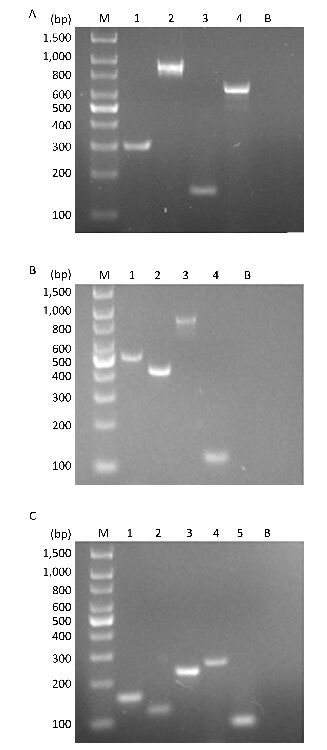
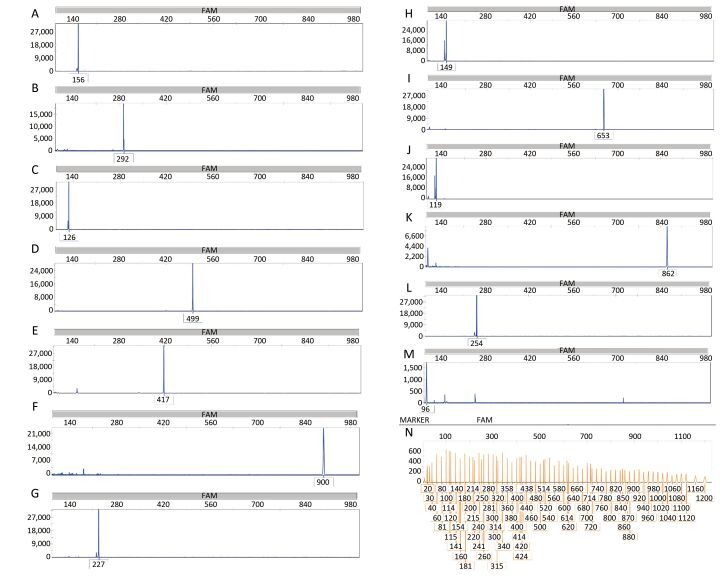
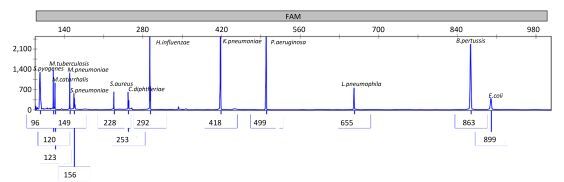
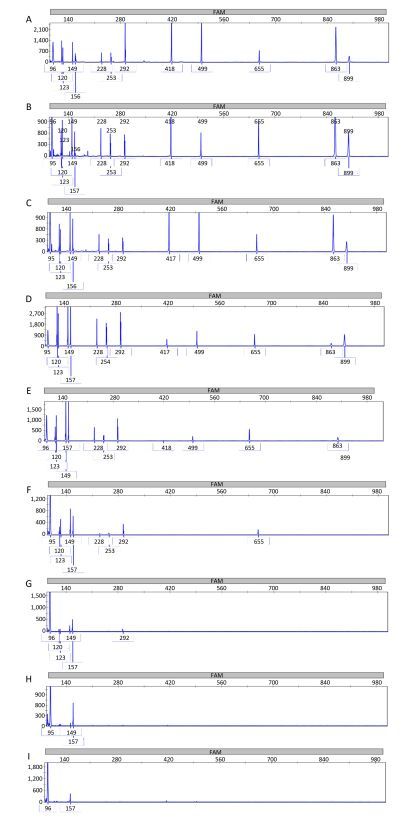
Objective Lower respiratory tract infections continue to pose a significant threat to human health. It is important to accurately and rapidly detect respiratory bacteria. To compensate for the limits of current respiratory bacteria detection methods, we developed a combination of multiplex polymerase chain reaction (PCR) and capillary electrophoresis (MPCE) assay to detect thirteen bacterial pathogens responsible for lower respiratory tract infections, including Streptococcus pneumoniae, Haemophilus influenzae, Moraxella catarrhalis, Pseudomonas aeruginosa, Klebsiella pneumoniae, Escherichia coli, Staphylococcus aureus, Mycoplasma pneumoniae, Legionella spp., Bordetella pertussis, Mycobacterium tuberculosis complex, Corynebacterium diphtheriae, and Streptococcus pyogenes.Methods Three multiplex PCR reactions were built, and the products were analyzed by capillary electrophoresis using the high-throughput DNA analyzer. The specificity of the MPCE assay was examined and the detection limit was evaluated using DNA samples from each bacterial strain and the simulative samples of each strain. This assay was further evaluated using 152 clinical specimens and compared with real-time PCR reactions. For this assay, three nested-multiplex-PCRs were used to detect these clinical specimens.Results The detection limits of the MPCE assay for the 13 pathogens were very low and ranged from 10-7 to 10-2 ng/μL. Furthermore, analysis of the 152 clinical specimens yielded a specificity ranging from 96.5%-100.0%, and a sensitivity of 100.0% for the 13 pathogens.Conclusion This study revealed that the MPCE assay is a rapid, reliable, and high-throughput method with high specificity and sensitivity. This assay has great potential in the molecular epidemiological survey of respiratory pathogens.
