
2017 Vol. 30, No. 11






2017, 30(11): 783-791.
doi: 10.3967/bes2017.106


2017, 30(11): 792-801.
doi: 10.3967/bes2017.107



2017, 30(11): 802-810.
doi: 10.3967/bes2017.108
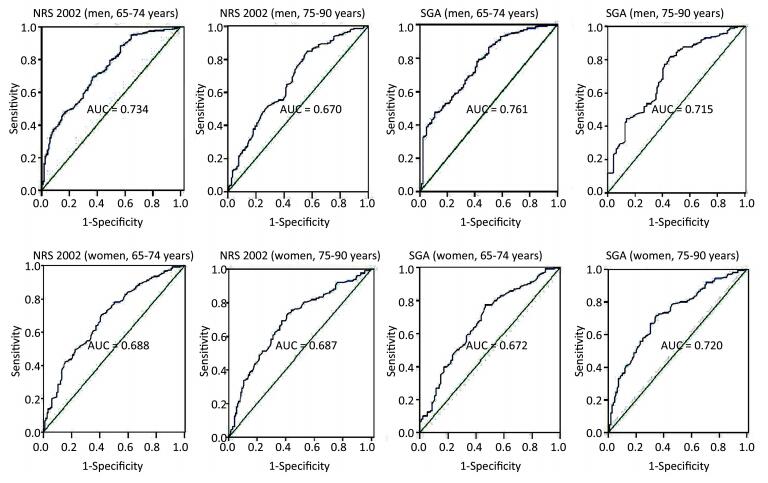

2017, 30(11): 811-824.
doi: 10.3967/bes2017.109


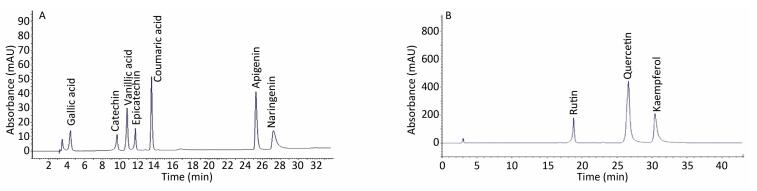


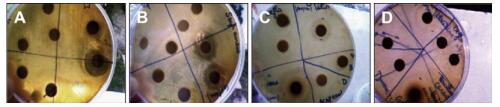
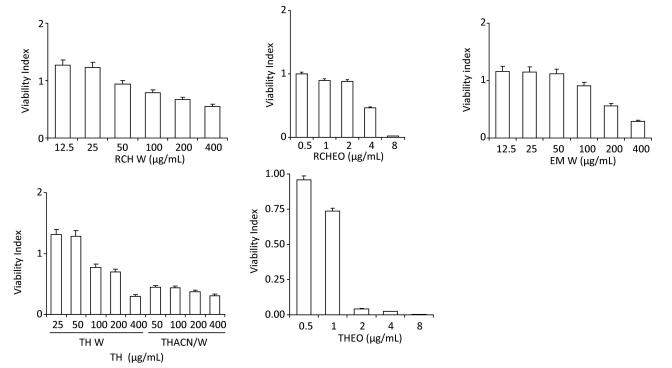
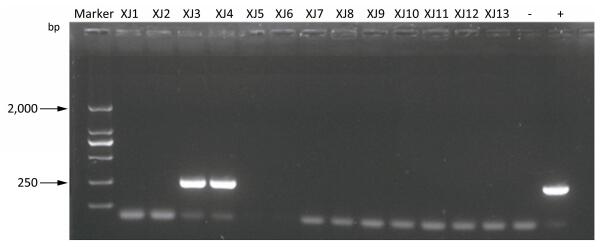
This study assesses the causes and prevention measures of rabies through epidemiological investigation and analysis. A field epidemiological survey was conducted to investigate a case of rabies by fox bite. The onset of symptoms began 50 days after the bite. The patient did not receive standard treatment, rabies vaccination, or rabies immunoglobulin injection. The fox was killed on the spot. Saliva and pre-death blood samples were collected at different periods, and only blood RT-PCR tests yielded positive results. Wild fox bite is a major risk factor of rabies infection in Xinjiang Province, China.
Population-based cancer registration data were collected to estimate the cancer incidence and mortality in Wuwei, Hexi Corridor Region, China in 2018. We used the 2011-2013 data to predict the number of new cases and deaths in 2018 and the 2003-2013 data to analyze trends in cancer incidence and mortality. The goal is to enable cancer prevention and control directions. Our results indicated that stomach cancer is the most common cancer. For all cancers combined, the incidence and mortality rates showed significantly increasing trends (+2.63% per year; P < 0.05 and +1.9% per year; P < 0.05). This study revealed a significant cancer burden among the population of this area. Cancer screening and prevention should be performed after an epidemiological study of the cause of the cancer is completed.

