

 下载:
下载:

-
China continues to suffer from one of the most serious rabies epidemics worldwide, even though the reported number of human death cases in 2017 (501) is the lowest in the past 10 years. This low number is due to increased public health awareness and the annual administration of 12-15 million doses of human vaccines[1, 2]. Nevertheless, the rabies epidemic continues to expand geographically over the entire country due to the lack of vaccination strategies for host animals[1]. Although approximately 99% of human rabies cases in China result from dog bites, more than 100 wildlife-transmitted human cases have been reported since the 1990s; among these cases, at least 87 cases originated from Chinese ferret badgers (Melogale moschata, FBs)[3]. Currently, FB-transmitted human rabies is reported only in the mainland of China. Indeed, in accordance with our continuous studies that started in 2008, epidemic rabies in FBs has become an independent cycle in the provinces of Jiangxi, Zhejiang, and Anhui[4, 5]. Although rabies in Taiwanese (in China) FBs has been circulating since 2012 and has started to cross to local dogs and house shrews (Suncus murinus), all FB rabies virus (RABV) isolates belong to China Ⅱ lineage. In addition, no FB-associated human case has been reported in Taiwan island (in China) possibly because of the elimination of dog-transmitted rabies and increased public awareness[6].
In China, RABV isolates can be divided into six genetic lineages (China Ⅰ-Ⅵ)[7]. Among these lineages, China Ⅰ has become the main epidemic group hosted and transmitted by dogs. This lineage has replaced the dog- and wildlife-associated China Ⅱ lineage in the past decades[7]. All RABV isolates from FBs belonged to China Ⅱ lineage prior to 2014, with the exception two isolates in 2009 and 2010 (China Ⅰ)[8]. To characterize the new characteristics of FB rabies epidemic in China, we have isolated RABVs from FBs since 2014. Many of these RABVs were found to be under the dog-associated China Ⅰ group, and this finding suggests the frequent crossover among the viruses from FBs and dogs.
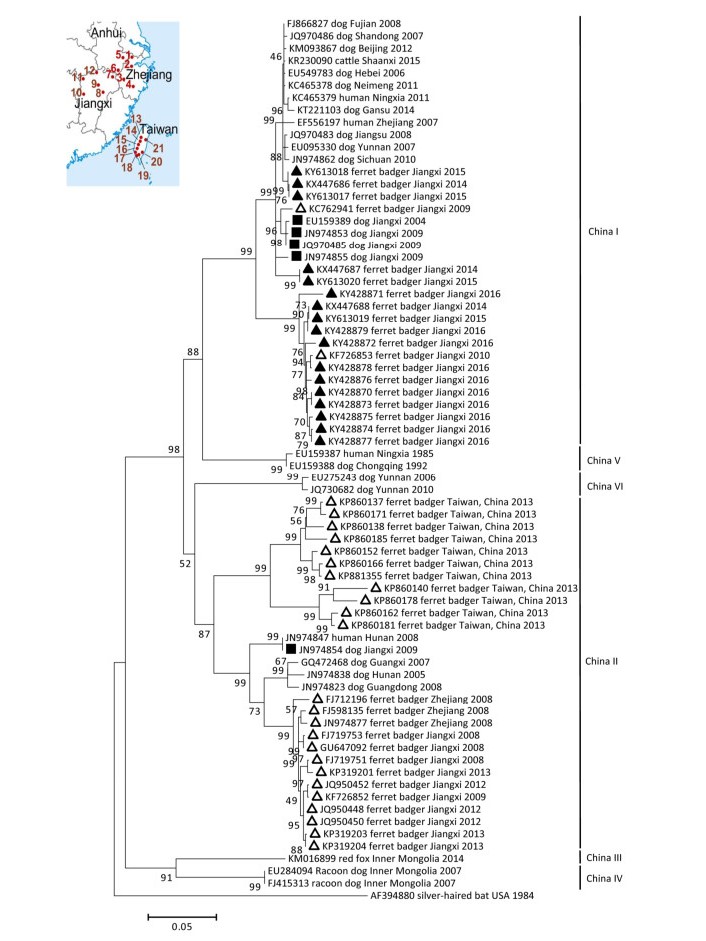
During 2014-2016, 1, 407 brain samples (428, 441, and 538 in 2014, 2015, and 2016, respectively) from dead FBs of unknown causes were collected by local hunters in Wuyuan (263 samples), Fuzhou (207 samples), Xinjian (194 samples), Yichun (188 samples), Xiushui (332 samples), and Hukou (223 samples) Counties of Jiangxi province. Of these, 43 samples (11, 13, and 19 in 2014, 2015, and 2016, respectively) were rabies-positive by laboratory diagnosis according to the results of the direct immunofluorescence assay[1]. Viral RNA was extracted from the positive samples and analyzed through nested reverse transcription polymerase chain reaction as described previously[1]. The complete viral nucleoprotein (N) genes were sequenced. Of these sequences, 25 duplicate N sequences were excluded, and the remaining 17 were submitted to GenBank, with accession numbers shown in Figure 1 (▲). The percentage identities and similarity scores of the N sequences were calculated by using DNAstar Lasergene software (DNASTAR, Inc., Madison, USA). A maximum likelihood phylogenetic tree was constructed using MEGA 7.0.14 as described previously[1]. Sixty representative sequences of China lineages of RABV, including previous rabies isolates from dogs (■) in Jiangxi province and FBs (△) nationwide (Figure 1), were retrieved from GenBank with the accession numbers shown in Figure 1.
Figure 1. Maximum likelihood phylogenetic tree based on the complete nucleoprotein gene of rabies viruses isolated from China, rooted with a RABV virus isolated from a silver-haired bat in USA.