Objective To characterize carbapenem (CPM)-non-susceptible Klebsiella pneumoniae (K. pneumoniae) and carbape-nemase produced by these strains isolated from Beijing Children's Hospital based on a five-year surveillance.Methods The Minimal Inhibition Concentration values for 15 antibiotics were assessed using the Phonix100 compact system. PCR amplification and DNA sequencing were used to detect genes encoding carbapenemases. WHONET 5.6 was finally used for resistance analysis.Results In total, 179 strains of CPM-non-susceptible K. pneumoniae were isolated from January, 2010 to December, 2014. The rates of non-susceptible to imipenem and meropenem were 95.0% and 95.6%, respectively. In the 179 strains, 95 (53.1%) strains carried the blaIMP gene, and IMP-4 and IMP-8 were detected in 92 (96.8%) and 3 (3.2%) IMP-producing isolates, respectively. 65 (36.3%) strains carried the blaNDM-1 gene. 6 (3.4%) strains carried the blaKPC gene, and KPC-2 were detected in 6 KPC-producing isolates. In addition, New Delhi-Metallo-1 (NDM-1) producing isolates increased from 7.1% to 63.0% in five years and IMP-4 producing isolates decreased from 75.0% to 28.3%.Conclusion High frequencies of multiple resistances to antibiotics were observed in the CPM-non-susceptible K. pneumoniae strains isolated from Beijing Children's Hospital. The production of IMP-4 and NDM-1 metallo-β-lactamases appears to be an important mechanism for CPM-non-susceptible in K. pneumoniae.
