Objective Low-density lipoprotein cholesterol (LDL-C) has been well known as the risk factor of coronary artery disease (CAD). However, the role of lipoprotein (a)[Lp(a)] in the development of CAD is of great interest but still controversial. Thus, we aim to explore the effect of Lp(a) on predicting the presence and severity of CAD in Chinese untreated patients, especially in combination with LDL-C.Methods We consecutively recruited 1, 980 non-treated patients undergoing coronary angiography, among which 1,162 patients were diagnosed with CAD. Gensini score (GS) was used to assess the severity of CAD. Lp(a) was measured by immunoturbidimetric method.Results Patients with CAD had higher level of LDL-C and Lp(a) compared with non-CAD (P < 0.05). Multivariable logistic regression revealed that Lp(a) > 205 mg/L (highest tertile) predicted 1.437-fold risk for CAD (95% CI:1.108-1.865, P=0.006) and 1.480-fold risk for high GS (95% CI:1.090-2.009, P=0.012) respectively. Interestingly, concomitant elevated level of Lp(a) and LDL-C conferred the highest risk for both presence[OR=1.845, 95% CI:1.339-2.541, P < 0.001] and severity[OR=1.736, 95% CI:1.188-2.538, P=0.004] of CAD.Conclusion Lipoprotein (a) is a useful marker for predicting the presence and severity of CAD, especially combined with LDL-C.
Objective To investigate the prevalence and possible factors influencing metabolic syndrome in people from Guizhou Province and to explore the predictive value of the fat-to-muscle ratio in diagnosing metabolic syndrome.Methods A multistage stratified sampling method was used in this cross-sectional study of 20-80 years old Han and Bouyei populations from Guizhou Province, southwestern China, from October-December 2012. The study included 4, 553 cases of metabolic syndrome, that was defined according to 2005 International Diabetes Federation criteria. The receiver operating characteristic curve was used for determining the sensitivity, specificity, and predictive ability of the fat-to-muscle ratio for the diagnosis of metabolic syndrome.Results The age-standardized prevalence of metabolic syndrome was 11.38% (men:9.76%; women:12.72%) for Han and 4.78% (men:4.43%; women:5.30%) for Bouyei populations. In Guizhou Province, the cut-off value for the men fat-to-muscle ratio was 0.34, the area under the curve was 0.95, and the sensitivity and specificity were 0.94 and 0.85, respectively. The cut-off value for the women fat-to-muscle ratio was 0.55, the area under the curve was 0.91, and the sensitivity and specificity were 0.93 and 0.79, respectively.Conclusion The fat-to-muscle ratio is highly predictive of metabolic syndrome in Guizhou Province, and a useful reference indicator.
