
-
Cerebral small vessel disease (CSVD) is a common age-related neuropathological condition that accounts for approximately 25%–50% of clinical stroke cases[1]. Its radiological features can be identified on magnetic resonance imaging (MRI) and include recent small subcortical infarcts, lacunes, white matter hyperintensities (WMH), perivascular spaces (PVS), cerebral microbleeds (CMBs), cortical cerebral microinfarcts, and brain atrophy. CSVD is also a major contributor to cognitive impairment (CI) in older adults[2]. Therefore, clarifying the mechanisms through which CSVD impairs cognitive function and identifying novel therapeutic targets are crucial for preventing disease onset and progression.
Accumulating evidence suggests that inflammation may be the main cause of CSVD-related CI[3]. Excessive microglial activation in a chronic hypoxic and ischemic environment lead to the release of large amounts of inflammatory cytokines. These cytokines have deleterious effects on brain tissue, endothelial function, and the cerebrovascular network, thereby increasing the risk of CI[4]. A previous case-control study demonstrated increased blood–brain barrier permeability, assessed by dynamic contrast-enhanced MRI, and enhanced microglial activation, assessed by [11C]PK11195 positron emission tomography, in the white matter of patients with sporadic CSVD; both findings were associated with blood inflammatory biomarkers[5].
Because blood sampling is more accessible than neuroimaging, peripheral inflammatory markers have been widely investigated in CSVD. C-reactive protein, a commonly used marker of inflammation, has been associated with WMH severity[6], increased CMB burden[7,8], and vascular CI[9]. Alterations in peripheral immune cell phenotypes and functions have also been linked to CSVD. A longitudinal study reported that the frequency of CD14+CD16+monocytes and cytokine production, including IL-8, IL-6, and IL-17, were associated with the development of CSVD[10]. In patients with vascular dementia, abnormal level of cytokines such as interferon-γ, tumor necrosis factor-α (TNF-α), and monocyte chemoattractant protein-1 have also been observed[11], along with elevated concentrations of IL-6 and IL-8 in cerebrospinal fluid[12]. Importantly, higher levels of inflammatory markers in both cerebrospinal fluid and plasma have been associated with CI[11]. Experimental studies further suggest that anti-inflammatory therapies can reduce inflammatory cytokine levels in cerebral white matter, promote myelin regeneration and repair, and improve CI[13,14]. Despite these findings, the role of inflammation is as a primary or secondary cause of CSVD remains unclear. Moreover, the specific circulating inflammatory cytokines that mediate the pathway from CSVD to CI have not been fully identified.
Mendelian randomization (MR) is a method that explores evidence of a causal association between exposures and outcomes by employing single nucleotide polymorphisms (SNPs) as genetic instrumental variables (IVs)[15]. Because genetic variants are randomly allocated during meiosis, MR analyses are less susceptible to environmental confounding and reverse causation than conventional observational studies. Consequently, MR provides more robust evidence regarding causal relationship between risk factors and disease outcomes[16,17]. Using MR methods may therefore help clarify the hypothesized causal relationships among CSVD, inflammation, and CI, identify early peripheral biomarkers, and uncover novel therapeutic targets.
Given the role of inflammation in CSVD and its cognitive consequences, we first used two-sample MR to reveal the causal relationship between CSVD and CI. We then performed a two-step, two-sample MR analysis to identify circulating inflammatory cytokines that mediate the association between CSVD and CI. Finally, to further explore the mechanism through which circulating inflammatory cytokines affect cognitive function, we conducted an additional two-sample MR analysis to examine the causal effects of these cytokines on brain structural alterations.
-
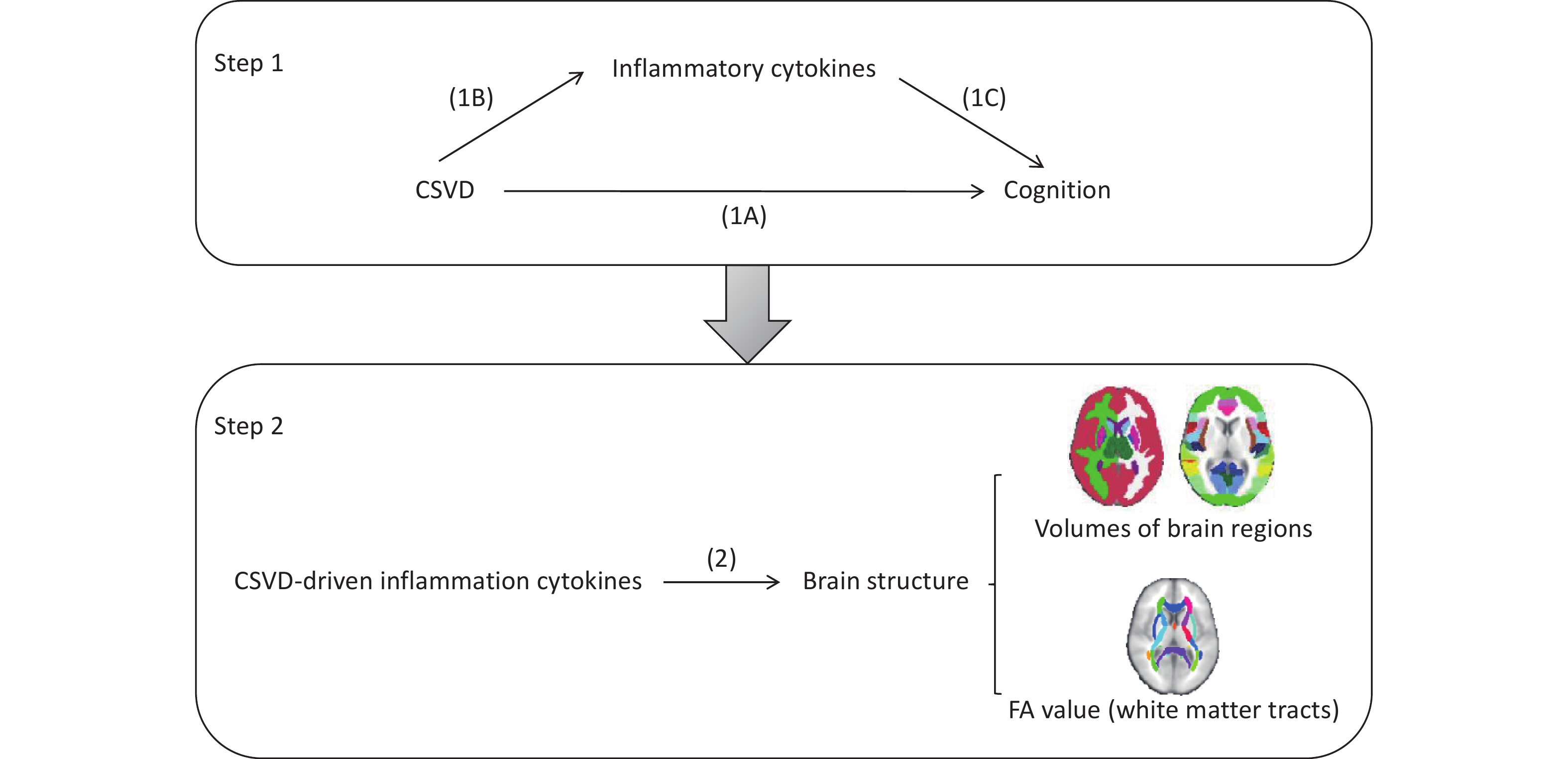
This study consisted of two major components, as outlined in Figure 1. First, we performed a two-step, two-sample MR analysis[18] using publicly available genome-wide association study (GWAS) datasets for CSVD, circulating inflammatory cytokines, and cognitive function. We initially evaluated the impact of CSVD on cognitive function (Figure 1A), and then assessed the potential mediating role of circulating inflammatory cytokines using a two-step MR framework. In the first step, we examined the causal effects of CSVD on circulating inflammatory cytokines (Figure 1B). In the second step, we evaluated the causal effects of CSVD-driven inflammatory cytokines on cognitive function (Figure 1C).
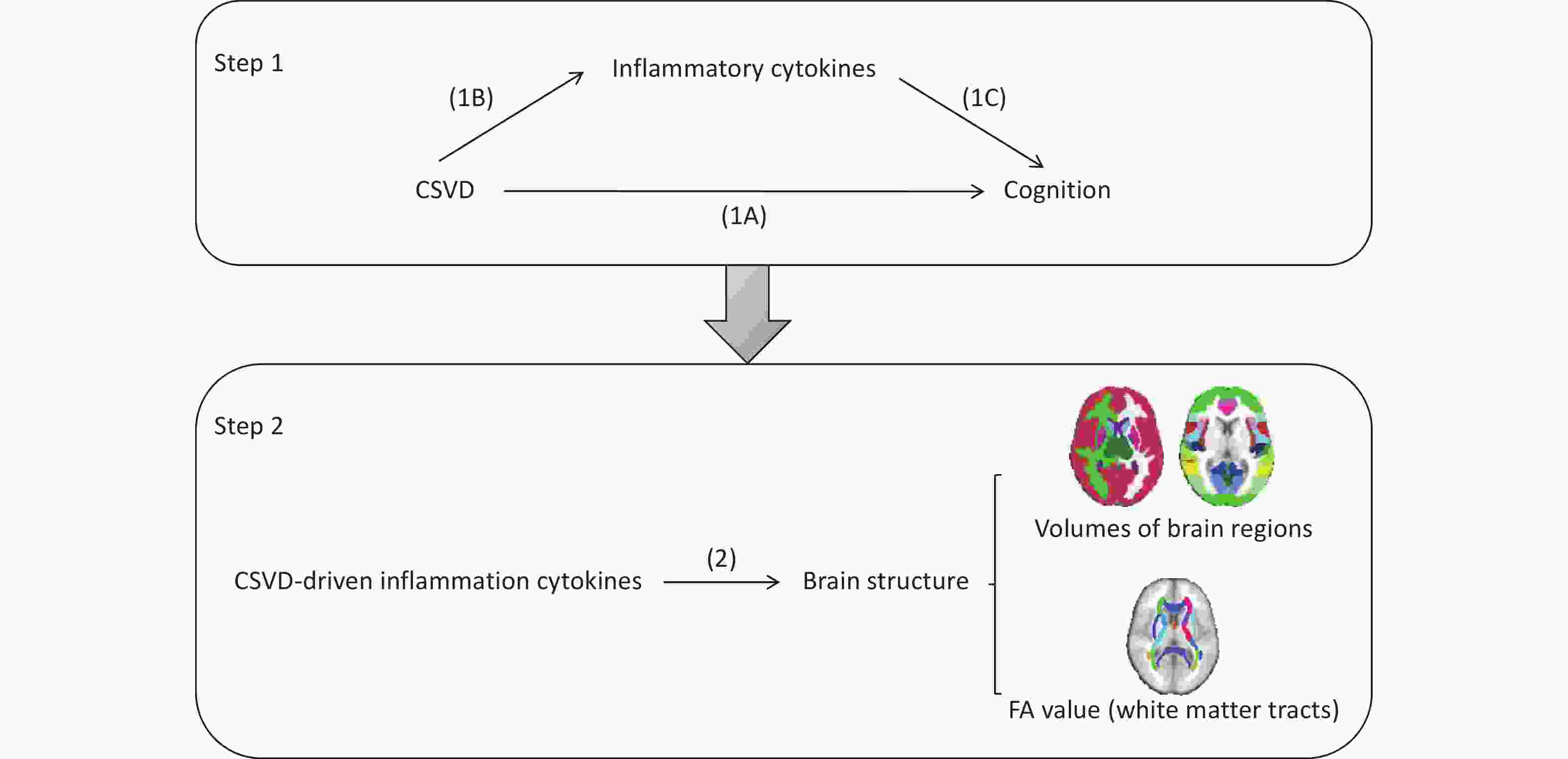
Figure 1. Schematic illustration of the analytical framework used in this study. (1A) Two-sample MR analysis assessing the effect of CSVD on cognition. (1B) Two-sample MR analysis assessing the effect of CSVD on circulating inflammatory cytokines. (1C) Two-sample MR analysis assessing the effect of inflammatory cytokines on cognition. (2) Two-sample MR analysis investigating the effect of CSVD-driven inflammatory cytokines on brain structural changes. Samples included the volumes of 110 brain regions, comprising 96 cortical and 14 subcortical regions based on the Harvard–Oxford atlas. The FA values of 48 white matter tracts were obtained using tract-based spatial statistics (TBSS) method. FA, fractional anisotropy.
