that may be attributable to ambient fine particulate matter (PM2.5) pollution in Guangzhou city in China from 2005 to 2013.Methods The data regarding PM2.5 exposure were obtained from the 'Ambient air pollution exposure estimation for the Global Burden of Disease 2013' dataset at 0.1°×0.1° spatial resolution. Disability-adjusted life years (DALYs) were estimated based on the information of mortality and incidence of lung cancer. Comparative risk analysis and integrated exposure-response function were used to estimate attributed disease burden.Results The population-weighted average concentration of PM2.5 was increased by 34.6% between 1990 and 2013, from 38.37 μg/m3 to 51.31 μg/m3. The lung cancer DALYs in both men and women were increased by 36.2% from 2005 to 2013. The PM2.5 attributed lung cancer DALYs increased from 12105.0 (8181.0 for males and 3924.0 for females) in 2005 to 16489.3 (11291.7 for males and 5197.6 for females) in 2013. An average of 23.1% lung cancer burden was attributable to PM2.5 pollution in 2013.Conclusion PM2.5 has caused serious but under-appreciated public health burden in Guangzhou and the trend deteriorates. Effective strategies are needed to tackle this major public health problem.
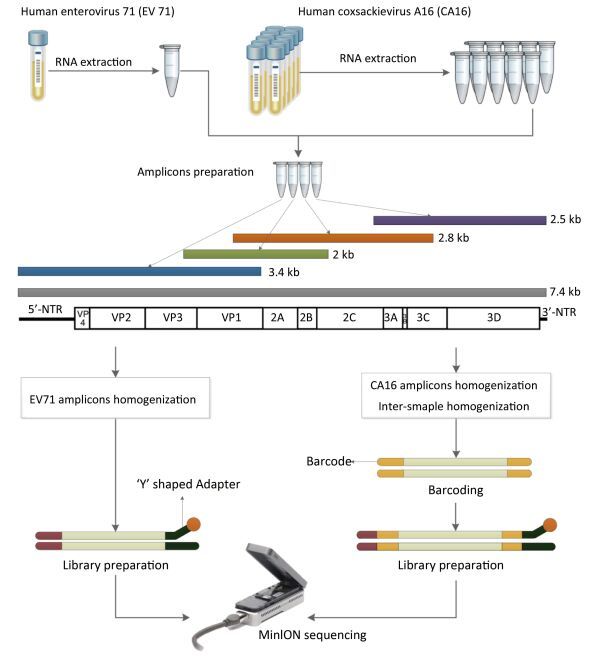
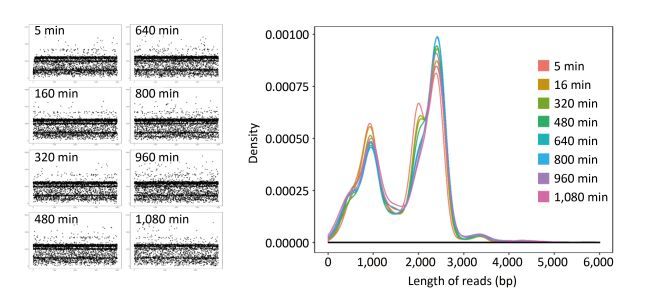
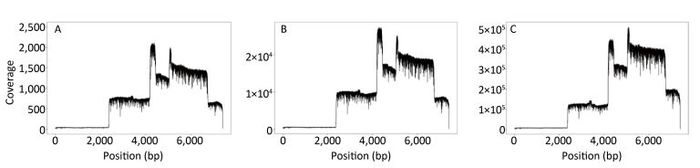
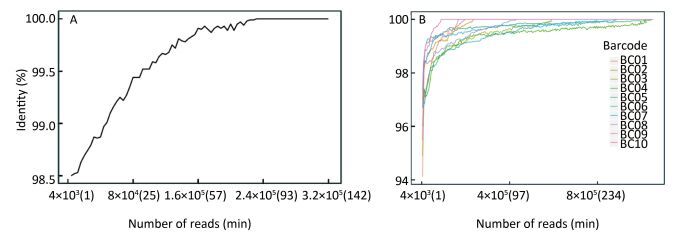
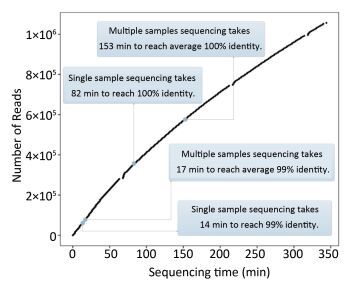
Objective Knowledge of an enterovirus genome sequence is very important in epidemiological investigation to identify transmission patterns and ascertain the extent of an outbreak. The MinION sequencer is increasingly used to sequence various viral pathogens in many clinical situations because of its long reads, portability, real-time accessibility of sequenced data, and very low initial costs. However, information is lacking on MinION sequencing of enterovirus genomes.Methods In this proof-of-concept study using Enterovirus 71 (EV71) and Coxsackievirus A16 (CA16) strains as examples, we established an amplicon-based whole genome sequencing method using MinION. We explored the accuracy, minimum sequencing time, discrimination and high-throughput sequencing ability of MinION, and compared its performance with Sanger sequencing.Results Within the first minute (min) of sequencing, the accuracy of MinION was 98.5% for the single EV71 strain and 94.12%-97.33% for 10 genetically-related CA16 strains. In as little as 14 min, 99% identity was reached for the single EV71 strain, and in 17 min (on average), 99% identity was achieved for 10 CA16 strains in a single run.Conclusion MinION is suitable for whole genome sequencing of enteroviruses with sufficient accuracy and fine discrimination and has the potential as a fast, reliable and convenient method for routine use.
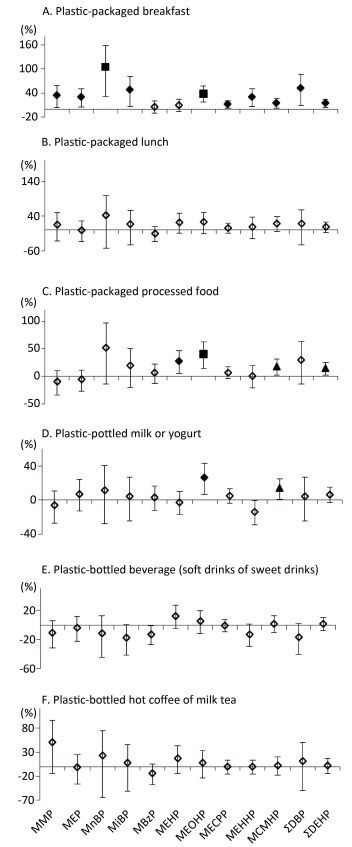
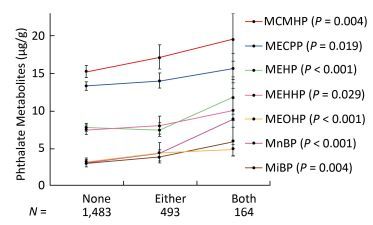
Objective Consuming phthalates may be due to the presence of food contact materials, such as plastic containers. In this study, we investigated the association between plastic container use and phthalate exposure in 2, 140 Shanghai adults.Methods Participants completed a questionnaire on the frequency of using plastic containers in different scenarios in the previous year (e.g., daily, weekly) and on the consumption of plastic-packaged foods in the previous three days (yes or no). Urinary phthalate metabolites were used to assess the association between phthalate exposure and the use of plastic containers.Results The metabolites of di-(2-ethylhexyl) phthalate (DEHP) were the most frequently detected in urine. The results revealed that phthalate exposure was associated with consumption of plastic-packaged breakfast or processed food items in the previous three days. The consumption of these two food items had strong synergistic effects on increasing urinary concentrations of most phthalate metabolites.Conclusion Our results of plastic-packaged breakfast and processed food may be explained by the use of flexible plastic containers, indicating the importance of risk assessment for the application of flexible plastic containers.
Objective Although sleep is one of the most important health-related behavioral factors, the association between night sleep duration and cognitive impairment has not been fully understood. A cross-sectional study was conducted with a random sample of 2, 514 participants (≥ 40 years of age; 46.6% women) in China to examine the association between night sleep duration and cognitive impairment.Methods Night sleep duration was categorized as ≤ 5, 6, 7, 8, or ≥ 9 h per night. Cognitive function was measured using the Mini-Mental State Examination. A multivariate regression analysis was used to analyze the association of night sleep duration with cognitive impairment. A total of 122 participants were diagnosed with cognitive impairment.Results A U-shaped association between night sleep duration and cognitive impairment was found. The odds ratios (95% confidence intervals) of cognitive impairment (with 7 h of daily sleep being considered as the reference) for individuals reporting ≤ 5, 6, 8, and ≥ 9 h were 2.14 (1.20-3.83), 1.13 (0.67-1.89), 1.51 (0.82-2.79), and 5.37 (1.62-17.80), respectively (P ≤ 0.01).Conclusion Short or long night sleep duration was an important sleep-related factor independently associated with cognitive impairment and may be a useful marker for increased risk of cognitive impairment.
Obesity and dental caries are increasing epidemics, especially among children and adolescents. This epidemiological observational cross-sectional study was conducted to assess the possible association between body mass index (BMI) and dental caries among 111, 792 school children and adolescents in Jiangsu Province. We found that 13.14% participants of the study sample were overweight, and 7.37% were obese. The prevalence of dental caries was 12.95% in overweight and 7.89% in obese students. There were significant differences in caries prevalence by sex, region, age group, and BMI. Overweight and obesity statuses were associated with dental caries among the study population. BMI and dental caries present a continuous health problem. Thus, we recommend that oral health promotion be used for caries prevention and control.
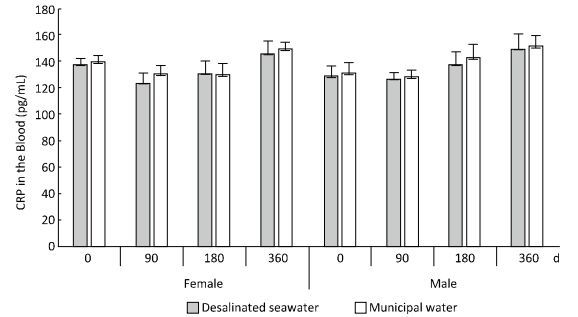
To evaluate the potential risk of arteriosclerosis caused by desalinated seawater, Wistar rats were provided desalinated seawater over a 1-year period, and blood samples were collected at 0, 90, 180, and 360 days. Blood calcium, magnesium, and arteriosclerosis-related indicators were investigated. Female rats treated with desalinated seawater for 180 days showed lower magnesium levels than the control rats (P < 0.05). The calcium and magnesium levels in female rats and the magnesium level in male rats were lower than the levels in the controls, following treatment with desalinated seawater for 360 days (P < 0.05). Blood levels of arteriosclerosis-related lipid peroxidation indicators and C-reactive protein (CRP) in the treatment group did not differ from those in the controls. The levels of lipid peroxidation indicators and CRP in rats were not significantly affected by drinking desalinated seawater, and no increase in risk of arteriosclerosis was observed.
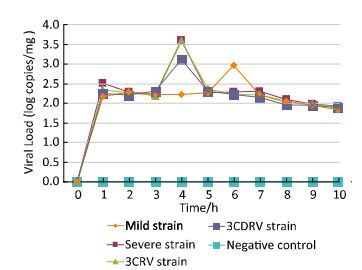
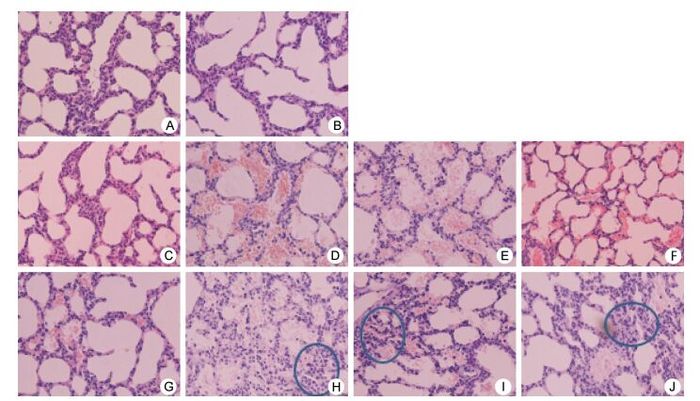
Enterovirus 71 is a neuroinvasive virus that is associated with severe neurological complications. We had earlier suggested that the replication capacity of a severe strain was higher than that of a mild strain. The recombinant 3CRV and 3CDRV virus strains were successfully rescued in our previous study. In the present study, we found no difference in virulence between 3CRV and severe strains. However, the capacity of replication and to cause cell injury of 3CDRV strain decreased in vitro, especially at 39.5 ℃. Replacement of 3CD region in the severe strain led to milder symptoms, less body weight loss, and lower viral load in ICR mice. Histopathological findings indicated less severe injury in mice infected with 3CDRV strain. This study suggests that the 3CD region contributes to the attenuation of the severe strain, including its replication capacity and temperature sensitivity.
To establish a genetic susceptibility assessment model of lung cancer risk potentially associated with polycyclic aromatic hydrocarbon (PAH) inhalation exposure among non-smokers in China, a total of 143 patients with lung adenocarcinoma and 143 cancer-free individuals were recruited. Sixty-eight genetic polymorphisms in 10 pathways related to PAH metabolism and tumorigenesis were selected and examined. It was observed that 3 genetic polymorphisms, along with 10 additional genetic polymorphisms via gene-gene interactions, significantly influenced lung cancer risk potentially associated with PAH inhalation exposure. Most polymorphisms were associated with PAH metabolism. According to the established genetic susceptibility score (GSS), lung cancer risk increased with a rise in the GSS level, thereby indicating a positive dose-response relationship.
Occupational carbon monoxide (CO) poisoning related to diesel motor fumes in an air-raid shelter (ARS) was first identified in Jinan City, China, in June 2015. A total of 17 cases were identified, including 14 possible cases of firemen and 3 confirmed cases of water channel clean-up workers. The overall attack rate (AR) of firemen was 42% (14/33). The firemen had a significantly higher AR with a longer exposure and more protracted time of rescue in the ARS (P < 0.05). All the cases stated that they did not realize the potentially high level of exposure to CO in the ARS. CO poisoning posed a risk to both patients and service providers. Occupational safety and health education should be promoted and enforced in all workplaces where CO sources exist.





























 Quick Links
Quick Links