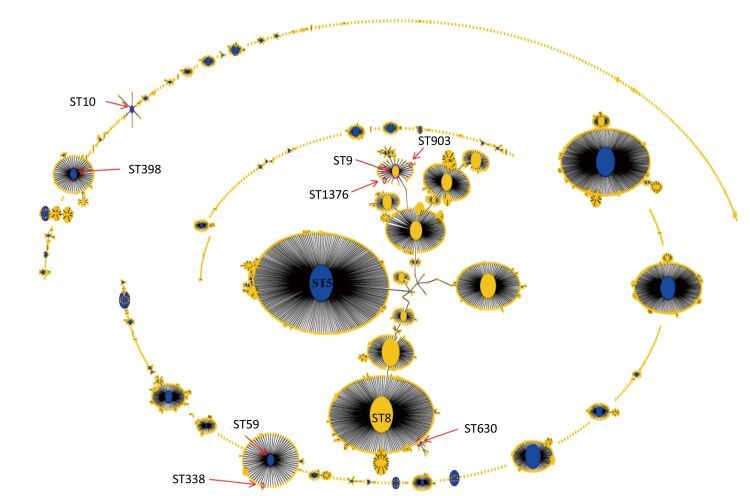
Objective To investigate the genotypic diversity of Methicillin-resistant Staphylococcus aureus (MRSA) isolated from pigs and retail foods from different geographical areas in China and further to study the routes and rates of transmission of this pathogen from animals to food.Methods Seventy-one MRSA isolates were obtained from pigs and retail foods and then characterized by multi-locus sequencing typing (MLST), spa typing, multiple-locus variable number of tandem repeat analysis (MLVA), pulsed-field gel electrophoresis (PFGE), and antimicrobial susceptibility testing.Results All isolated MRSA exhibited multi-drug resistance (MDR). Greater diversity was found in food-associated MRSA (7 STs, 8 spa types, and 10 MLVA patterns) compared to pig-associated MRSA (3 STs, 1 spa type, and 6 MLVA patterns). PFGE patterns were more diverse for pig-associated MRSA than those of food-associated isolates (40 vs. 11 pulse types). Among the pig-associated isolates, CC9-ST9-t899-MC2236 was the most prevalent clone (96.4%), and CC9-ST9-t437-MC621 (20.0%) was the predominant clone among the food-associated isolates. The CC9-ST9 isolates showed significantly higher antimicrobial resistance than other clones. Interestingly, CC398-ST398-t034 clone was identified from both pig-and food-associated isolates. Of note, some community-and hospital-associated MRSA strains (t030, t172, t1244, and t4549) were also identified as food-associated isolates.Conclusion CC9-ST9-t899-MC2236-MDR was the most predominant clone in pigs, but significant genetic diversity was observed in food-associated MRSA. Our results demonstrate the great need for improved surveillance of MRSA in livestock and food and effective prevention strategies to limit MDR-MRSA infections in China.
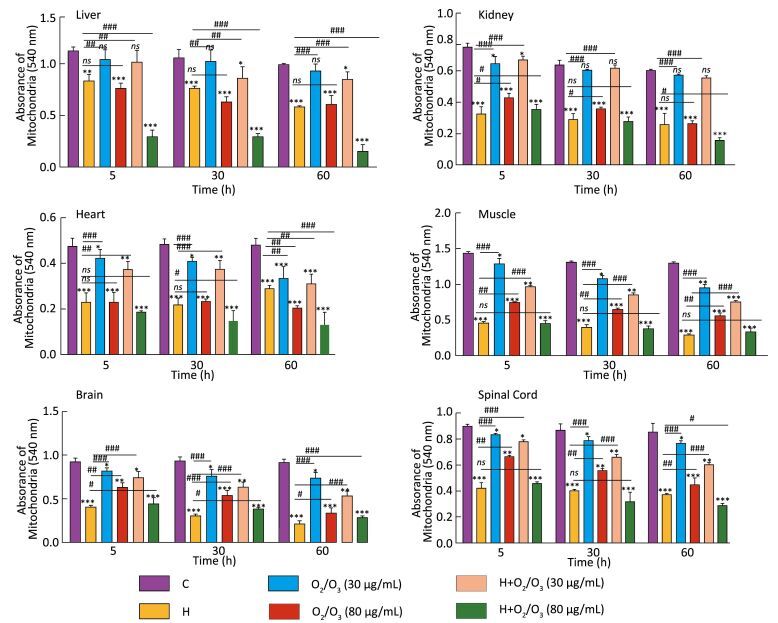
Objective Scorpion (Hemiscorpius lepturus) stings are a public health concern in Iran, particularly in south and southwestern regions of Iran. The gold standard for the treatment of a scorpion sting is anti-venom therapy. However, immunotherapy can have serious side effects, such as anaphylactic shock (which can sometimes even lead to death). The aim of the current study was to demonstrate the protective effect of ozone against toxicity induced by Hemiscorpius lepturus (H. lepturus) venom in mice.Methods Eight hours after the injection of ozone to the experimental design groups, the male mice were decapitated and mitochondria were isolated from five different tissues (liver, kidney, heart, brain, and spinal cord) using differential ultracentrifugation. Then, assessment of mitochondrial parameters including mitochondrial reactive oxidative species (ROS) production, mitochondrial membrane potential (MMP), ATP level, and the release of cytochrome c from the mitochondria was performed.Results Our results showed that H. lepturus venom-induced oxidative stress is related to ROS production and MMP collapse, which is correlated with cytochrome c release and ATP depletion, indicating the predisposition to the cell death signaling.Conclusion In general, ozone therapy in moderate dose can be considered as clinically effective for the treatment of H. lepturus sting as a protective and antioxidant agent.