Second, to investigate the potential mechanisms through which CSVD-driven inflammatory cytokines influence cognition, we performed an additional analysis to examine the causal effects these inflammatory cytokines on brain structural changes (2).
-
In the first step of the analysis, we used summary statistics from the most recent GWASs of CSVD-related phenotypes as exposure dataset. These included WMH volume (N = 32,114), small vessel ischemic stroke (5,386 cases and 192,662 controls), lacunar stroke (6,030 cases and 248,929 controls), CMBs (3,556 cases and 22,306 controls), small vessel stroke (6,811 cases and 1,234,808 controls), intracerebral hemorrhages (ICH) (655 cases and 455,693 controls), and basal ganglia-PVS (BG-PVS) (9,317 cases and 29,281 controls). Fractional anisotropy (FA), a reliable measure of white matter microstructural integrity, was also included as a CSVD-related phenotype using whole-brain mean FA values (N = 17,663). Higher FA values generally indicate better white matter integrity. Detailed definitions of CSVD neuroimaging biomarkers are provided in the Supplementary materials.
For potential mediators, we used the largest publicly available GWAS meta-analysis of 41 circulating inflammatory cytokines, based on unbiased genome-wide screening, including up to 8,293 participants from three independent population cohorts: the Cardiovascular Risk in Young Finns Study, FINRISK 1997 and FINRISK 2002[19,20]. BMI was not included as a covariate in the original model. Detailed information regarding the 41 circulating inflammatory cytokines is presented in Supplementary Table S1.
For cognitive outcomes, we used general cognitive performance data (N = 257,841), obtained from the IEU GWAS pipeline (https://gwas.mrcieu.ac.uk/datasets). Six cognitive measures from the UK Biobank were used to assess four cognitive domains: processing speed, prospective memory, working memory, and verbal and numerical reasoning[21]. Processing speed was evaluated using the number of correct symbol-digit matches, mean time to correctly identify matches, and response time. Prospective memory was assessed using prospective memory test performance. Working memory was evaluated using numeric memory test, whereas verbal and numerical reasoning was assessed using fluid intelligence tasks completed within a specified time limit. Additional details regarding test administration and scoring procedures are provided in Supplementary Materials.
In the second stage of the analysis, structural brain alterations were characterized using brain regional volumes and white matter tract integrity. Summary statistics for 110 brain regions, including 96 cortical and 14 subcortical regions (excluding the brainstem) based on the Harvard–Oxford atlas (https://fsl. fmrib. ox. ac. uk/fsl/fsl. s/Atlases), were derived from the largest available GWAS conducted in the UK Biobank (UKB) population[22]. FA values from 48 white matter tracts were obtained using diffusion MRI based on tract-based spatial statistics. Detailed information on the 110 brain regions and 48 white matter fibers can be found in Supplementary File 1.
To minimize confounding due to population stratification, all datasets primarily restricted to individuals of European ancestry. The only exception was the CMB dataset, which include a small proportion of participants of other ancestries (97% European, 2% Black, < 1% Malay, < 1% Chinese)[23-25]. Detailed characteristics of all datasets included in the analysis are summarized in Table 1.
Trait Author SNPs (N) Sample size Year CSVD Phenotype WMH volume Smith SM et al. 17,103,079 33,224 2021 Small vessel ischemic stroke Malik R et al. 8,280,845 Cases: 5,386
Controls: 192,6622018 Lacunar stroke Traylor M et al. 6,932,927 Cases: 6,030
Controls: 248,9292021 Cerebral microbleeds Knol MJ et al. / Cases: 3,556
Controls: 22,3062020 Small vessel stroke Mishra A et al. 5,784,789 Cases: 6,811
Controls: 1,234,8082022 Fractional anisotropy Persyn E, et al. / 17,663 2020 Intracerebral hemorrhages Jiang L et al. 11,842,647 Cases: 655
Controls: 455,6932021 Basal ganglia
perivascular spacesDuperron MG et al. 8,783,191 Cases: 9,317
Controls: 29,2812023 Cognition Cognitive performance Lee JJ et al. 10,066,414 257,841 2018 Mean time to correctly identify matches Ben Elsworth et al. 9,851,867 459,523 2018 Number of fluid intelligence questions attempted within time limit Ben Elsworth et al. 9,851,867 149,051 2018 Number of symbol digit matches made correctly Ben Elsworth et al. 9,851,867 113,106 2018 Prospective memory result Ben Elsworth et al. 9,851,867 152,605 2018 Round of numeric memory test Ben Elsworth et al. 9,851,867 48,080 2018 Time to answer Ben Elsworth et al. 9,851,867 152,605 2018 41 inflammatory cytokines Ahola-Olli et al. / 8,293 2017 110 brain volume measurement Smith SM et al. 17,103,079 33,219 2021 48 white matter tracts (FA) Smith SM et al. 17,103,079 31,356 2021 Note. CSVD, cerebral small vessel disease; FA, fractional anisotropy; GWAS, genome-wide association study; SNPs, single nucleotide polymorphisms. Table 1. Data sources used to obtain summary statistics for MR analyses
-
Genetic IVs were selected according to the three core assumptions of MR analysis[26]. SNPs associated with CSVD and inflammatory cytokines at genome-wide significance level (P < 1 × 10−5) were selected as IVs to ensure analytical reliability. To ensure independence among IVs, linkage disequilibrium (LD) clumping was with a clumping distance of 10,000 kb and an LD threshold of r2 < 0.001[27]. Instrument strength was assessed using F-statistics, with F < 10 considered indicative of weak instruments; such SNPs were excluded from further analyses[28]. The F-statistic was calculated as follows:
$$ {R}^{2}=\frac{{\beta }^{2}}{{\beta }^{2}+S{E}^{2}\times N} $$ (1) $$ F=\frac{{R}^{2}\times \left(N-2\right)}{1-{R}^{2}} $$ (2) where R2 represents the proportion of variance in the exposure explained by the SNP, β denotes the magnitude of allele-associated effect, SE estimate the precision of the exposure effect, and N indicates the sample size of the exposure GWAS.
To reduce potential pleiotropy, we searched PhenoScanner V2 for SNPs associated with known confounders of the exposure–outcome relationship (e.g., educational attainment for cognitive outcomes and autoimmune diseases for inflammatory cytokines) or traits directly related to the outcomes (e.g., Alzheimer’s disease or neuropsychiatric disorders associated with altered brain structure)[29]. These SNPs were excluded from the analyses.
-
Inverse-variance-weighted (IVW) analysis was used as the primary MR method. To evaluate the robustness of the findings, four complementary MR approaches were additionally applied: MR-Egger, weighted median, simple mode, and weighted mode. To account for multiple testing, Bonferroni correction was applied according to the number of outcome variables within each category. The corrected significance thresholds were set as follows: analyses involving cognitive outcome (n = 7), P < 0.05/7 = 7.14 × 10-3; inflammatory cytokines (n = 41), P < 1.22 × 10-3; brain volumes (n = 110), P < 4.55 × 10-4; and white matter tract FA values (n = 48), P < 1.04 × 10-3. Given the exploratory nature of this study, associations with P-values between the Bonferroni-corrected threshold and 0.05 were considered nominally significant[24,30].
Heterogeneity was assessed using Cochran’s Qvalues statistics[31], with P < 0.05 indicating significant heterogeneity. Leave-one-out sensitivity analyses were performed to identify influential SNPs and assess the reliance of the causal effect estimate on individual genetic variants[32]. Horizontal pleiotropy was evaluated using both the MR-Egger intercept test and the MR pleiotropy residual sum and outlier (MR-PRESSO)[33]. A significant MR-Egger intercept (P <0.05) was considered indicative of horizontal pleiotropy. MR-PRESSO was further used to identify outlier IVs and to estimate causal effects after excluding these outliers[17,34].
All MR analyses were performed using R software (version 4.3.1) with the “TwoSampleMR” (version 0.5.7)[35] and “MR-PRESSO” (version 1.0) packages [34].
-
To obtain the mediation effect of each circulating inflammatory cytokine, the effect of CSVD phenotype on each circulating inflammatory cytokine was multiplied with an estimate of the effect of each circulating inflammatory cytokine on cognition. The proportion mediated was subsequently calculated by dividing the mediation effect by the total effect of CSVD on cognitive[36].
-
Following quality control procedures, the lists of included SNPs are provided in Supplementary File 2.