Figure 1 illustrates that since the first isolation of RABV strain ZJ-LA (Genbank #FJ598135) from an FB in 2008, almost all FB isolates until 2013 belong to the China Ⅱ lineage apart from strains JX09-17 (Genbank #KC762941) and JX10-37 (Genbank #KF726853) in the China Ⅰ lineage[5, 8]. However, since 2014, all RABVs isolated from the FBs in Jiangxi have been classified under the China Ⅰ lineage. The current main lineage (including isolates from dogs in Jiangxi) originated from dogs nationwide. This lineage is present in at least two sub-lineages with 94%-98% similarity in N genes between each other. The similarity of FB-RABVs in N genes between China Ⅰ and Ⅱ is approximately 88%. The FB-associated China Ⅰ lineage is rapidly displacing the FB-associated China Ⅱ lineage, gaining dominance of the FB-transmitted rabies epidemic. Moreover, the FB-associated China Ⅰ sub-lineage is distinguished from others in dog-hosted China Ⅰ lineage and possibly becoming the main independent FB-associated clade in the phylogenetic tree. Although only limited geographic origin of FB RABV sequences and numbers of dog RABV sequences from the Jiangxi province were analyzed in the study, the findings suggests that the genetic diversity of RABVs in FBs and the significant differences in the host range of the lineages result from the coexisting dog rabies in China.
In accordance with previous studies, FB RABVs have formed independent infection cycles that probably originated from trans-species infection of dog rabies during long-term epidemics[4, 5]. In China, dog rabies is not thoroughly controlled on the records due to the absence of widespread vaccination programs[7]. The emergence of the China Ⅰ lineage in FB-transmitted rabies and the gradual displacement of the China Ⅱ lineage are also possibly associated with the fundamental role of dog rabies. The reason is that dog-associated China Ⅰ lineage has become the main epidemic group, replacing the China Ⅱ lineage hosted and transmitted by dog and wildlife in the past decades[7]. Furthermore, the ongoing independent rabies epizootic in FBs suggests that sustained rabies transmission chains among FBs seem to occur between 3 and 5 years or longer following dog rabies transmission, such as the shift of dog-hosted RABV into wildlife in the USA[9]. Therefore, the threat of FB transmission to humans has become complicated and serious with the influence of dog rabies in China.
Rabies is a vaccine-preventable zoonosis that has been controlled and even eliminated in many countries by mass vaccination of reservoir animals. For the control and prevent FB-associated rabies from the origin, performing mass immunization of dog against rabies should be prioritized. In China, the product of oral vaccines for wildlife remains insufficient due to certain technical and financial reasons. As FBs frequent rice fields and vegetable gardens as feeding sites, efficient and effective vaccinations can be performed in these places[10].
Maintaining rabies control in animals is a long-term national project. Thus, China should acknowledge the facts about rabies transmission and contribute to the eliminate human rabies cases worldwide by 2030.
The authors declare that they have no competing interests.
Conceived and designed the experiments: HU Rong Liang and LIU Ye. Performed the experiments: MIAO Fa Ming, CHEN Teng, ZHANG Shou Feng, ZHANG Fei, and LI Nan. Contributed reagents/ materials/analysis tools: MIAO Fa Ming, CHEN Teng, and LIU Ye.