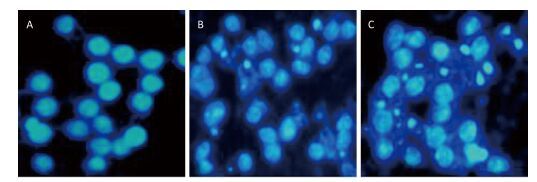
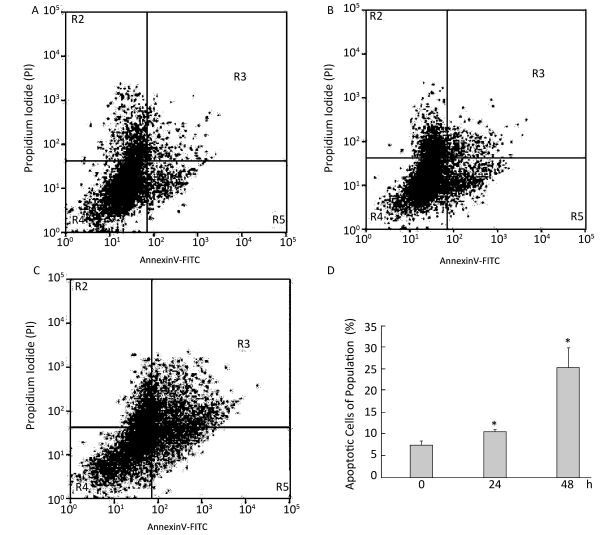
Objective We investigated the role of endoplasmic reticulum stress (ERS) in silica-induced apoptosis in alveolar macrophages in vitro.Methods RAW264.7 cells were incubated with 200 μg/mL silica for different time periods. Cell viability was assayed by the MTT assay. Cell apoptosis was evaluated by DAPI staining, flow cytometry analysis, and Western blot analysis of caspase-3. Morphological changes in the endoplasmic reticulum were observed by transmission electron microscopy. The expression of ERS markers binding protein (BiP) and CCAAT-enhancer-binding protein homologous protein (CHOP) was examined by Western blotting and real-time PCR. As an inhibitor of ERS, 4-phenylbutyric acid (4-PBA) was used in the experiments.Results Silica exposure induced nuclear condensation and caspase-3 expression in RAW264.7 cells. The number of apoptotic cells increased after silica exposure in a time-dependent manner. Silica treatment induced expansion of the endoplasmic reticulum. In addition, the expression of BiP and CHOP increased in silica-stimulated cells. Furthermore, 4-PBA treatment inhibited silica-induced endoplasmic reticulum expansion and the expression of BiP and CHOP. Moreover, 4-PBA treatment attenuated nuclear condensation, reduced apoptotic cells, and downregulated caspase-3 expression in silica-stimulated cells.Conclusion Silica-induced ERS is involved in the apoptosis of alveolar macrophages.
Infections by Cronobacter spp. are hazardous to infants since they can lead to neonatal meningitis, bacteremia, and necrotizing enterocolitis. Cronobacter spp. are frequently resistant to β-lactam derivatives, macrolides, and aminoglycosides. In addition, multi-resistant strains have also been detected. In China, the isolation rate of Cronobacter spp. from commercial powdered infant formula (PIF) or follow-up formula (FUF) is relatively high. Nevertheless, clinical cases of Cronobacter infection have been ignored to date. Here we describe two cases of Cronobacter infection detected at the Wuhan Women and Children Medical Care Center Hospital (Wuhan City, China). We provide the genomic analysis of the isolates and the antibiotic-resistance profiles of the two strains. The Cronobacter strains identified in this study were not susceptible to third-generation cephalosporins, aminoglycoside, and/or trimethoprim-sulfamethoxazole. Whole genome sequencing revealed various genes known to encode antibiotic resistance. Future studies are needed to determine whether the genes predicted in this study are functional. As with Enterobacter spp., the antibiotic resistance of Cronobacter is a serious issue that requires more attention.
We used a proteomic approach to identify IbpA in Cronobacter sakazakii (C. sakazaki), which is related to heat tolerance in this strain. The abundance of IbpA in C. sakazakii strains strongly increased after heat shock. C. sakazakii CMCC 45402 ibpA deletion mutants were successfully constructed. The C. sakazakii CMCC 45402 ΔibpA and wild-type strains could not be distinguished based on colony morphology on LB agar plates or biochemical assays. The growth of the C. sakazakii CMCC 45402 ΔibpA mutant in heat shock conditions was indistinguishable from that of the isogenic wild-type, but showed greater heat resistance than E. coli O157:H7 strain CMCC 44828. This study suggests that the absence of a single ibpA gene has no obvious effect on the phenotype or heat resistance of the strain C. sakazakii CMCC 45402.
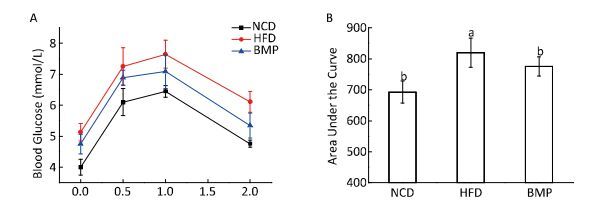
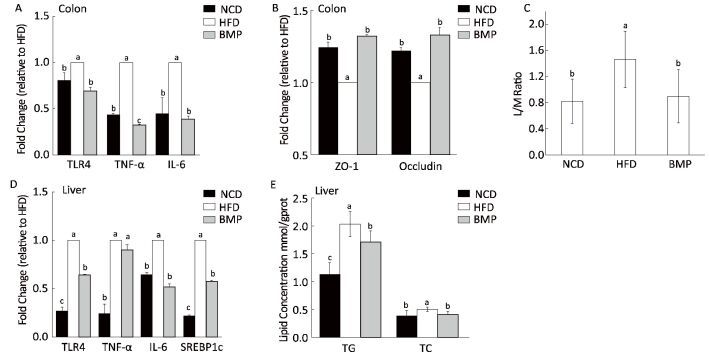
This study explored how bitter melon powder (BMP) alters the colonic microenvironment during the development of obesity-associated fatty liver in rats. We observed that BMP effectively inhibited the body weight gain and lipid accumulation in the liver, ameliorated glucose intolerance, and increased the colon weight after an 8-week treatment compared to that in the high-fat diet (HFD) group. BMP significantly decreased fecal water toxicity towards HT-29 cells, as revealed by the cell counting kit (CCK)-8 assay results, and the mRNA expression of Toll-like receptor 4 (TLR4) in colon mucosa. Additionally, gut permeability in the BMP group was restored to normal levels. Finally, BMP alleviated the inflammatory state of the rat colon mucosa and liver tissues as well as the systemic inflammation.



































 Quick Links
Quick Links