-
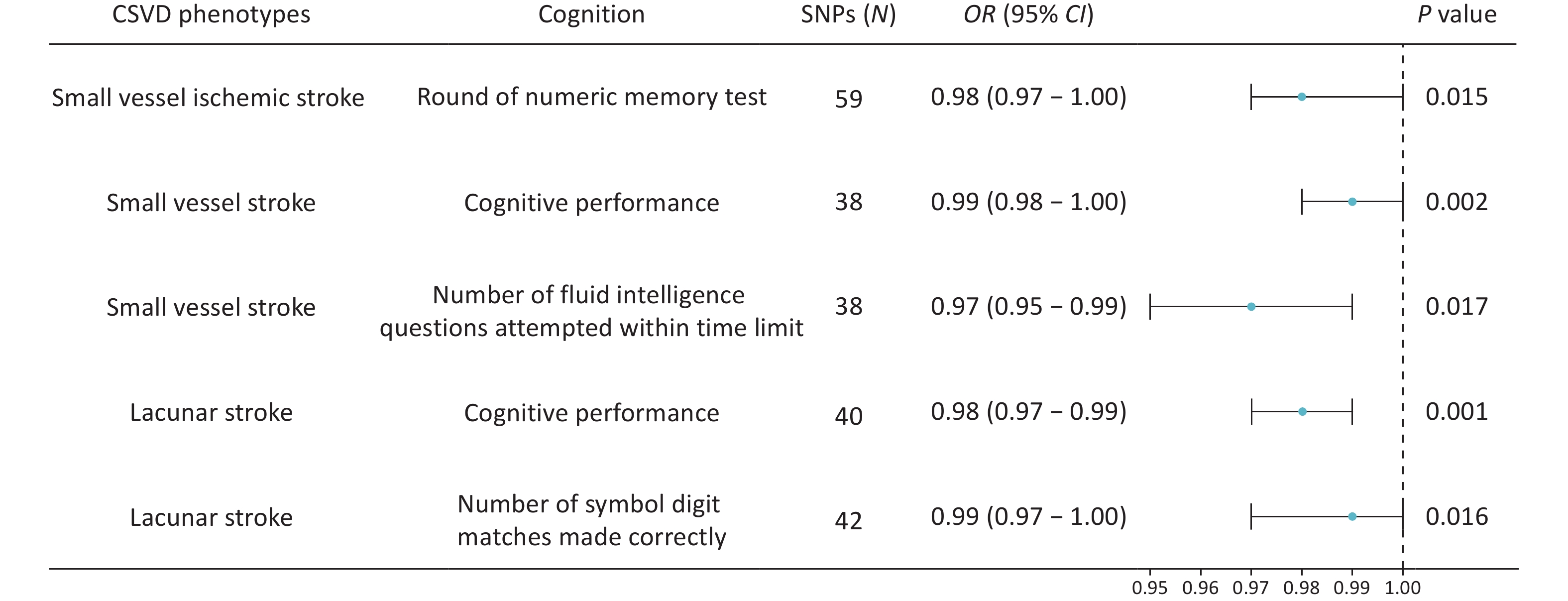
The primary IVW analysis suggested that genetically predicted CSVD was inversely associated with cognitive function. After Bonferroni correction for multiple testing (P < 7.14 × 10-3), only genetically predicted lacunar stroke showed a significant inverse association with cognitive performance (OR = 0.98, P = 0.001, Figure 2).
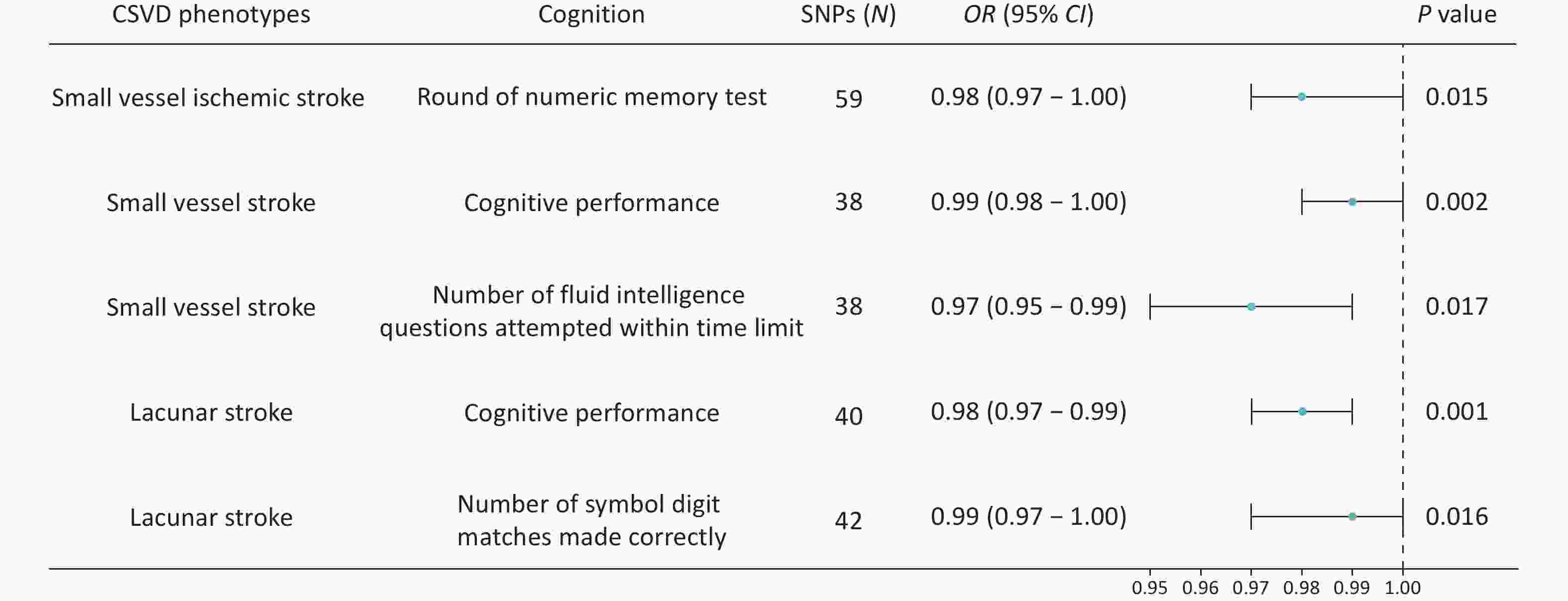
Figure 2. Forest plots showing the causal associations between genetically predicted CSVD and cognition using IVW methods. The SNPs column represents the number of SNPs retained after clumping for independence and data harmonization. CI, confidence interval; OR, odds ratio; SNPs, single nucleotide polymorphisms.
Several associations reached nominal significance. Genetically predicted small-vessel ischemic stroke risk was nominally associated with poorer performance on the numeric memory test (OR = 0.98, P = 0.008; Figure 2). Similarly, small vessel stroke risk showed inverse causal associations with overall cognitive performance (OR = 0.99, P = 0.031; Figure 2) and the number of fluid intelligence questions attempted within the time limit (OR = 0.97, P = 0.017; Figure 2). Genetically predicted lacunar stroke risk was also inversely associated with the number of correct symbol-digit matches (OR = 0.99, P = 0.016; Figure 2).
In addition, genetically predicted ICH (OR = 0.99, P = 0.083) and CMBs (OR = 0.99, P = 0.086) showed trends toward poorer performance on the numeric memory test. Whole-brain FA values also demonstrated an inverse trend with the mean time required to correctly identify matches, as detailed in Supplementary File 3. Results obtained using the MR-Egger, weighted median, simple mode, and weighted mode methods are presented in Supplementary File 3.
Cochran’s Q statistic indicated no evidence of substantial heterogeneity in any analyses. Furthermore, neither the MR-Egger intercept test nor the MR-PRESSO global test detected evidence of horizontal pleiotropy. Detailed results are presented in Supplementary File 3 and 4. Leave-one-out sensitivity analyses are shown in Supplementary Figure S1.
-
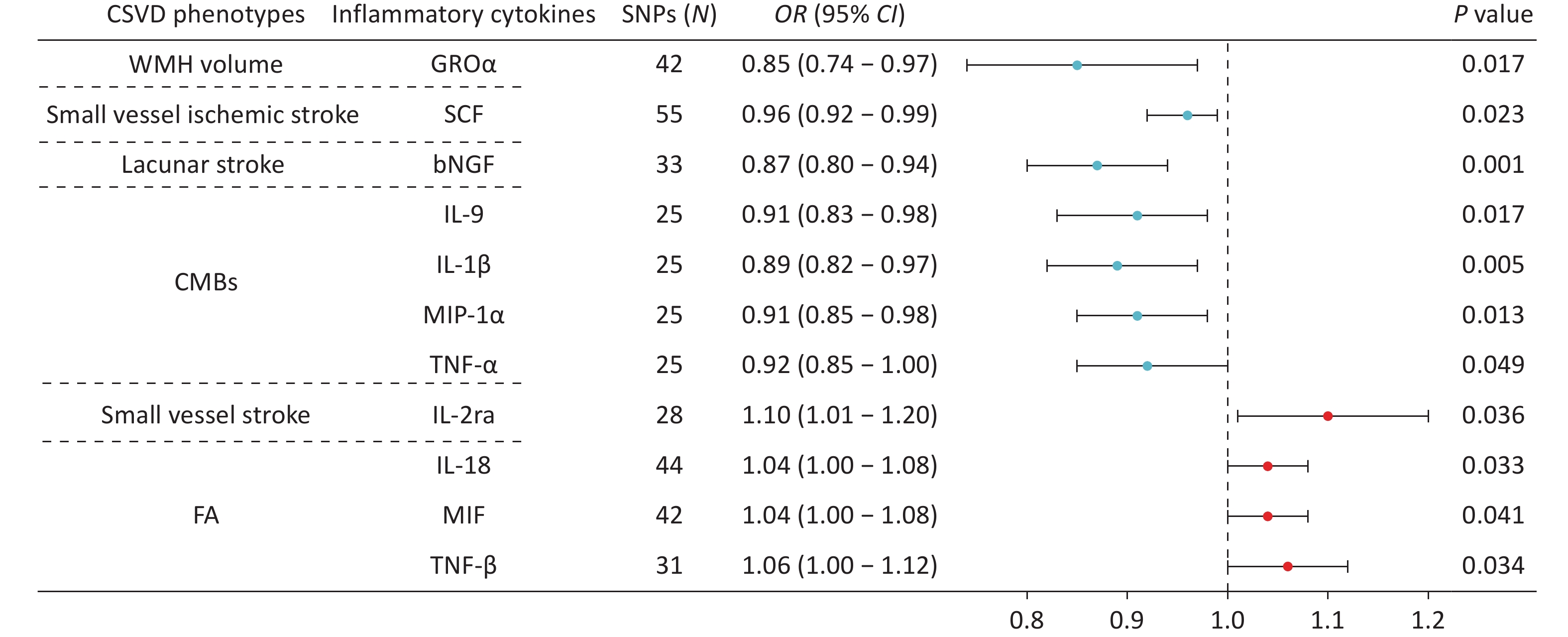
Using IVW analysis, we identified 11 causal relationships between genetically predicted CSVD phenotypes and circulating inflammatory cytokines (Figure 3). After Bonferroni correction (P = 1.22 × 10−3), only the association between genetically predicted lacunar stroke circulating bNGF remained statistically significant (OR = 0.87, P = 0.001).