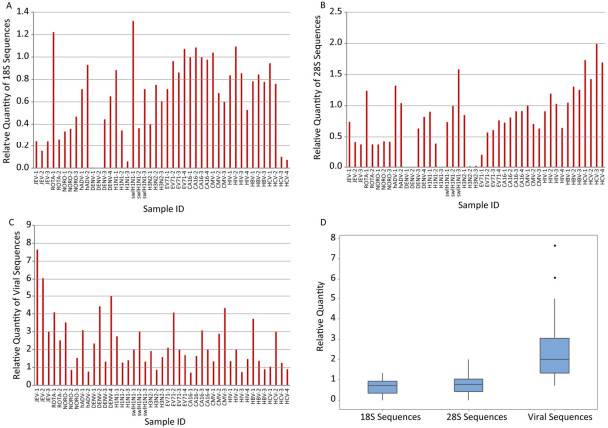
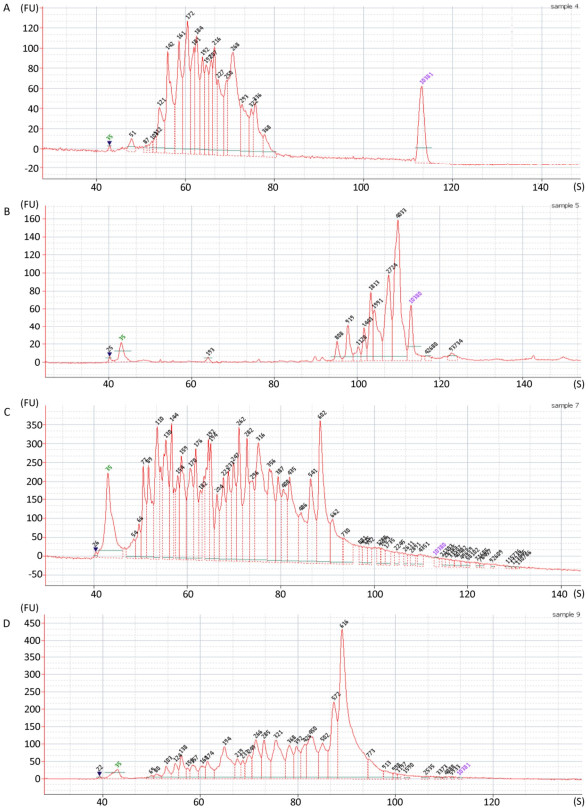
Objective Unbiased next generation sequencing (NGS) is susceptible to interference from host or environmental sequences. Consequently, background depletion and virome enrichment techniques are usually needed for clinical samples where viral load is much lower than background sequences.Methods A viral Sequence Independent Targeted Amplification (VSITA) approach using a set of non-ribosomal and virus-enriched octamers (V8) was developed and compared with traditionally used random hexamers (N6). Forty-five archived clinical samples of different types were used in parallel to compare the V8 and N6 enrichment performance of viral sequences and removal performance of ribosomal sequences in the step of reverse transcription followed by quantitative PCR (qPCR). Ten sera samples from patients with fever of unknown origin and 10 feces samples from patients with diarrhea of unknown origin were used in comparison of V8 and N6 enrichment performance following NGS analysis.Results A minimum 30 hexamers matching to viral reference sequences (sense and antisense) were selected from a dataset of random 4, 096 (46) hexamers (N6). Two random nucleotides were added to the 5' end of the selected hexamers, and 480 (30×42) octamers (V8) were obtained. In general, VSITA approach showed higher enrichment of virus-targeted cDNA and enhanced ability to remove unwanted ribosomal sequences in the majorities of 45 predefined clinical samples. Moreover, VSITA combined with NGS enabled to detect not only more viruses but also achieve more viral reads hit and higher viral genome coverage in 20 clinical samples with diarrhea or fever of unknown origin.Conclusion The VSITA approach designed in this study is demonstrated to possess higher sensitivity and broader genome coverage than traditionally used random hexamers in the NGS-based identification of viral pathogens directly from clinical samples.
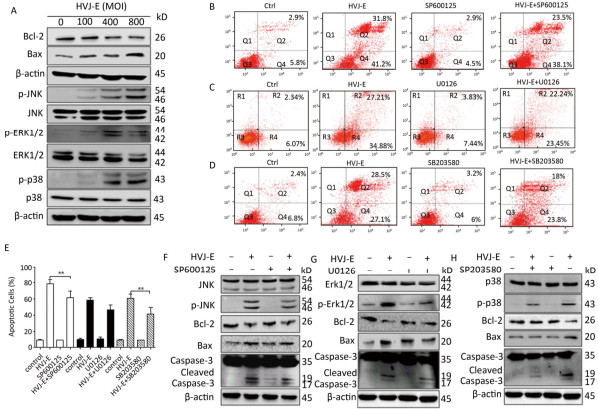
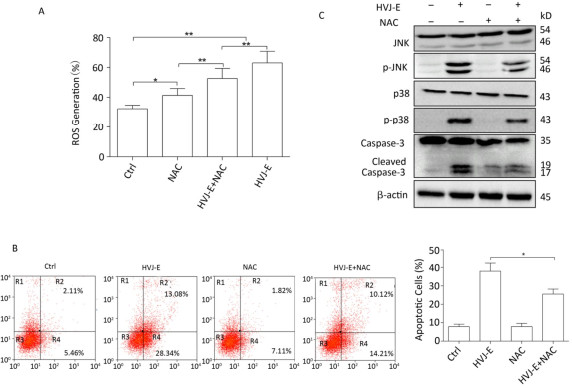
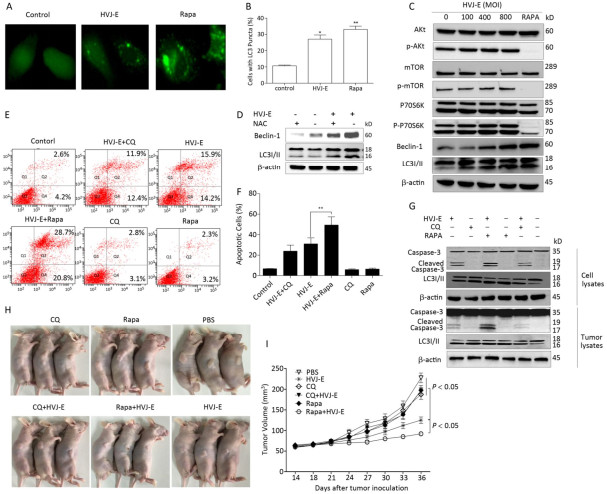
Objective The current study aims to investigate the effect of Hemagglutinating virus of Japan envelope (HVJ-E) on induction of apoptosis and autophagy in human prostate cancer PC3 cells, and the underlying mechanisms.Methods PC3 cells were treated with HVJ-E at various multiplicity of infection (MOI), and the generated reactive oxygen species (ROS), cell viability, apoptosis, and autophagy were detected, respectively. Next, the role of ROS played in the regulation of HVJ-E-induced apoptosis and autuphagy in PC3 cells were analysed. In the end, the relationship between HVJ-E-induced apoptosis and