doi: 10.3967/bes2018.064
-
Abstract: Chinese ferret badger (FB)-transmitted rabies is a serious threat to public health in southeast China. Although mostly associated with dogs, the rabies virus (RABV) presents genetic diversity and has a significantly wide host range in China. Instead of the dog- and wildlife-associated China Ⅱ lineage in the past decades, the China Ⅰ lineage has become the main epidemic group hosted and transmitted by dogs. In this study, four new lineages, including 43 RABVs from FBs, have been classified within the dog-dominated China Ⅰ lineage since 2014. FB RABVs have been previously categorized in the China Ⅱ lineage. Moreover, FB-hosted viruses seem to have become the main independent FB-associated clade in the phylogenetic tree. This claim suggests that the increasing genetic diversity of RABVs in FBs is a result of the selective pressure from coexisting dog rabies. FB transmission has become complicated and serious with the coexistence of dog rabies. Therefore, apart from targeting FB rabies, priority should be provided by the appropriate state agencies to perform mass immunization of dog against rabies.
-
Figure 1. Maximum likelihood phylogenetic tree based on the complete nucleoprotein gene of rabies viruses isolated from China, rooted with a RABV virus isolated from a silver-haired bat in USA.
-
[1] Liu Y, Zhang HP, Zhang SF, et al. Rabies outbreaks and vaccination in domestic camels and cattle in Northwest China. PLOS Negl Trop Dis, 2016; 10, e0004890. doi: 10.1371/journal.pntd.0004890 [2] Li R, Li Y, Wen S, et al. Immunogicity and safety of purified chick-embryo cell rabies vaccine under Zagreb 2-1-1 or 5-dose Essen regimen in Chinese children 6 to 17 years old and adults over 50 years:a randomized open-label study. Hum Vaccin Immunother, 2015; 11, 435-42. doi: 10.4161/21645515.2014.994460 [3] Wang L, Tang Q, Liang G. Rabies and rabies virus in wildlife in mainland China, 1990-2013. Int J Infect Dis, 2014; 25, 122-9. doi: 10.1016/j.ijid.2014.04.016 [4] Zhang S, Tang Q, Wu X, et al. Rabies in ferret badgers, southeastern China. Emerg Infect Dis, 2009; 15, 946-9. doi: 10.3201/eid1506.081485 [5] Liu Y, Zhang S, Wu X, et al. Ferret badger rabies origin and its revisited importance as potential source of rabies transmission in Southeast China. BMC Infect Dis, 2010; 10, 234. doi: 10.1186/1471-2334-10-234 [6] Chiou HY, Hsieh CH, Jeng CR, et al. Molecular characterization of cryptically circulating rabies virus from ferret badgers, Taiwan. Emerg Infect Dis, 2014; 20, 790-8. doi: 10.3201/eid2005.131389 [7] Tao XY, Tang Q, Rayner S, et al. Molecular phylodynamic analysis indicates lineage displacement occurred in Chinese rabies epidemics between 1949 to 2010. PLOS Negl Trop Dis, 2013; 7, e2294. doi: 10.1371/journal.pntd.0002294 [8] Zhao J, Liu Y, Zhang S, et al. Molecular characterization of three ferret badger (Melogale moschata) rabies virus isolates from Jiangxi province, China. Arch Virol, 2014; 159, 2059-67. doi: 10.1007/s00705-014-2044-0 [9] Velasco-Villa A, Mauldin MR, Shi M, et al. The history of rabies in the Western Hemisphere. Antiviral Res, 2017; 146, 221-32. doi: 10.1016/j.antiviral.2017.03.013 [10] Wang H, Fuller TK. Ferret badger Melogale moschata activity, movements, and den site use in southeastern China. Acta Theriol, 2003; 48, 73-8. doi: 10.1007/BF03194267 -

 点击查看大图
点击查看大图
图(1)
计量
- 文章访问数: 2199
- HTML全文浏览量: 792
- PDF下载量: 80
- 被引次数: 0

 Quick Links
Quick Links