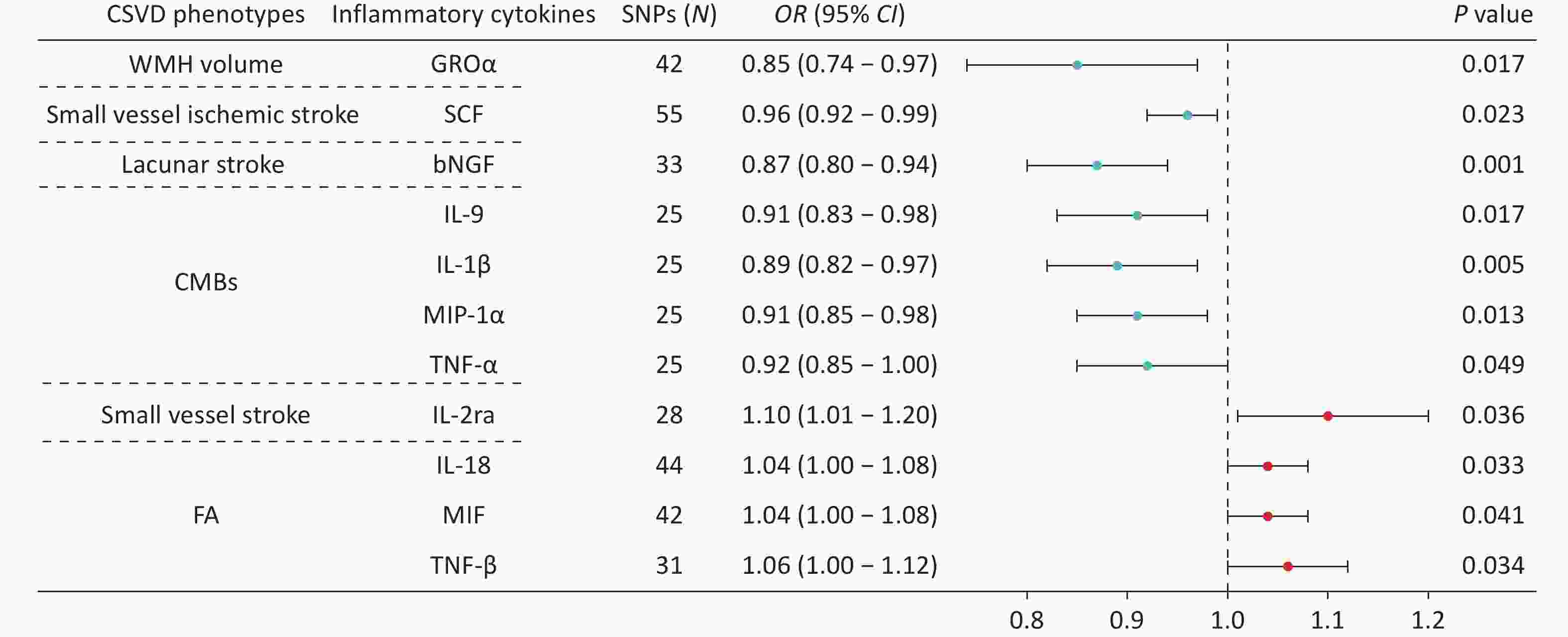
Figure 3. Forest plots showing the causal associations between genetically predicted CSVD and inflammatory cytokines using IVW methods. The SNPs column indicates the number of SNPs retained after clumping for independence and data harmonization. The core biological functions of the cytokines are as follows: GROα (chemotaxis/angiogenesis), SCF (neuroprotection), bNGF (neurotrophin/neuronal survival), IL-9 (T cell growth factor/mast cell growth factor), IL-1β (pro-inflammatory), MIP-1α (inflammatory chemokine), TNF-α (pro-inflammatory/neurotoxicity), IL-2ra (IL-2 receptor subunit/Treg homeostasis/immune tolerance), IL-18 (pro-inflammatory), MIF (pro-inflammatory/tissue repair), TNF-β (cytotoxic/immunomodulatory). CI, confidence interval; CMBs, cerebral microbleeds; CSVD, cerebral small vessel disease; FA, fractional anisotropy; OR, odds ratio; SNPs, single nucleotide polymorphisms; WMH, white matter hyperintensity.
The remaining associations were nominally significant and exploratory in nature. Genetically predicted WMH volume was inversely associated with circulating GROα levels (OR = 0.85, P = 0.017), whereas small vessel ischemic stroke was inversely associated with circulating SCF concentrations (OR = 0.96, P = 0.023). Genetically predicted CMBs were negatively correlated with circulating IL-9 (OR = 0.91, P = 0.017), IL-1β (OR = 0.89, P = 0.005), MIP-1α (OR = 0.91, P = 0.013), and TNF-α (OR = 0.92, P = 0.049). In contrast, genetically predicted small vessel stroke was positively associated with IL-2ra (OR = 1.10, P = 0.036). Genetically predicted FA values were also positively correlated with IL-18 (OR = 1.04, P = 0.033), MIF (OR = 1.04, P = 0.041), and TNF-β (OR = 1.06, P = 0.034). Results from the MR-Egger, weighted median, simple mode, and weighted mode analyses demonstrated the robustness of these findings (Supplementary Figure S2).
No significant heterogeneity was detected in any analysis, and neither the MR-Egger intercept test nor the MR-PRESSO global test indicated horizontal pleiotropy (Supplementary Figure S2 and Supplementary File 4). Leave-one-out sensitivity analyses are shown in Supplementary Figure S3.
-
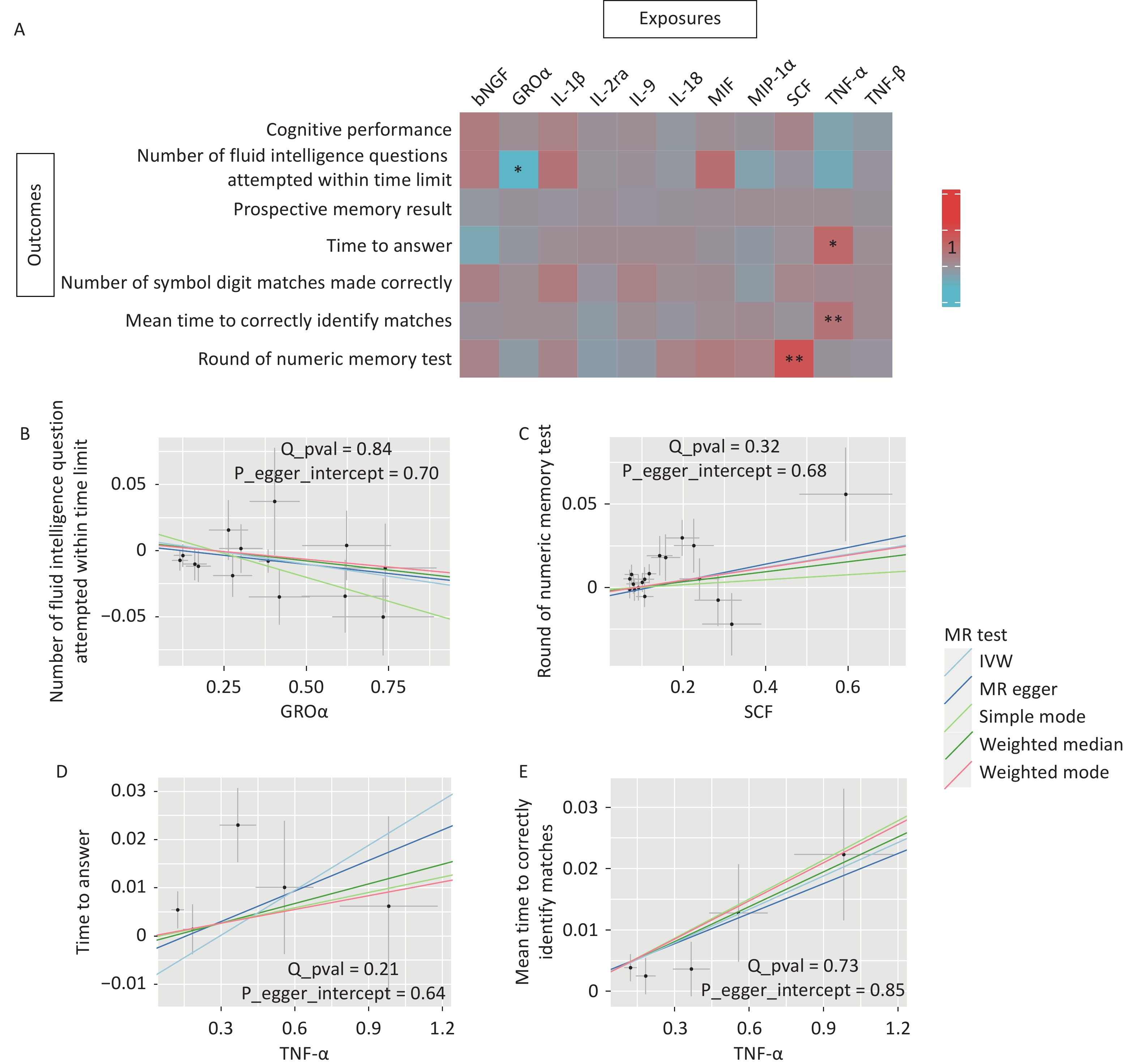
To investigate whether inflammatory cytokines mediate the relationship between CSVD and cognitive function, we used 11 circulating inflammatory cytokines causally associated with CSVD phenotypes as exposures and cognition as outcomes. The primary IVW analysis identified two associations that survived stringent Bonferroni correction (P < 7.14 × 10-3). Genetically predicted higher circulating SCF concentrations were significantly associated with better numeric memory performance (OR = 1.04, P = 0.005; Figure 4). In contrast, genetically predicted higher circulating TNF-α levels were significantly associated with longer mean times to correctly identify matches (OR = 1.02, P = 0.002; Figure 4).