2017, 30(11): 834-840.
doi: 10.3967/bes2017.112


The aim of the present study is to analyze the global research trend of radiation-responsive genes and identify the highly reproducible radiation-responsive genes. Bibliometric methods were applied to analyze the global research trend of radiation-responsive genes. We found 79 publications on radiation-responsive genes from 2000 to 2017. A total of 35 highly reproducible radiation-responsive genes were identified. Most genes are involved in response to DNA damage, cell proliferation, cell cycle regulation, and DNA repair. The p53 signal pathway was the top enriched pathway. The expression levels of 18 genes in human B lymphoblastoid cell line (AHH-1) cells were significantly up-regulated in a dose-dependent manner at 24 h after exposure to 0-5 Gy 60Co γ-ray irradiation. Our results indicate that developing a gene expression panel with the 35 high reproducibility radiation-responsive genes may be necessary for qualitative and quantitative assessment after exposure.

2017, 30(11): 841-845.
doi: 10.3967/bes2017.113
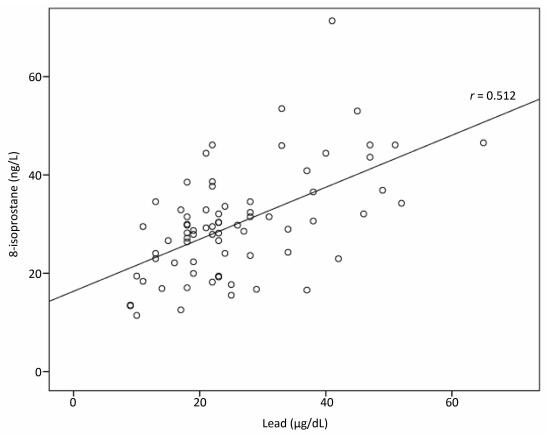
We aimed to investigate the short-term correlation between blood lead levels and oxidative stress generation in coal miners. The study involved 94 male coal miners from the Velenje Coal mine, arranged into four groups: three groups according to the number of consecutive working days, and a fourth control group. Miners who worked for three consecutive days had higher blood levels of lead and 8-isoprostane than the control group (P < 0.001). Correlation between lead and 8-isoprostane was of medium strength (r = 0.512, P < 0.001). Short-term lead environmental exposure can potentially harmful and should be considered when formulating improvements in working processes.

2017, 30(11): 846-850.
doi: 10.3967/bes2017.114

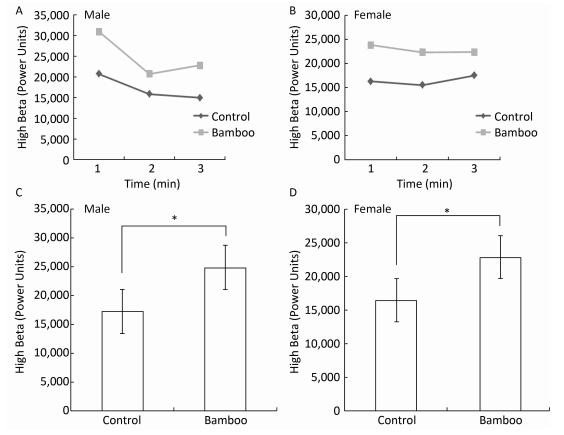
The present study was conducted to clarify the psychophysiological relaxation effects of viewing bamboo on university students. Forty healthy Chinese participants enrolled in this study to clarify the psychophysiological relaxation effects of viewing bamboo. The effects of visual stimulation using a pot both with and without a bamboo were recorded by measuring the student's blood pressure, EEG and STAI. We observed that viewing bamboo plants resulted in significantly lower systolic (female, P < 0.001; male, P < 0.001; P < 0.05) and diastolic (female, P < 0.001; male, P < 0.001; P < 0.05) blood pressures, but no changes in the pulse rate (female, P = 0.09; male, P = 0.07; P > 0.05) were observed. The results of the EEG analysis indicated brainwave variation (all P < 0.05) and lower anxiety scores (P < 0.01) after 3 min of viewing bamboo compared with the control. These findings indicate that visual stimulation with bamboo plants induced psychophysiological relaxation effects on adults.
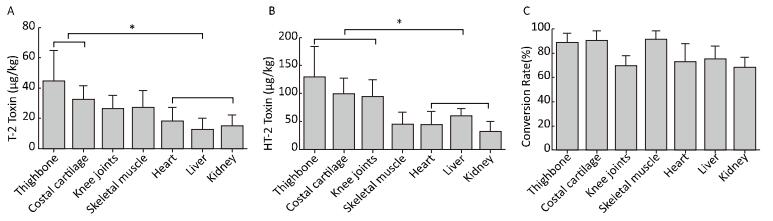
Twelve healthy rats were divided into the T-2 toxin group receiving gavage of 1 mg/kg T-2 toxin and the control group receiving gavage of normal saline. Total relative concentrations of T-2 toxin and HT-2 toxin in the skeletal system (thighbone, knee joints, and costal cartilage) were significantly higher than those in the heart, liver, and kidneys (P < 0.05). The relative concentrations of T-2 toxin and HT-2 toxin in the skeletal system (thighbone and costal cartilage) were also significantly higher than those in the heart, liver, and kidneys. The rats administered T-2 toxin showed rapid metabolism compared with that in rats administered HT-2 toxin, and the metabolic conversion rates in the different tissues were 68.20%-90.70%.
2017, 30(11): 855-861.
doi: 10.3967/bes2017.116













 Quick Links
Quick Links