autuophagy was investigated by using rapamycin and chloroquine.Results Flow cytometry assay revealed that HVJ-E treatment induced dose-dependent apoptosis and that the JNK and p38 MAPK signaling pathways were involved in HVJ-E-induced apoptosis in PC3 cells. In addition, HVJ-E was able to induce autophagy in PC3 cells via the class Ⅲ PI3K/beclin-1 pathway. The data also implyed that HVJ-E-triggered autophagy and apoptosis were ROS dependent. When ROS was blocked with N-acetylcysteine (NAC), HVJ-E-induced LC3-Ⅱ conversion and apoptosis were reversed. Interestingly, HVJ-E-induced apoptosis was significantly increased by an inducer of autophagy, rapamycin pretreatment, both in vitro and in vivo.Conclusion HVJ-E exerts anticancer effects via autophagic cell death in prostate cancer cells.
Objective Macrolide susceptibility and drug resistance mechanisms of clinical non-tuberculous mycobacteria (NTM) isolates were preliminarily investigated for more accurate diagnosis and treatment of the infection in China.Methods Four macrolides, including clarithromycin (CLAR), azithromycin (AZM), roxithromycin (ROX), and erythromycin (ERY), were used to test the drug susceptibility of 310 clinical NTM isolates from six provinces of China with the broth microdilution method. Two resistance mechanisms, 23S rRNA and erm, were analyzed with nucleotide sequence analysis.Results Varied effectiveness of macrolides and species-specific resistance patterns were observed. Most Mycobacterium abscessus subsp. massiliense were susceptible and all M. fortuitum were highly resistant to macrolides. All the drugs, except for erythromycin, exhibited excellent activities against slow-growing mycobacteria, and drug resistance rates were below 22.2%. Only four highly resistant strains harbored 2, 058/2, 059 substitutions on rrl and none of other mutations were related to macrolide resistance. G2191A and T2221C on rrl were specific for the M. abscessus complex (MABC). Seven sites, G2140A, G2210C, C2217G, T2238C, T2322C, T2404C, and A2406G, were specifically carried by M. avium and M. intracellulare. Three sites, A2192G, T2358G, and A2636G, were observed only in M. fortuitum and one site G2152A was specific for M. gordonae. The genes erm(39) and erm(41) were detected in M. fortuitum and M. abscessus and inducible resistance was observed in relevant sequevar.Conclusion The susceptibility profile of macrolides against NTM was demonstrated. The well-known macrolide resistance mechanisms, 23S rRNA and erm, failed to account for all resistant NTM isolates, and further studies are warranted to investigate macrolide resistance mechanisms in various NTM species.



























 Quick Links
Quick Links