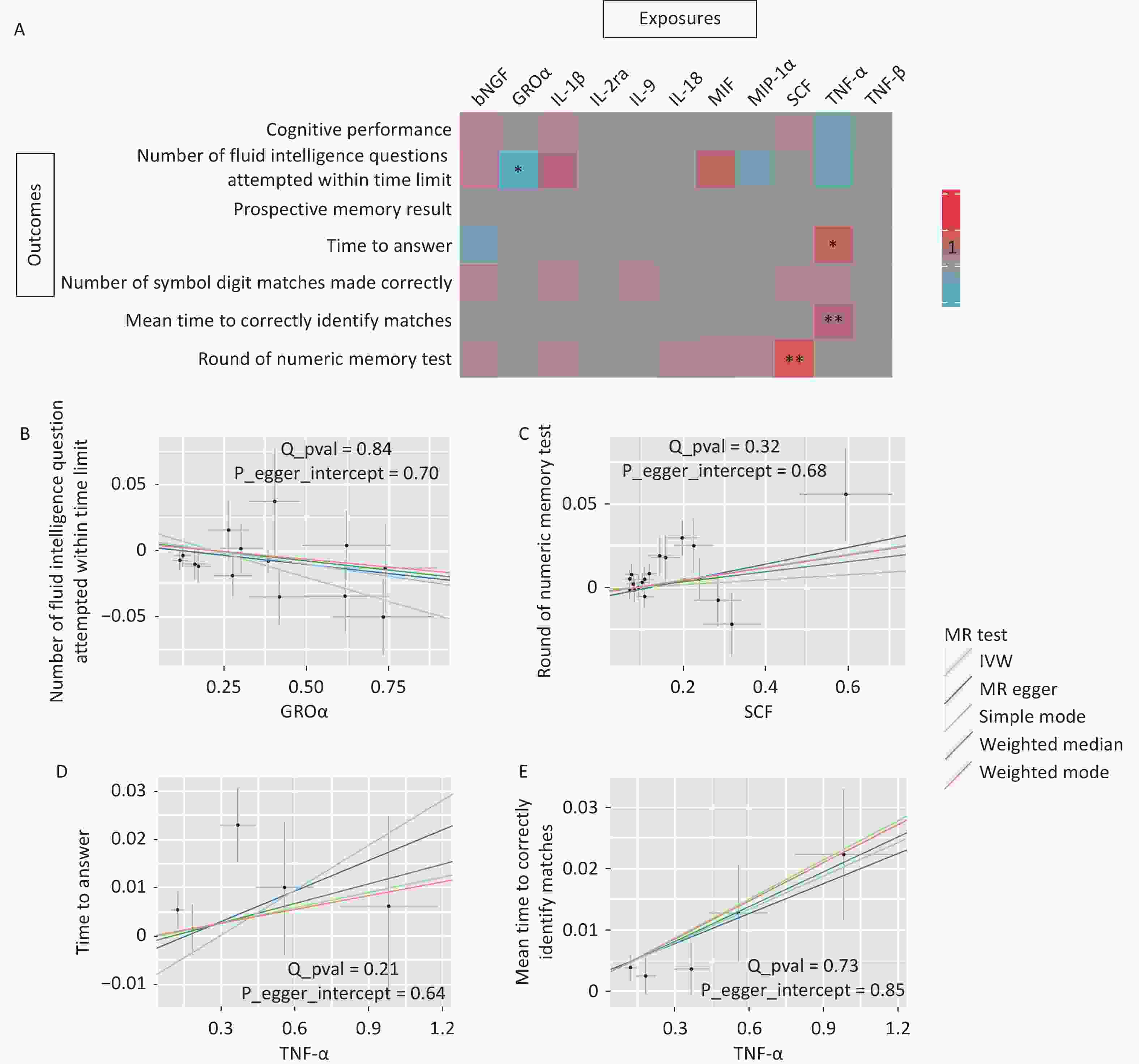
Figure 4. MR associations between CSVD-driven inflammatory cytokines and cognitive function. (A) Heatmap illustrating the causal effects of 11 CSVD-driven inflammatory cytokines on seven cognitive outcomes. Colors indicate the magnitude and direction of OR values for association of inflammatory cytokines and each cognitive test by MR analysis (red indicates OR > 1; blue indicates OR < 1). Asterisks denote statistical significance: *p < 0.05; **p < 7.14 × 10-3 (Bonferroni corrected threshold). (B–E) Scatter plots showing MR analyses of causal associations between circulating GROα concentrations and the number of fluid intelligence questions attempted within time limit, SCF concentrations and round of numeric memory test, TNF-α concentrations and time to answer, and TNF-α concentrations and mean time required to correctly identify matches. The SNP effects are plotted for IVW (light blue line), MR-Egger (dark blue line), simple mode (light green line), weighted median (dark green line), and weighted mode (red line). The slope of each line corresponds to the estimated causal effect.
Two additional associations were nomically significant. Higher genetically predicted GROα concentrations were associated with fewer fluid intelligence questions attempted within time limit (OR = 0.97, P = 0.012, Figure 4). Additionally, higher circulating TNF-α concentrations were nominally associated with longer response time when answering questions (OR = 1.03, P = 0.020, Figure 4).
Cochran’s Q statistics showed no evidence of heterogeneity in any analysis. Likewise, the MR-Egger intercept and MR-PRESSO global tests did not show evidence of horizontal pleiotropy (Figure 4 and Supplementary File 4). Leave-one-out sensitivity analyses are shown in Supplementary Figure S4.
-
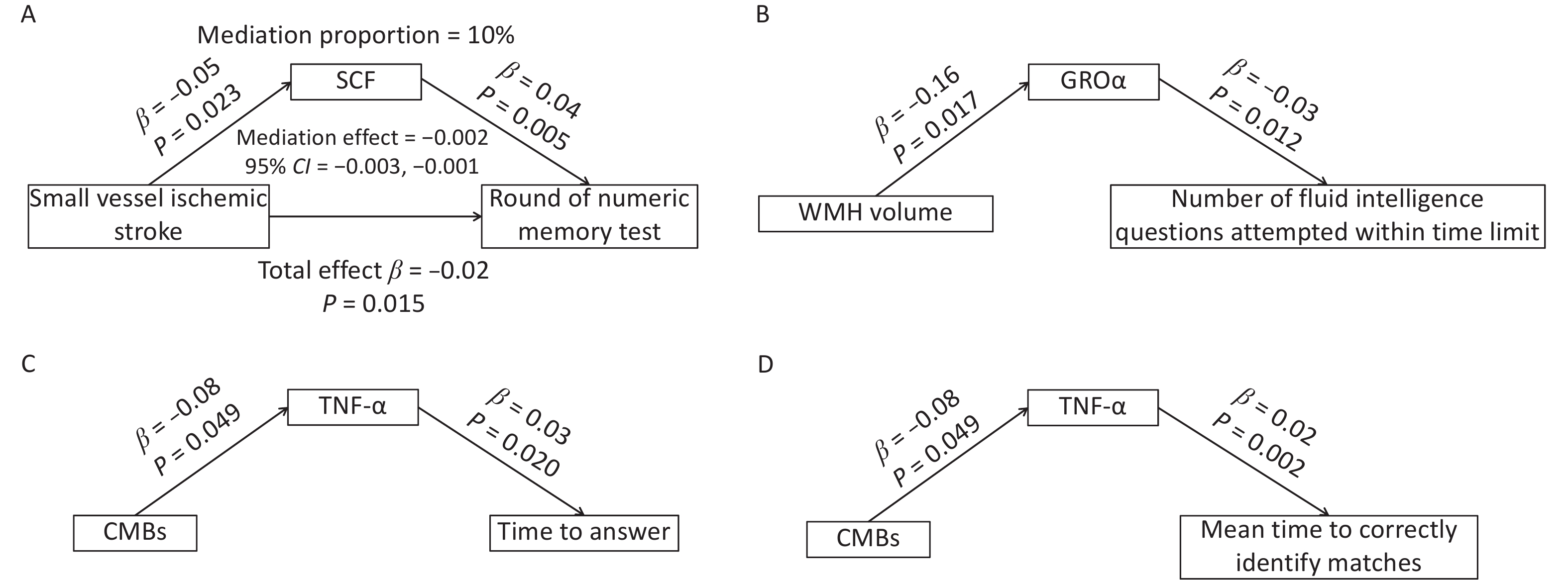
After excluding circulating inflammatory cytokines that were not causally associated with either CSVD or cognitive function, SCF, GROα, and TNF-α were included in the mediation analyses. SCF mediated approximately 10% of the causal effect of small vessel ischemic stroke on numeric memory performance (Figure 5A). Although no direct causal association was identified between WMH volume and fluid intelligence results, circulating GROα appeared to fully mediate component in the relationship between these two variables (Figure 5B). Similarly, circulating TNF-α fully mediated the associations between CMBs and response time (Figure 5C), as well as between CMBs and the mean time required to correctly identify matches (Figure 5D).
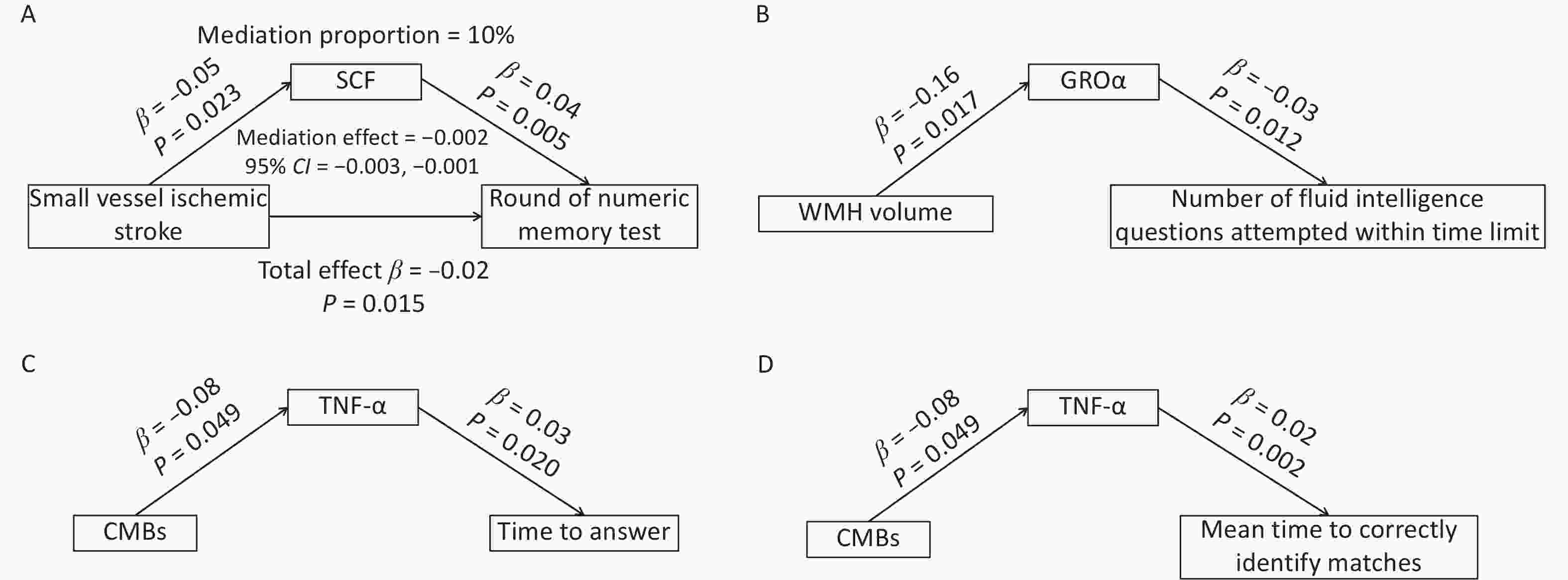
Figure 5. Mediation effects of circulating SCF, GROα, and TNF-α in the association between CSVD and cognition. (A) Circulating SCF partially mediated the association between small-vessel ischemic stroke and round of numeric memory test. (B) Circulating GROα fully mediated the relationship between WMH volume and the number of fluid intelligence questions attempted within time limit. (C) Circulating TNF-α fully mediated the association between CMBs and time to answer questions. (D) Circulating TNF-α fully mediated the association between CMBs and the mean time required to correctly identify matches. CMBs, cerebral microbleeds; WMH, white matter hyperintensity.
-
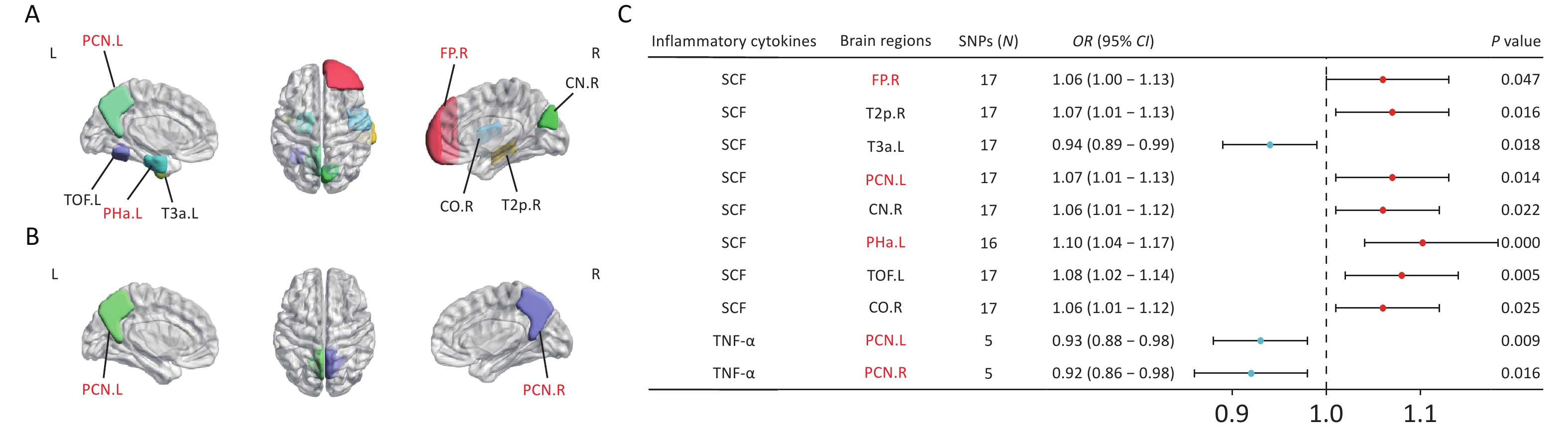
To explore the mechanism through which CSVD-driven inflammatory cytokines influence cognitive function, we investigated the causal effects of SCF, GROα, and TNF-α on brain structural alterations. MR analyses demonstrated that genetically predicted circulating SCF concentrations were nominally associated with the volumes of several brain regions (Figure 6A). Specifically, SCF showed positive associations with the volumes of the right frontal pole (FP.R), right posterior middle temporal gyrus (T2p.R), left precuneus cortex (PCN.L), right cuneal cortex (CN.R), left anterior parahippocampal gyrus (Pha.L), left temporal-occipital fusiform cortex (TOF.L), and right central opercular cortex (CO.R). In contrast, SCF showed an inverse association with left anterior inferior temporal gyrus (T3a.L) (Figure 6C). Circulating TNF-α concentrations were negatively associated with the volumes of the bilateral precuneus cortex (PCN.L and PCN.R) (Figures 6B and 6C). No causal associations were observed between circulating GROα and brain area volumes. After Bonferroni correction (P < 4.55 × 10-4), only the association between circulating SCF and the volume of Pha.L remained statistically significant (OR = 1.1, P = 4.54 × 10-4). Results from the MR-Egger, weighted median, simple mode, and weighted mode analyses are presented in Supplementary Figure S5. Leave-one-out sensitivity analyses are shown in Supplementary Figure S6.
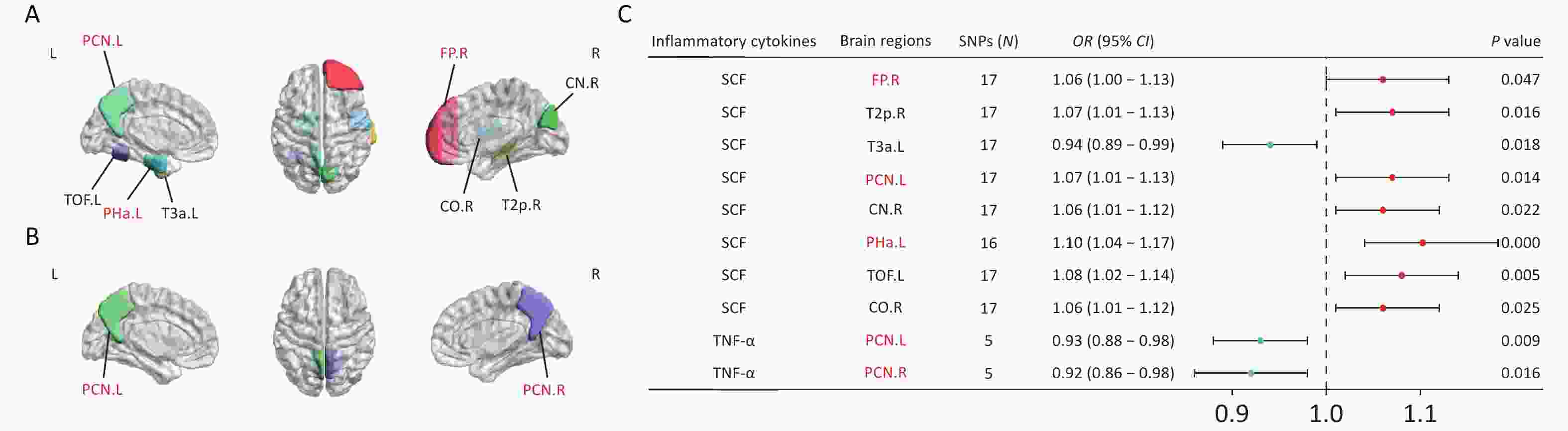
Figure 6. MR estimates of causal associations between mediating inflammatory cytokines and brain regional volumes. (A) Brain regions showing causal associations with SCF. (B) Brain regions with causal associations with TNF-α. (C) Forest plots illustrating the causal associations between genetically predicted inflammatory cytokines and brain regional volumes using the IVW methods. Brain regions highlighted in red represent the default mode network (DMN), which is involved in episodic memory, self-referential processing, and other cognitive functions. Black indicates non-DMN regions (e.g., T2p.R, T3a.L, and CN.R), which are involved in visuospatial function, language function, and motor control. BrainNet Viewer toolbox (https://helab.bnu.edu.cn/brainnet-viewer/) was used to display brain regions using automated anatomical labeling based on Harvard-Oxford atlas.
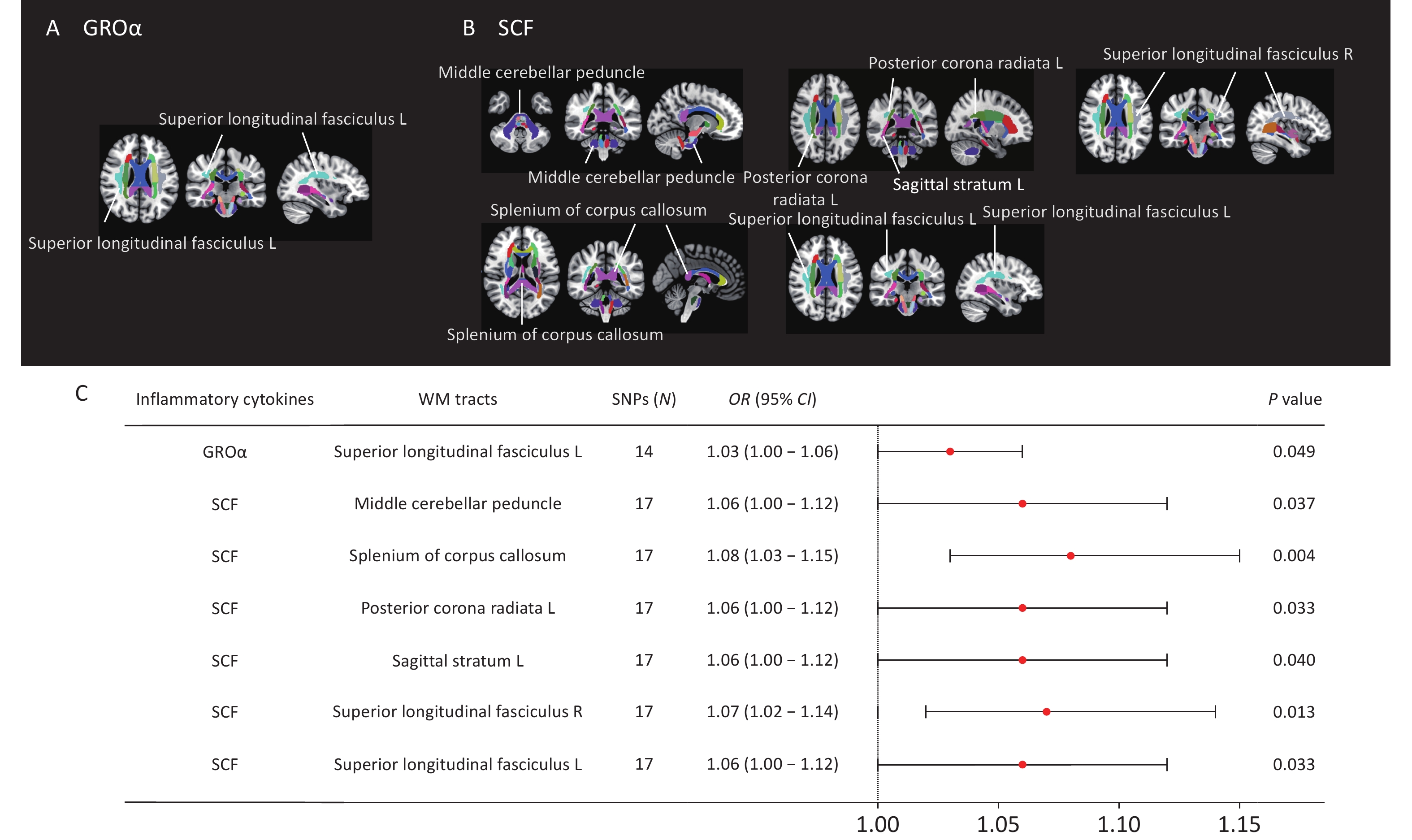
We identified eight nominally significant causal associations between circulating inflammatory cytokines and white matter tract integrity (Figure 7). Circulating GROα levels were positively correlated with FA values in the left superior longitudinal fasciculus (OR = 1.03, P = 0.049). Circulating SCF concentrations demonstrated positive causal relationship with FA values in the middle cerebellar peduncle (OR = 1.06, P = 0.037), splenium of corpus callosum (OR = 1.08, P = 0.004), left posterior corona radiata (OR = 1.06, P = 0.033), left sagittal stratum (OR = 1.06, P = 0.040), right external capsule (OR = 1.07, P = 0.049), and bilateral superior longitudinal fasciculus (left: OR = 1.06, P = 0.033; right: OR = 1.07, P = 0.013). However, none of these associations survived Bonferroni correction for multiple testing (P < 1.04 × 10-3). No causal relationship was identified between circulating TNF-α and white matter integrity. Results from the MR-Egger, weighted median, simple mode, and weighted mode analyses are shown in Supplementary Figure S7. Leave-one-out sensitivity analyses are shown in Supplementary Figure S8.
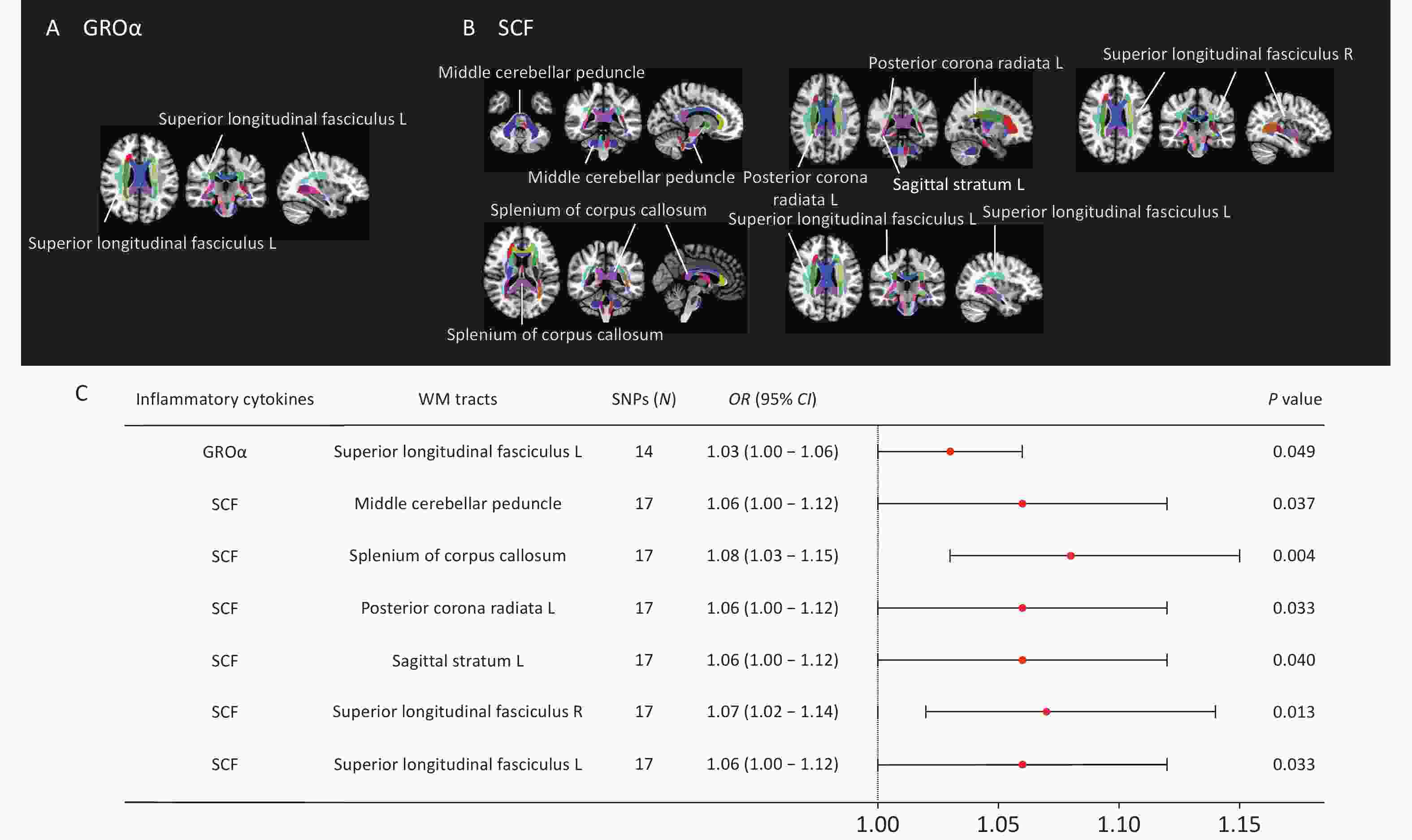
Figure 7. MR estimates of causal associations between mediating inflammatory cytokines and FA values of white matter tracts. (A) Distribution map of white matter fiber tracts causally associated with GROα. (B) Distribution map of white matter fiber tracts with causal relationship with SCF. (C) Forest plots illustrating the causal associations between genetically predicted inflammatory cytokines and FA values of white matter fiber tracts by using the IVW methods. White matter tract visualization was performed using the JHU-ICBMlabels-1 mm white matter atlas as a template and the FSLeyes package (https://fsl.fmrib.ox.ac.uk/fsl/fslwiki/FSLeyes).
No significant heterogeneity was observed in any analysis. The MR-Egger intercept test indicated potential horizontal pleiotropy in the association between circulating SCF and FA values in the right external capsule (P = 0.044), whereas the MR-PRESSO global test was not significant (P = 0.081). No evidence of horizontal pleiotropy was observed in the remaining analyses. Detailed results are provided in Supplementary Figure S5 and S7 and Supplementary File 4.
-
Using Mendelian randomization analyses based on GWAS datasets, we identified evidence supporting a causal effect relationship between CSVD and cognitive impairment and further demonstrated that several circulating inflammatory cytokines play important roles in this pathway. Specifically, we found that CSVD had a causal effect on 11 circulating inflammatory cytokines. Among these cytokines, GROα was associated with lower verbal and numerical reasoning, SCF was associated with better working memory, and TNF-α was associated with impaired processing speed. Importantly, SCF, the most prominent mediator identified in this study, showed causal associations with the volumes of multiple brain regions within the default mode network, including the PHa.L, FP.R, T2p.R, T3a.L, and PCN. L, as well as the integrity of seven white matter tracts. These findings partially validate previous observations while also providing novel insights into the complex role of cytokines in CSVD-related CI and highlighting potential circulating biomarkers and therapeutic targets.
As expected, our findings support an association between CSVD and cognitive dysfunction, consistent with previous observational studies. Cognitive dysfunction associated with CSVD is typically characterized by deficits in attention, processing speed, and executive function, whereas memory impairment is relatively mild[37,38]. From a genetic perspective, we observed that small-vessel ischemic stroke significantly increased the risk of working memory impairment, whereas CMBs and ICH showed trends towards poorer working memory performance. Genetically predicted small vessel stroke was inversely associated with overall cognitive function and verbal and numerical reasoning, whereas lacunar stroke was linked to impaired processing speed and increased risk of global CI. Whole-brain white matter integrity also showed a trend toward association with processing speed.
These findings suggest that distinct CSVD phenotypes may differentially affect specific cognitive domains, an issue that has not been extensively explored in prior studies. Although previous evidence has demonstrated WMHs as the primary pathological basis underlying cerebrovascular impairment in older adults, we did not observe a direct causal relationship between WMH volume and cognition. One possible explanation is that executive function, which is a primary characteristic of patients with WMHs, was not assessed in this study. By leveraging genetic variation and excluding environmental influences, our study provides additional evidence supporting the causal relationship between CSVD and CI.
Cytokines are key regulators of inflammatory responses and are involved in protecting brain tissue from damage and promoting tissue repair. Therefore, identifying circulating biomarkers associated with CSVD progression has substantial clinical relevance. During CSVD development, acute inflammatory stimuli within the brain trigger inflammatory cytokine productions. Previous studies have demonstrated that brain inflammatory responses promote the accumulation of circulating neutrophils and a cytokine storm-like syndrome, resulting in profound pathological effects on the peripheral immune system[39]. The immune-inflammatory responses in CSVD is highly complex, involving multiple interconnected pathways.
In this study, we identified causal relationships between CSVD phenotypes and 11 circulating inflammatory cytokines. Specifically, CSVD phenotypes were negatively correlated with circulating GROα, bNGF, IL-1β, IL-9, MIP-1α, TNF-α, and SCF concentrations, whereas small vessel stroke was positively correlated with IL-2ra. FA values, reflecting white matter integrity, were positively associated with IL-18, MIF, and TNF-β. These findings are generally consistent with previous studies[40-42]. For example, Schmitz et al. reported altered serum IL-1β and TNF-α levels in patients with vascular dementia[11]. Similarly, inflammatory profiles centered on IL-18 have been associated with increased risk of CSVD and CI[41,43], whereas plasma MIF has been proposed as potential biomarkers for CSVD-related CI[44,45]. By using MR analyses, our study strengthens the evidence supporting causal relationships between CSVD and circulating inflammatory cytokines. More importantly, three cytokines (SCF, GROα, TNF-α) emerged as potentially important mediators linking the development of CI in CSVD, thereby highlighting their possible clinical utility as biomarkers.
Inflammation is involved in cognitive decline and the neurodegeneration of brain[46]. Focusing specifically on inflammatory cytokines influenced by CSVD, we investigated whether these cytokines exerted causal effects on cognition. We found that higher circulating SCF concentrations were associated with better working memory performance. Further MR analyses using brain regional volumes and white matter integrity (FA values) as outcomes supported the protective effect of circulating SCF on cognitive function. Specifically, circulating SCF showed positive associations with the volumes of most brain regions within the default mode network and with the integrity of multiple while matter fiber bundles implicated in cognition.
SCF is an essential cytokine regulating hematopoietic stem cell survival and differentiation, and has been shown to promote proliferation-induced angiogenesis and neuroprotection within the central nervous system[47,48]. Its receptor, c-Kit (CD117), is expressed in endothelial cells[49], and SCF transcripts are abundant in the adult thalamus, cerebral cortex, and white matter[50]. Previous studies have shown that SCF treatment enhances remyelination of the ipsilateral outer capsule and striatum[51]. Experimental studies have further demonstrated that systemic SCF administration during the acute phase of stroke reduces infarct size and improves neurological outcomes[52].
In this study, reduced circulating SCF levels in individuals with small-vessel ischemic stroke were associated with impaired working memory, suggesting potential clinical benefit of peripheral SCF administration in treating CSVD-associated CI. This interpretation is supported by experimental evidence from cerebral autosomal dominant arteriopathy with subcortical infarct and leukoencephalopathy (CADASIL) mouse model, in which combined SCF and granulocyte colony-stimulating factor treatment protected against ischemic neuronal damage caused by capillary thrombosis[53]. In another study, SCF and granulocyte colony-stimulating factor improved spatial learning and memory impairment by increasing cerebral vascular endothelial growth factor levels, restoring cerebral blood vessels, and enhancing neuroregeneration and synapse formation[54]. Our findings extend these observations by providing genetic evidence that reduced SCF levels may contribute to cortical atrophy, white matter damage, and CI in humans with CSVD.
Notably, in the analysis examining circulating SCF and FA values in the right external capsule, the MR-Egger intercept test indicated significant horizontal pleiotropy, whereas the MR-PRESSO global test did not. This discrepancy likely reflects methodological differences between the two approaches. The MR-Egger intercept is more sensitive to widespread directional pleiotropy as it evaluates the intercept of a regression between the instrument-outcome associations and its exposures[33]. In contrast, the MR-PRESSO global test primarily identifies pleiotropy by detecting “outliers” within the distribution of genetic variant effects, making it more sensitive at detecting strongly pleiotropic variants[34]. Thus, our findings suggest the presence of a relatively uniform and widespread pleiotropic pattern with small effect sizes, rather than strong effects driven by isolated SNPs. Because the observed P-values were borderline and MR-PRESSO did not identify influential outliers, horizontal pleiotropy is likely weak and unlikely to be dominated by a single variant with strong pleiotropic effects. Nevertheless, this finding should be interpreted cautiously and validated in larger and more diverse cohorts.
GROα, a member of the chemokine family, has been implicated in Alzheimer's disease and other neuroinflammatory disorders[46]. Previous case-control study has shown increased GROα expressions in the brains of patients with Alzheimer's disease after adjustment for vascular comorbidities[55]. Experimental evidence has found that GROα promotes tau hypermethylation in primary cortical neurons[56]. These findings support our observation that GROα is implicated the pathological progression of CI.
Beyond neurodegeneration, GROα is participates in multiple inflammatory processes, including neutrophil recruitment, angiogenesis, and wound healing[57]. We observed negative correlation between circulating GROα concentrations and WMH volume, indicating that advanced white matter injury may significantly decrease peripheral GROα levels. On one hand, GROα-mediated angiogenesis and neutrophil recruitment are essential for tissue repair following injury. However, sustained or excessive activation of these processes can contribute to blood-brain barrier disruption, neuroinflammation, and white matter damage[58]. Therefore, this decrease may reflect either a compensatory response aimed at limiting excessive inflammation and vascular injury or an exhaustion of vascular repair capacity. As CSVD progresses, persistent ischemic injury and endothelial dysfunction can deplete the reservoir of repair mediators, leading to insufficient angiogenesis and impaired wound healing, and accelerating white matter lesions and cognitive decline. However, because our analyses were based on cross-sectional data, the temporal dynamics underlying these changes remain unclear. Future longitudinal and functional studies are warranted to clarify these associations and to determine whether modulation of GROα may have therapeutic potential in CSVD.
TNF-α is a multifunctional cytokine involved in neurodegenerative diseases[59]. Through interactions with TNFR1 and TNFR2, TNF-α regulates cytokine production, cell survival, differentiation, and downstream signaling cascades[60]. TNFR1 signaling is generally associated with neuroinflammation and neuronal injury, whereas TNFR2 may promote neuroprotection and cell survival[60,61]. Zhang et al. found that impaired microglial function and TNF-α secretion in the medial prefrontal cortex during adolescence was an important cause of cognitive flexibility impairment[62]. Ortí-Casañ et al. demonstrated that treatment with anti-TNFR1 antibodies reduce neuronal loss and memory deficits in a neurodegenerative mouse model[63].
Consistent with these observations, our study demonstrated a causal association between elevated circulating TNF-α concentrations and impaired processing speed. Furthermore, TNF-α was negatively associated with bilateral precuneus cortex volume. The precuneus is involved in advanced cognitive function involving situational memory, visual space, and self-related information processing[64]. Consequently, TNF-α may represent a potential target for immunomodulatory interventions in CSVD-related cognitive decline.
This study has some limitations. First, given the large number of exposures evaluated and to retain sufficient statistical power[16,65], we primarily relied on a genome-wide strategy without strictly restricting selection to biologically curated genomic regions. Although this approach can provide supportive evidence, future studies using larger GWAS datasets should employ more biologically informed strategies to further validate these findings[65]. Second, only a subset of associations survived stringent Bonferroni correction; therefore nominally significant associations should be considered exploratory and interpreted cautiously[30]. Third, peripheral inflammatory markers may not fully reflect inflammation within the central nervous system. Future studies incorporating in vivo measurement and distribution analysis of neuroinflammation are needed to better characterize the inflammatory mechanisms underlying CSVD-related cognitive impairment.
Fourth, although most analyses showed no evidence of horizontal pleiotropy according to the MR-Egger intercept and MR-PRESSO global tests, pleiotropic effects cannot be entirely excluded[66]. Additionally, the biological interactions among cytokines are highly complex, and the observed associations may reflect indirect or network-mediated effects rather than direct causal relationships. Future studies using multivariate MR (MVMR) approach may help disentangle these interactions. However, because the number of genetic instruments used for each cytokine was limited, MVMR was not feasible in the current study. Larger and more diverse GWAS meta-analyses and integrative multi-omics approaches may identify more specific genetic instruments for inflammatory cytokines and enhance statistical power.
Fifth, although the CMB cohort predominantly included individuals of European ancestry, a small proportion of participants were from other ancestral backgrounds. Because individual-level genetic data were unavailable, we could not perform stratified analyses or exclude these participants, a challenge common to many previous studies[23,67,68]. However, given the relatively small proportion of non-European participants, the overall impact on our overall findings is limited. Future research with diverse multi-ethnic cohorts is essential to validate the generalizability of our findings.
Finally, because this study relied on summary-level GWAS data, we were unable to adjust for individual-level covariates such as sex, age, BMI, medication use, and lifestyle factors, all of which may influence or modify the observed genetic effects. Future studies integrating individual-level datasets and prospective cohort designs are needed to enable more nuanced causal inferences and external validation.
Using a Mendelian randomization framework, we identified evidence supporting causal relationship between CSVD, circulating inflammatory cytokines, and cognitive impairment. Among CSVD-driven inflammatory cytokines, SCF, GROα, and TNF-α appeared to play key roles in the pathway linking CSVD to cognitive impairment. In particular, circulating SCF may influence cognition through effects on brain volume and white matter integrity. These findings suggest that targeting circulating SCF may be a novel therapeutic strategy for CSVD-related CI and provide important insights into potential biomarkers for early disease management.
HTML
Study Design
Data Sources
Selection of Genetic Instruments
Statistical Analyses
Mediation Analysis
Inverse Association between Genetically Predicted CSVD and Cognition
Association between Genetically Predicted CSVD and Inflammatory Cytokines
Association between CSVD-driven Inflammatory Cytokines and Cognitive Function
Mediation Effects of Circulating Inflammatory Cytokines in the Association between CSVD and Cognition
Association of Circulating SCF, GROα, and TNF-α with Brain Structural Changes
Competing Interests All authors declare no competing interests.
Ethics The publicly available datasets used in this study were obtained from studies that had already received ethical approval. Therefore, no additional ethical review was required for the present analysis.
Authors’ Contributions Mengxue Wang: conceptualization, data curation, formal analysis, methodology, visualization, and drafting of the original manuscript. Zan Wang: conceptualization, investigation, validation, manuscript writing, review, and editing, and funding acquisition. Jingting Kong: resources, formal analysis, methodology, validation. Lv Zhou: methodology and software. Xuezi Zhang: data curation. Yanjuan Wang: manuscript writing, review, and editing. Qingguo Ren: conceptualization, project administration, supervision, funding acquisition.
Data Availability All data generated or analyzed during this study are included in this published article and its supplementary files.
&These authors contributed equally to this work.

 Quick Links
Quick Links
 DownLoad:
DownLoad: