
-
Campylobacter jejuni, one of the most common causes of acute human gastroenteritis worldwide, is an important foodborne pathogen[1]. In addition, C. jejuni has received increasing attention for its association with Guillain-Barre and Miller-Fisher syndromes[2]. Although relatively little is known about the molecular mechanisms underlying C. jejuni pathogenesis, it is widely accepted that flagellar motility is necessary for gastrointestinal tract colonization. Efficient colonization is a prerequisite for C. jejuni pathogenesis, and only motile strains have been recovered from human volunteers[3,4].
NCTC11168 has been extensively used in basic research on Campylobacter and represents a standard strain for studies on the assembly or conformation of flagella. A comprehensive analysis of flagellin glycosylation in Campylobacter jejuni NCTC 11168 revealed its importance in host colonization[5]. The genes rpoN, flaA, fliA, flhA, flgA, motA, and flgR have been shown to play important roles in flagellar biosynthesis in Campylobacter jejuni[6-10].
FlhA and flhB are the most well-characterized subunits of the membrane-inserted components of the flagellar export apparatus[11-16]. FlhA encodes a ~700 residue protein that is essential for flagellum biogenesis and contains an N-terminal integral membrane domain (amino acids 1–350), predicted to contain either six or eight membrane-spanning helices. In the absence of crystal structure experiments on the FlhAC protein from C. jejuni, the crystal structure of the cytoplasmic domain of FlhA from Helicobacter pylori was used[17].
Compared to other enteric pathogenic bacteria, such as Salmonella typhimurium and Escherichia coli, C. jejuni does not utilize carbohydrates as a food source. Campylobacter have been labeled as asaccharalytic microorganisms since early biological investigations on substrate consumption demonstrated that C. jejuni is unable to utilize glucose or other six-carbon carbons as a carbon source[1,18].
The aim of the present study was to elucidate the genetic mechanism of flagellar loss in NCTC11168 using experimental laboratory passages and evaluate phenotypic variations in the mutant. The genomes of the mutant isolate CJ1Z (NCTC11168, flhA mutant strain, lawn colony) and wild-type isolate CJ2S (NCTC11168, flhA complemented strain, normal colony) were compared ad hoc in this study. We identified 15 single nucleotide polymorphisms (SNPs) between the two genomes as well as a motility-related gene flhA mutation. An insertion in the flhA gene was identified, which has been shown to affect the assembly or conformation of the flagella. Phenotypic variations in the FlhA mutant and their predicted functions are described in this study.
-
Campylobacter jejuni NCTC11168 is a well-characterized, sequenced strain that is widely used in infection studies. The frozen stock from which all the strains were derived was originally obtained from the American Type Culture Collection (ATCC 700819). Strain NCTC11168 was used for the laboratory evolution experiments in this study. C. jejuni cultures were grown from freezer stocks on Campylobacter agar base (Karmali) plates with 5% (v/v) sheep blood for 48 h at 37 °C in a microaerophilic environment (5% O2, 10% CO2, and 85% N2).
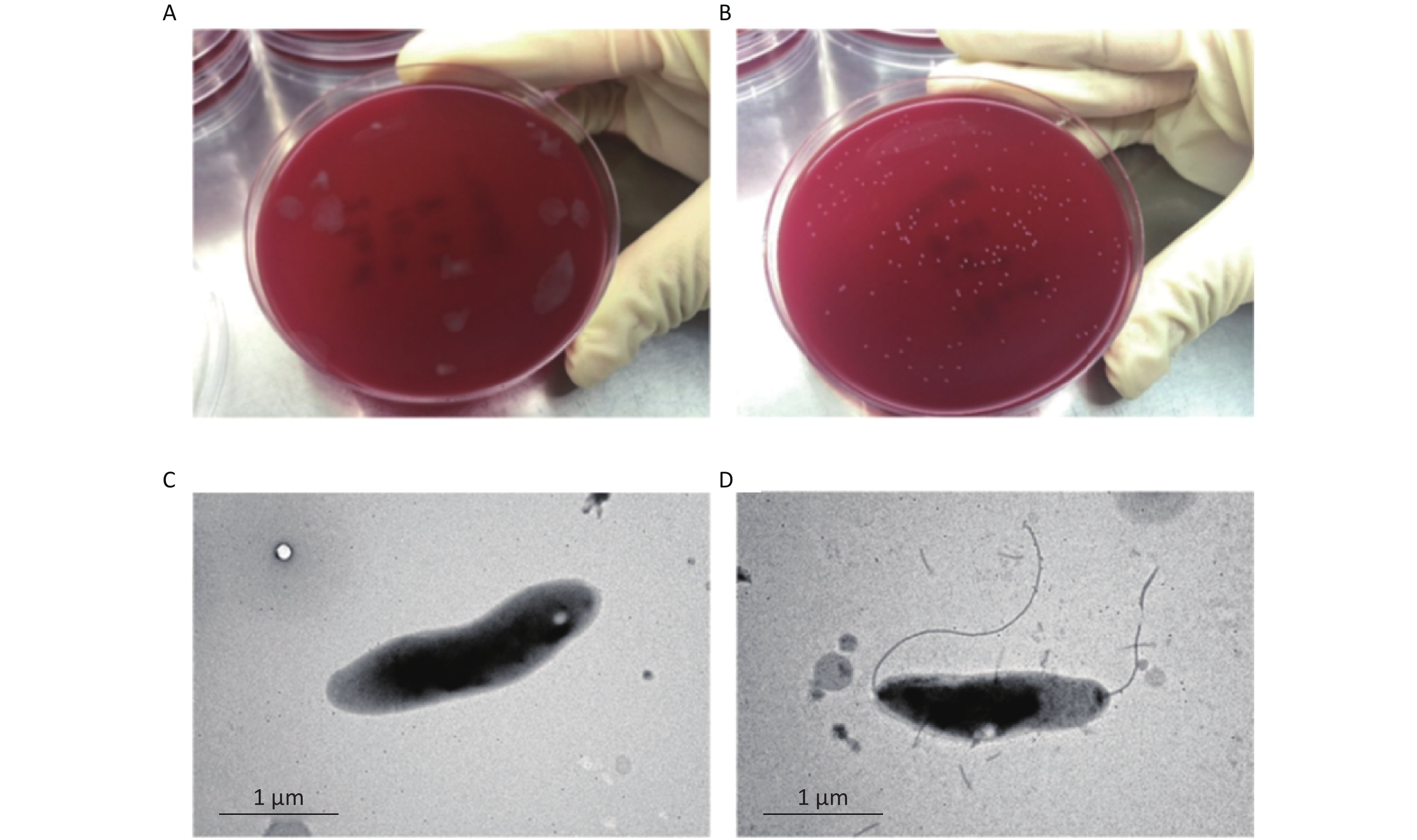
Colony variants emerged during serial passage by subculturing Campylobacter every 48 h at 37 °C in a microaerophilic environment for 30 d, named strain CJ1Z. We characterized the variant colonies and identified two distinct phenotypes: lawn colonies (CJ1Z, flhA mutant strain) and normal colonies (CJ2S, flhA complemented strain) (Figure 1A and 1B).
Figure 1. Difference in single colony and flagellation of flhA mutants. (A) CJ1Z in BHI agar following microaerobic growth; (B) CJ2S in BHI agar following microaerobic growth; (C, D) TEM was used to visualize the flagella of CJ1Z and CJ2S. The panels show representative TEM images for cells with: (C) no flagella, CJ1Z is represented; (D) bipolar flagella, CJ2S is represented.
-
The motility of C. jejuni strains was measured as described by Kalmokoff et al. with some modifications[19]. Bacteria were cultured on Mueller–Hinton agar (MH agar) plates for 48 h in PBS, and the optical density at 600 nm (OD600) was adjusted to 0.5. Then, 10 mL of the bacterial suspension was inoculated into the center of a motility agar plate (MH broth containing 0.4% nutrient agar). Each plate was incubated upright for 48 h at 37 °C, after which it was photographed, and the halo diameter was recorded.
-
C. jejuni grown on Campylobacter agar base (Karmali) was resuspended in Mueller-Hinton broth to an OD600 of 0.2 and grown for 4 h. The bacteria were diluted to an OD600 of 1 and collected by gentle centrifugation. The pellets were gently resuspended in a 2.5% (v/v) glutaraldehyde solution for fixation. Fixation was completed by incubating strains for 1 h at 4 °C. CJ1Z and CJ2S strains were stained with 1% (w/v) uranyl acetate and visualized using a JOEL 1,200× transmission electron microscope at 80 kV.
-
CJ1Z and CJ2S strains cultured at 37 °C on Campylobacter agar base (Karmali) under microaerophilic conditions for 48 h were inoculated into a 50 mL MH broth to a final OD600 of 0.02. The OD600 was recorded every 2 h for 30 h. At each time point, the turbidity of the medium was measured at 600 nm.
-
The ability of C. jejuni strains to form biofilms was measured using crystal violet staining[8]. Briefly, bacteria cultured for 20 h were harvested and diluted in MH broth to an OD600 of 0.025, after which CJ1Z and CJ2S strains were inoculated into a 96-well plate with 1 mL suspension at 37 °C under microaerobic conditions. After incubation for 24, 48, and 72 h, the plates were washed with PBS and dried at 55 °C for 30 min. Next, 1 mL of 1% crystal violet solution was added to each well to stain the formed biofilms. After staining at room temperature for 5 min, the plates were rinsed thoroughly with PBS thrice to remove unbound crystal violet. The plates were subsequently dried at 55 °C for 15 min, and the biofilms were quantified by determining the OD570 values after the remaining crystal violet was solubilized in a solution composed of 80% ethanol and 20% acetone.
-
The autoagglutination assay was performed as previously described[20]. C. jejuni strains were grown for 20 h prior to use, after which they were suspended in PBS (pH7.5) and adjusted to an OD600 of 1.0. Then, 4 mL aliquots of the suspension were transferred to a series of sterile glass tubes. The bacterial solution was placed at room temperature (25 °C), 200 μL of the bacterial suspension was carefully removed from the top of each tube every 1 h, and the OD600 was measured to evaluate the degree of autoagglutination (AAG). Bacterial cells that strongly agglutinate do not remain in the upper part of the supernatant but rather fall to the bottom of the tube, leading to a decreased OD600 value.
-
DNA extraction was performed using the Qiagen QIAamp DNA Mini Kit according to the manufacturer’s instructions. Genome sequences were obtained from the Beijing Genomics Institute. Here, 42 draft genomes were sequenced using the Illumina HiSeq 2500Xten platform (Illumina Inc., San Diego, CA, USA), generating reads of 300 bp in length. Complete sequencing was performed using PacBio, and long reads were analyzed using the pipelines provided by the SMRT Portal software. Clean reads were assembled using SOAPdenovo (SOAPdenovo 2.04,
http://soap.genomics.org.cn/soapdenovo.html ). The assembled genomes were annotated to predict the genes and their functions using Prokka V1.13.3. The genomes of strains CJ1Z and CJ2S were submitted to NMDC, and the accession numbers were assigned as NMDC60029075 and NMDC60029076. Whole genome sequencing data from the mutant and wild strains were compared by mapping against the reference strain C. jejuni NCTC11168 genome sequence (RefSeq: NC_002163.1) for all types of mutations using Mummer. -
In the present study, we investigated whether the flagellar structure of C. jejuni NCTC11168 is completely inhibited or affected in a more subtle manner. We examined two colony morphologies of the strains (CJ1Z and CJ2S) by transmission electron microscopy and found noticeable differences. The CJ1Z strain had no flagella but contained a spiral shape (Figure 1C). In contrast, the CJ2S strain had bipolar flagella (Figure 1D). Together, these observations show that the variant colony was accompanied by flagellar synthesis or assembly changes.
-
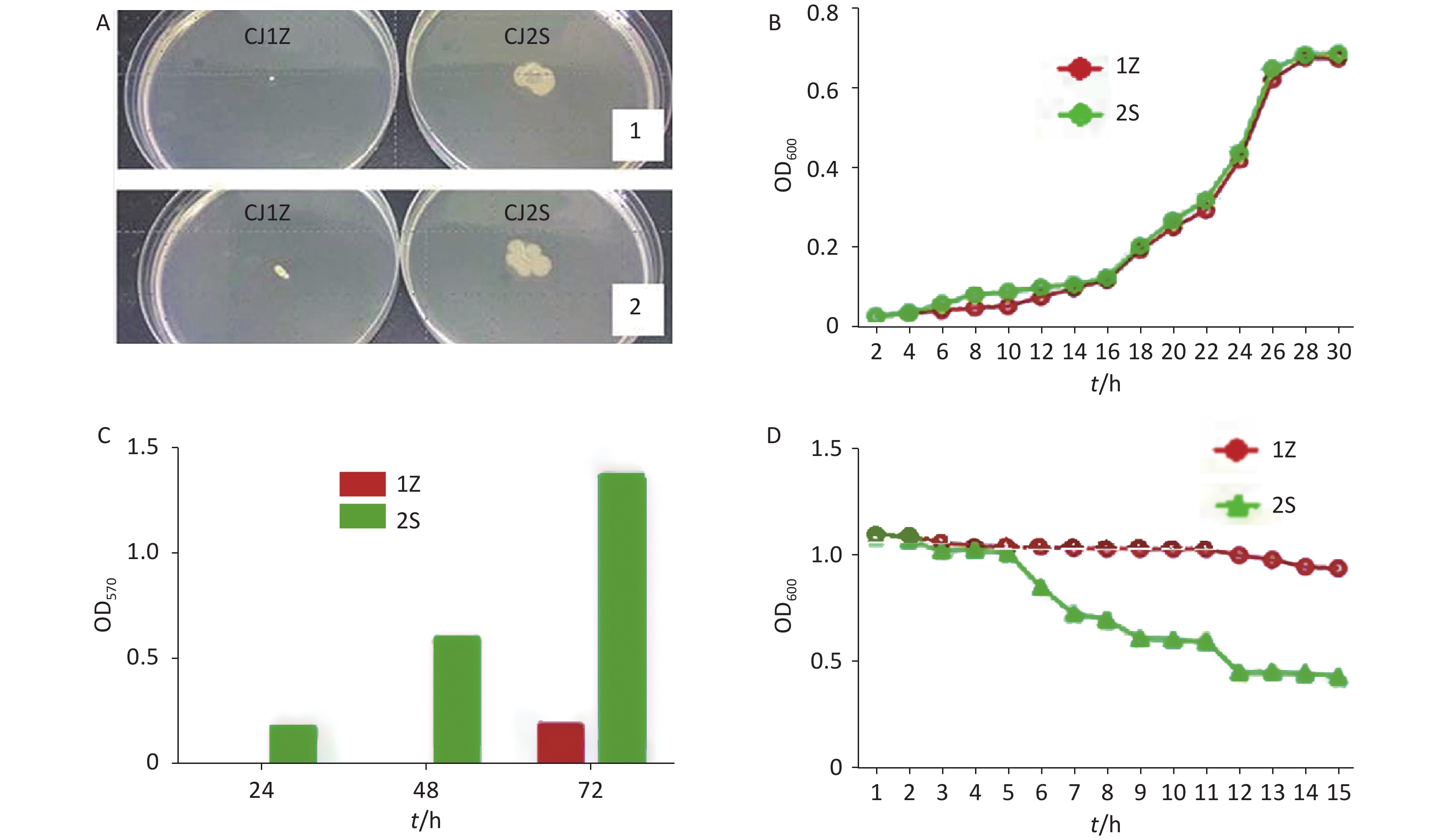
CJ1Z and CJ2S isolates were assessed for their swimming motility in MH agar broth. The data (Figure 2A) showed that the mutant CJ1Z strain had a motility defect compared to the CJ2S strain over the course of 48 h. This suggests that the loss of flagella may be a major contributor to the motility loss of C. jejuni NCTC11168.
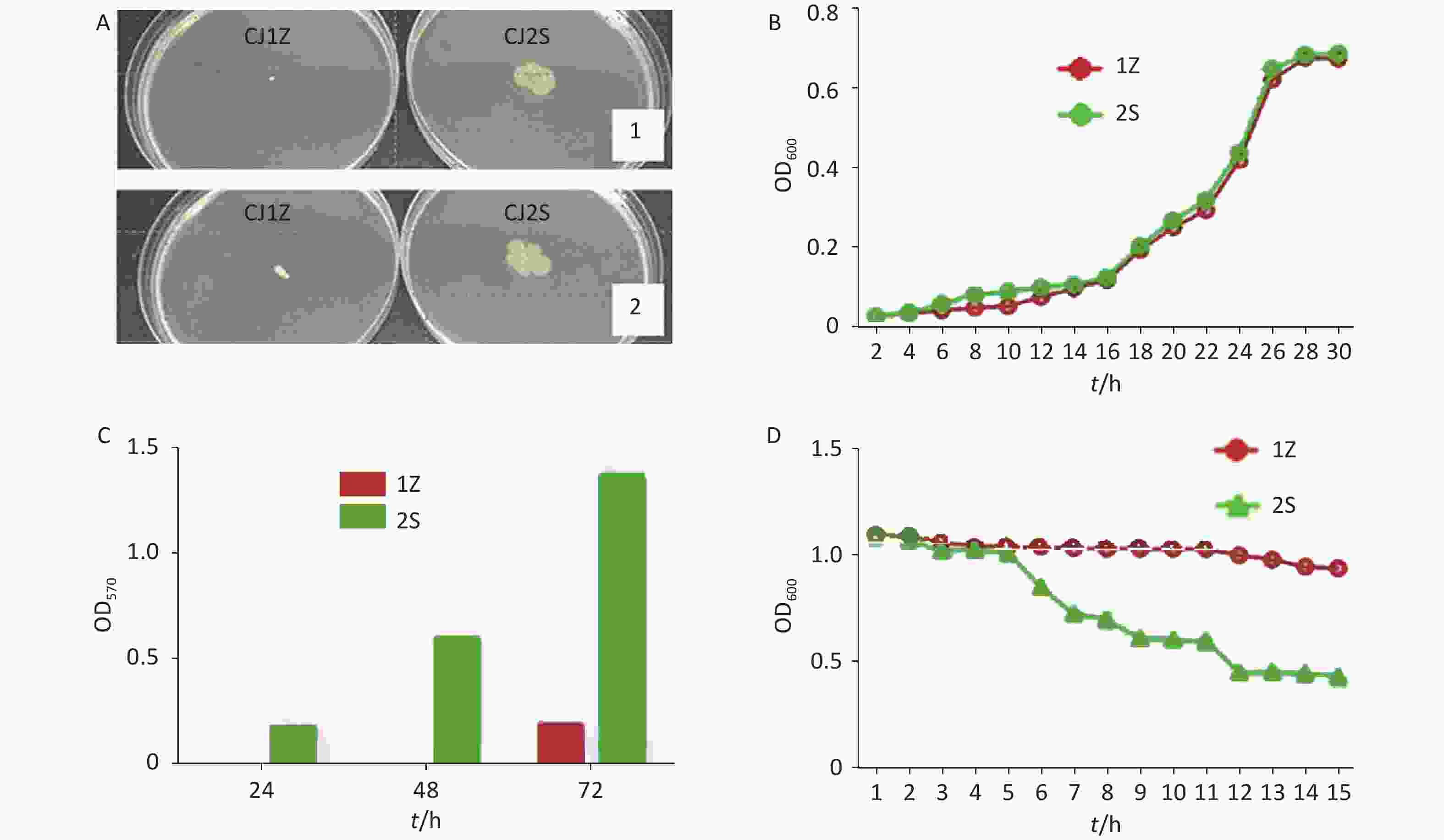
Figure 2. Motility, growth curves, biofilm formation, and autoagglutination of CJ1Z and CJ2S. (A) Motility haloes of C. jejuni CJ1Z (flhA mutant) and CJ2S on soft agar; (B) Growth curve of C. jejuni CJ1Z and CJ2S. C. jejuni CJ1Z and CJ2S were grown in MH broth, with 100 μL of samples withdrawn at designated time point to examine growth. The CJ1Z strain (red cycle) and CJ2S strain (green cycle) are represented by each sample; (C) Formed biofilms of C. jejuni CJ1Z and CJ2S. The formed biofilms were stained with crystal violet, and biofilm formation was quantitated by determining the OD570 value; (D) Autoagglutination abilities of the CJ1Z strain (hollow circle) and CJ2S strain (hollow triangle).
The observed growth curves indicated that mutation of the colony did not influence the normal growth of C. jejuni. When cultured in MH broth, the OD600 values for C. jejuni CJ1Z and CJZS strains were nearly the same at different times, with complemented differences observed, suggesting that these strains have a similar growth rate (Figure 2B).
The biofilms formed by the strains were quantified after static culture for 24, 48, and 72 h. As shown in Figure 2C, the biofilms formed by the CJ2S strain continuously increased over time; however, the flhA mutant CJ1Z strain showed a defect in biofilm formation, as the OD570 values of the tested strains across all assayed time points decreased compared to those observed for the CJ2S strain.
The AAG ability of C. jejuni strains was compared after incubation each hour (Figure 2D). In the first 5 h, there was no significant difference in the AAG ability between the CJ1Z and CJ2S strains. The influence of colony variants on the AAG of C. jejuni became noticeable after incubation for 5 h. The OD600 of C. jejuni CJ1Z was higher than that of the CJ2S strain, and this effect increased over time as the difference was more significant at 12–15 h.
-
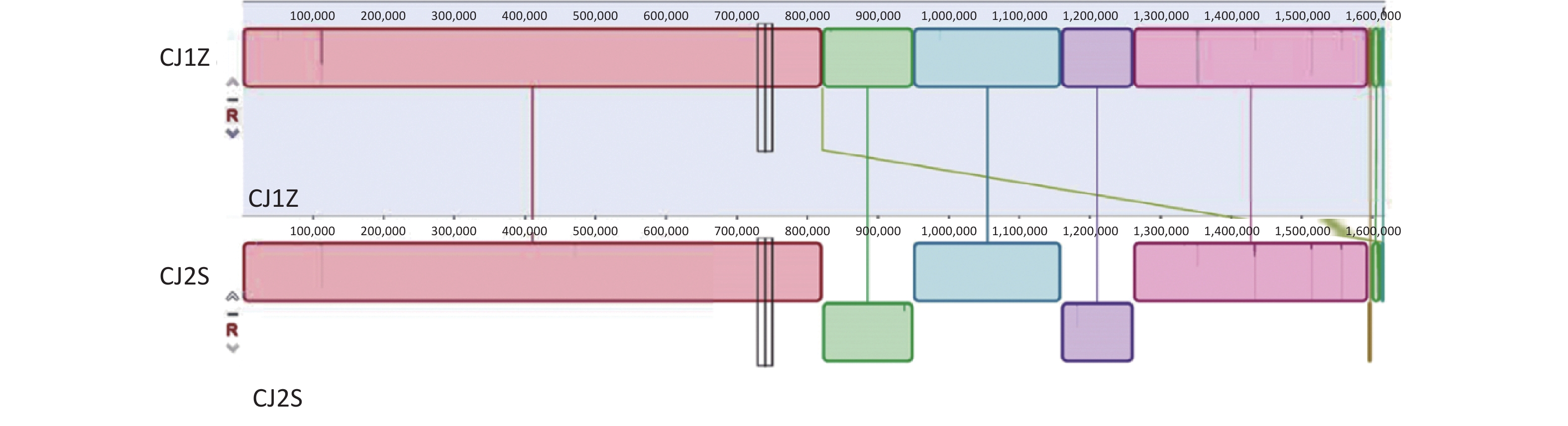
We identified 15 SNPs in the two genomes (5 nonsynonymous, 1 stop-gain, 1 synonymous, and 8 noncoding), among which 7 were located in the open reading frame (ORF) spacer region, and 2 INDELs were located in the ORF, which were distributed in 9 genes (Supplementary Table S1, available in www.besjournal.com). A total of one base pair insertion mutation (termination) was found in the motility-related gene flhA (Table 1). Additionally, eight distinct mutations were identified in seven non-motility-related genes (Figure 3 and Table 1). However, multiple other mutations were found in this strain and need to be acknowledged as potential influences on the overall phenotype of the bacterium.
Position CDS Mutation Mutation type Motility-related Strain CJ1Z Scaffold1 (51967) flhA +G (R721*) Insertion (Termination) Non-motility-related Strain CJ1Z Scaffold4 (5943) hypothetical protein +C Insertion Scaffold4 (12612) hypothetical protein ∆TATCT Deletion Scaffold15 (2182) hisC_3 +C Insertion Strain CJ2S Scaffold1 (79809) hypothetical protein G→A Transition Scaffold1 (471226) hypothetical protein +C Insertion Scaffold4 (117178) hypothetical protein +28 bp Insertion Scaffold5 (36939) kdpA_3 ∆G Deletion Scaffold13 (6301) hypothetical protein +C Insertion Note. + indicates the insertion of the following base(s); for longer insertions, + is followed by the number of bases inserted. ∆ indicates deletion of the following base(s); *symbolizes a stop codon. Table 1. All mutations predicted in the flhA mutant and complemented wild strains
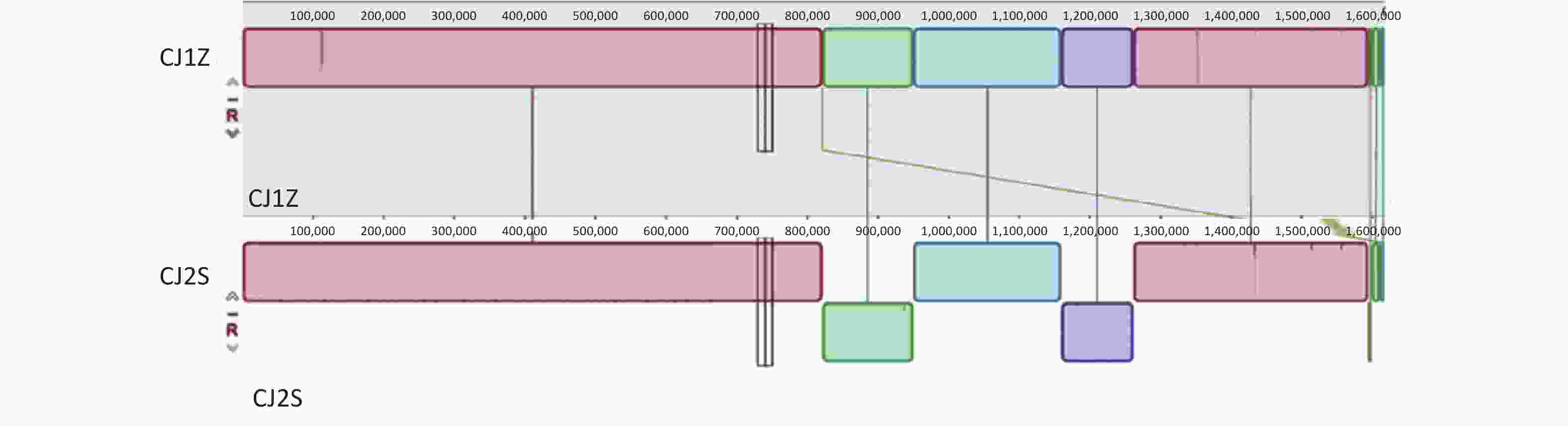
Figure 3. Comparison of complete genomes of C. jejuni using Mauve alignment tool. (A) Comparison of complete genomes of C. jejuni CJ1Z and CJ2S strains using strain CJ1Z as the reference genome.
Scaffold CJ1Z Base Base CJ2S CDS Scaffold1 51967 C − 51966 flhA Scaffold1 79810 G A 79809 hypothetical protein Scaffold1 471433 − C 471226 hypothetical protein Scaffold13 6300 − C 6301 hypothetical protein Scaffold13 9164 C − 9164 intergenic region Scaffold15 2182 C − 2372 hisC3 Scaffold26 298 C − 297 intergenic region Scaffold27 292 A C 292 intergenic region Scaffold27 293 G A 293 intergenic region Scaffold27 295 T A 295 intergenic region Scaffold4 5943 C − 123798 hypothetical protein Scaffold4 12536 − T 117178 hypothetical protein Scaffold4 12536 − T 117179 hypothetical protein Scaffold4 12536 − C 117180 hypothetical protein Scaffold4 12536 − T 117181 hypothetical protein Scaffold4 12536 − A 117182 hypothetical protein Scaffold4 12536 − T 117183 hypothetical protein Scaffold4 12536 − C 117184 hypothetical protein Scaffold4 12536 − T 117185 hypothetical protein Scaffold4 12536 − N 117186 hypothetical protein Scaffold4 12536 − N 117187 hypothetical protein Scaffold4 12536 − N 117188 hypothetical protein Scaffold4 12536 − N 117189 hypothetical protein Scaffold4 12536 − N 117190 hypothetical protein Scaffold4 12536 − N 117191 hypothetical protein Scaffold4 12536 − N 117192 hypothetical protein Scaffold4 12536 − N 117193 hypothetical protein Scaffold4 12536 − N 117194 hypothetical protein Scaffold4 12536 − N 117195 hypothetical protein Scaffold4 12536 − N 117196 hypothetical protein Scaffold4 12536 − N 117197 hypothetical protein Scaffold4 12536 − N 117198 hypothetical protein Scaffold4 12536 − N 117199 hypothetical protein Scaffold4 12536 − N 117200 hypothetical protein Scaffold4 12536 − N 117201 hypothetical protein Scaffold4 12536 − N 117202 hypothetical protein Scaffold4 12536 − N 117203 hypothetical protein Scaffold4 12536 − N 117204 hypothetical protein Scaffold4 12536 − N 117205 hypothetical protein Scaffold4 12612 − T 117097 hypothetical protein Scaffold4 12612 − A 117098 hypothetical protein Scaffold4 12612 − T 117099 hypothetical protein Scaffold4 12612 − C 117100 hypothetical protein Scaffold4 12612 − T 117101 hypothetical protein Scaffold5 36860 G − 36939 kdpA 3 Scaffold7 79979 − C 22919 intergenic region Scaffold7 79979 − C 22920 intergenic region Scaffold8 71610 − C 71611 intergenic region Table S1. All SNPs predicted in the mutant and wild strains
-
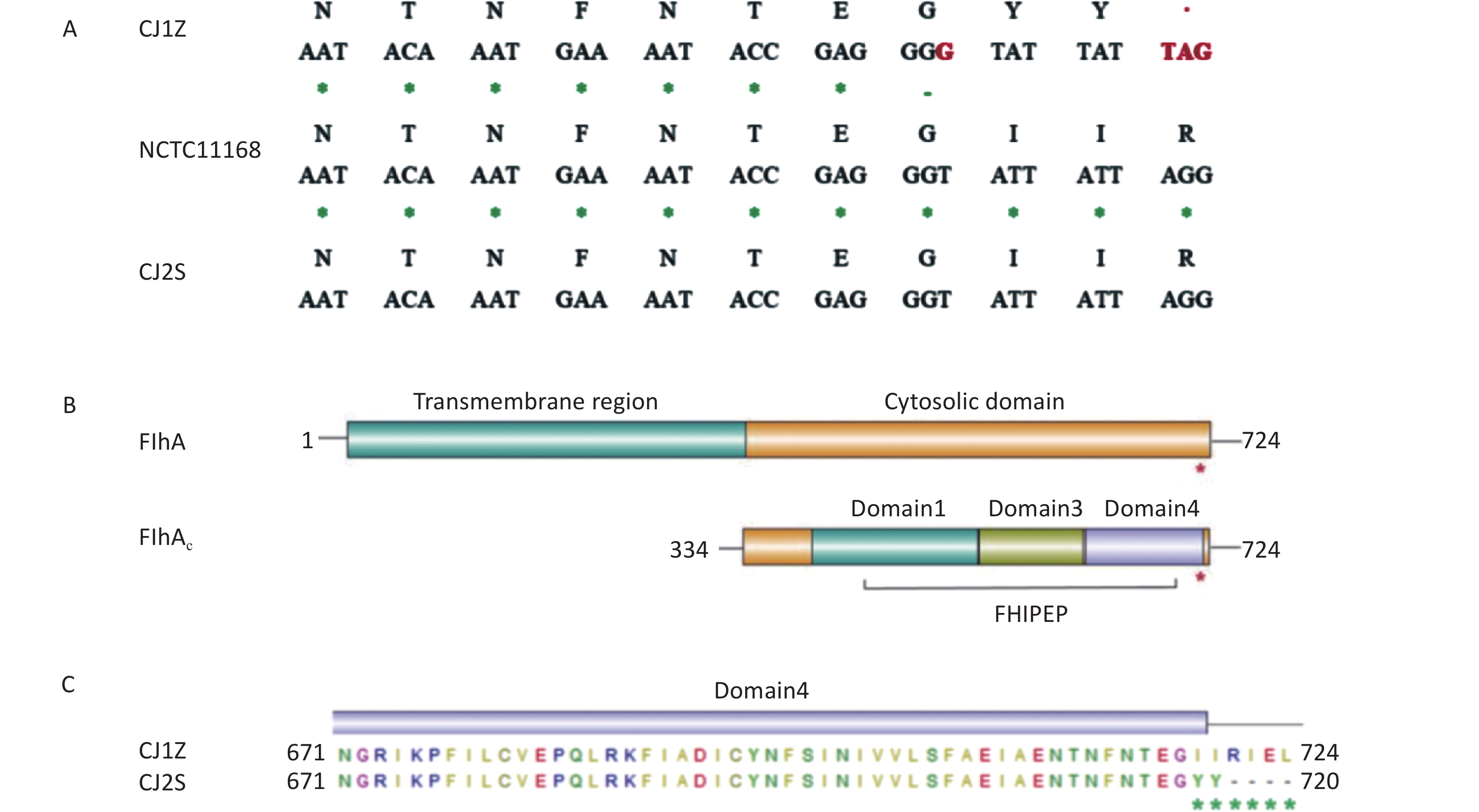
As part of our efforts to investigate the reason for the phenotypic variation in the CJ1Z strain, we found that strains CJ1Z and CJ2S have different flhA alleles. Specifically, the flhA allele in strain CJ1Z had a one-base-pair insertion within the 3’ end of the gene, which resulted in the formation of a premature stop codon (Figure 4A). FlhA consists of an N-terminal transmembrane (TM) domain with eight predicted TM α-helices and a large cytosolic domain at its C-terminal (FlhAC) (Figure 4B). The truncated FlhA protein produced by the strain CJ1Z had two I719Y/I720Y missense mutations and residues 721 to 724 deleted, compared to 724 amino acids for the full-length protein (Figure 4C). Thus, the sequence data suggest that strain CJ1Z has a null mutant flhA allele.
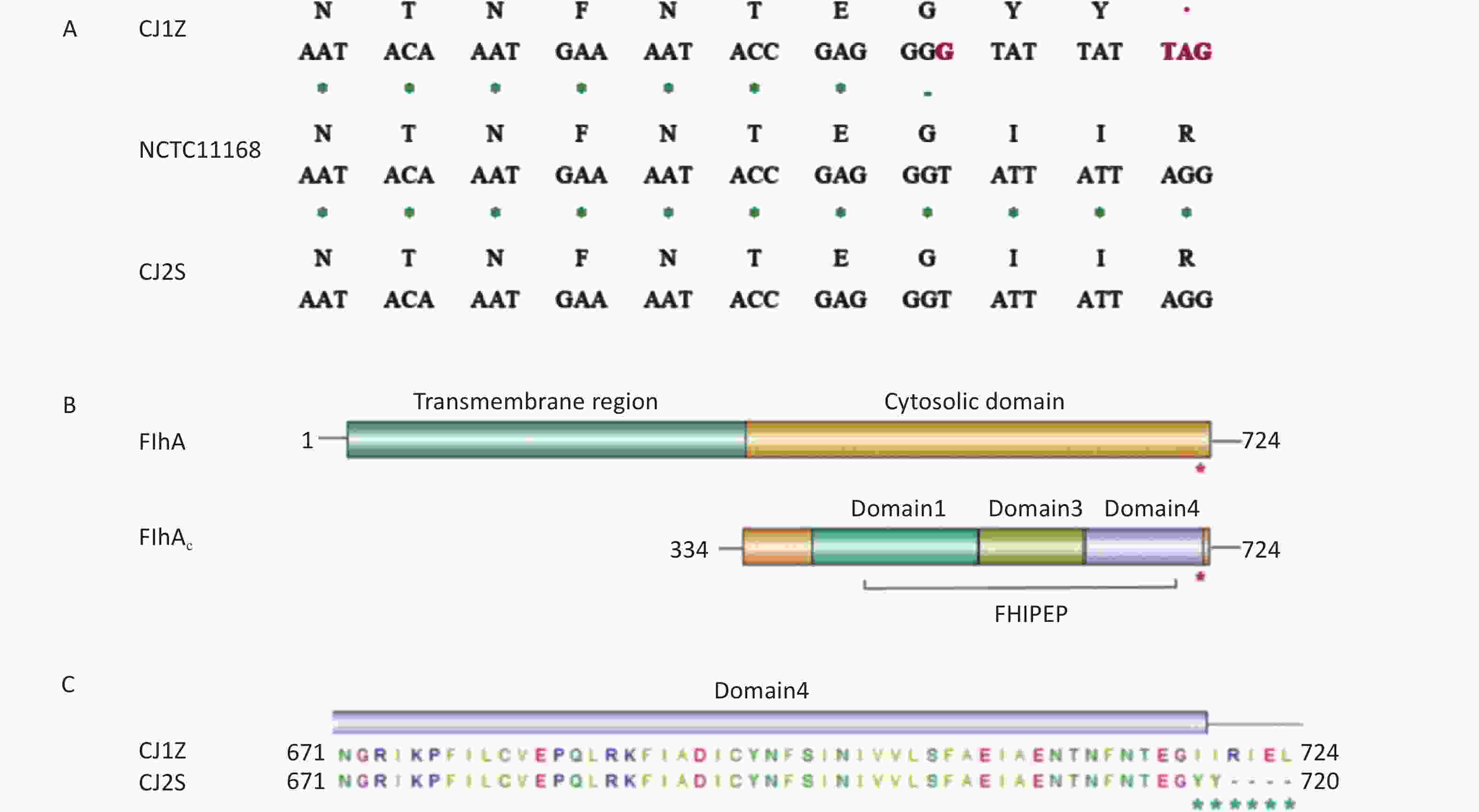
Figure 4. FlhA mutation in strain CJ1Z resulting in the early truncation of the protein. (A) Comparison of a section of the flhA gene and the translated product between the strains CJ1Z and CJ2S. The location of the 1-bp insert in strain CJ1Z is highlighted in red, as is the location of the stop codon that is subsequently introduced; (B) Domain structure of the FlhA protein. The location of the truncation in strain CJ1Z is highlighted with a red asterisk. The numbers correspond to amino acid positions; (C) Amino acid changes in strain CJ1Z. The numbers correspond to amino acid positions.
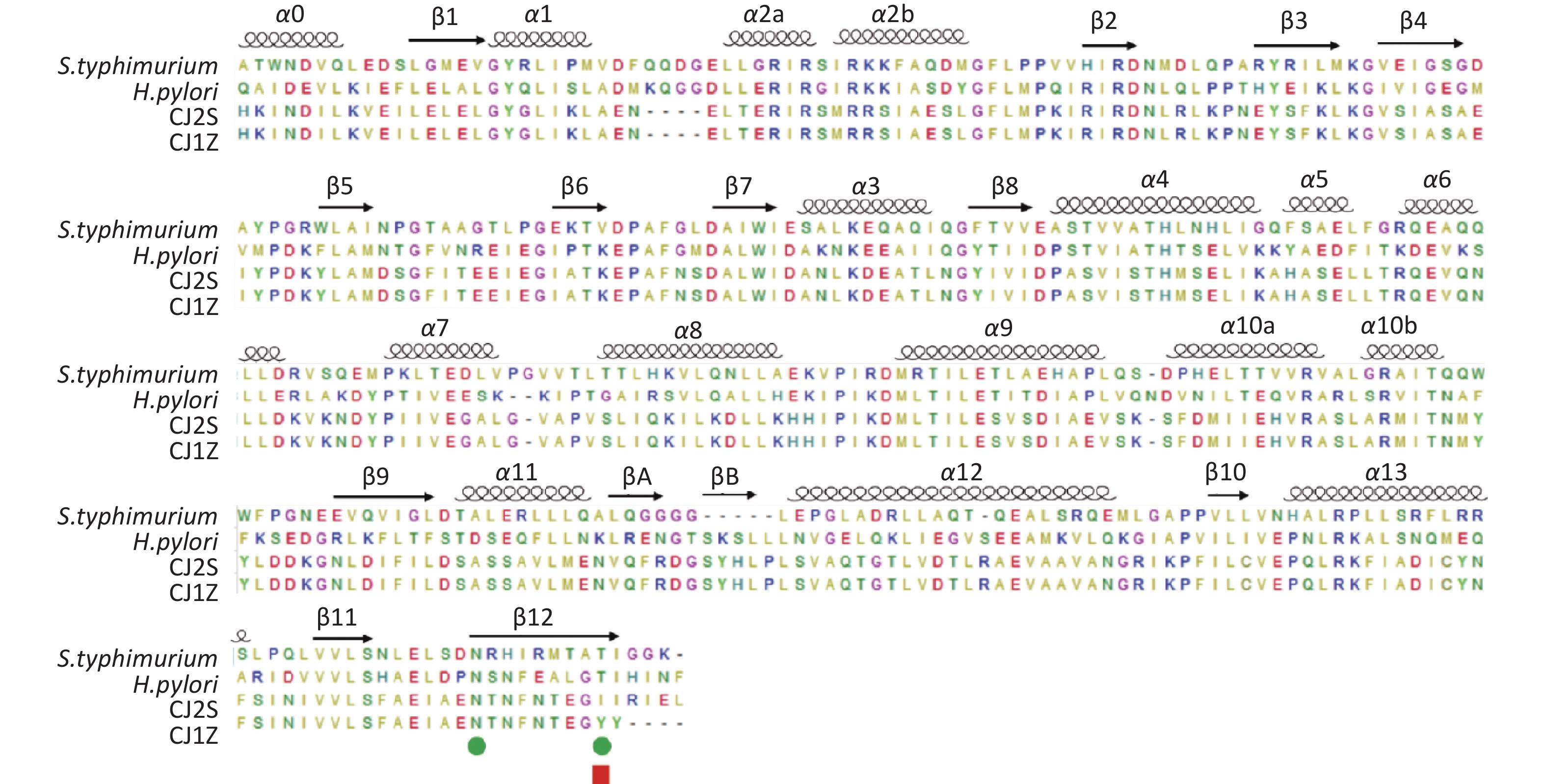
The crystal structures of the FlhAC protein from the CJ1Z and CJ2S strains were obtained using the SWISS-model (
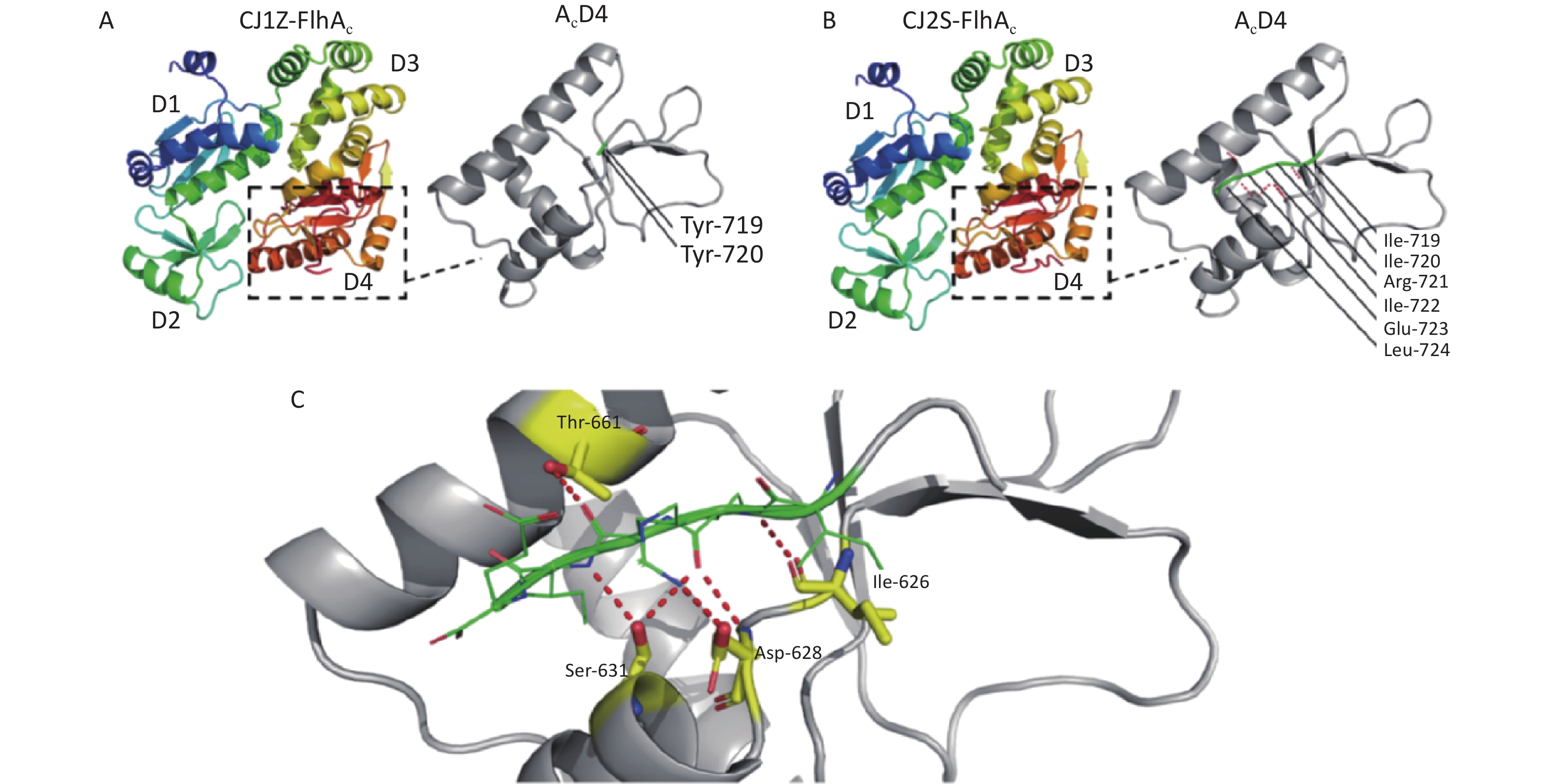
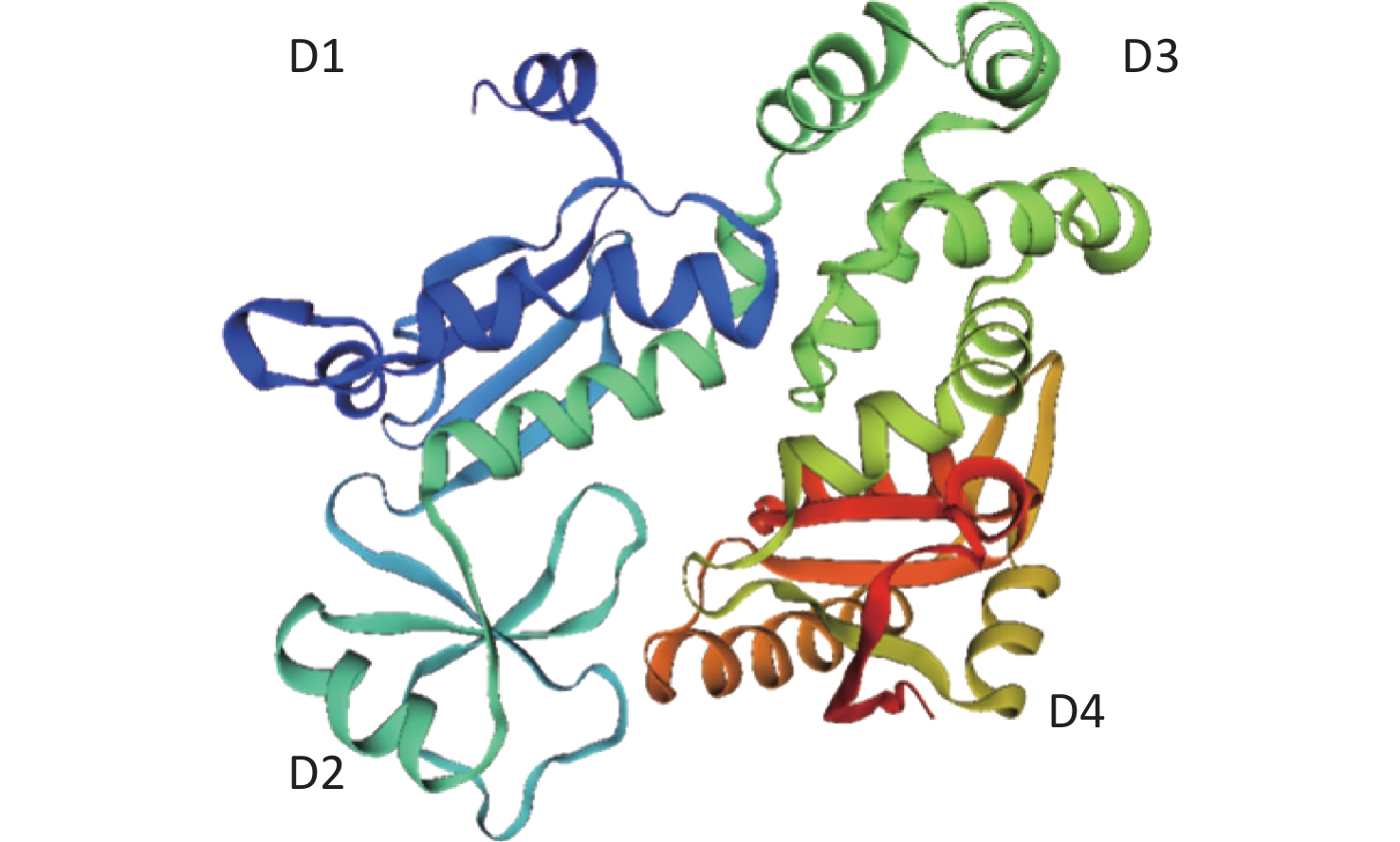
https://swissmodel.expasy.org/ )[21-25]. In the absence of crystal structure experiments on the FlhAC protein from C. jejuni, the crystal structure of the cytoplasmic domain of FlhA from Helicobacter pylori was used (Supplementary Figure S1, available in www.besjournal.com)[17]. The flhA mutation (FlhA719) is located in the chaperone-binding site of FlhAC (Figure 5A), raising the question of whether the flhA mutation affects the interaction of FlhAC with chaperone/filament-type export substrate complexes. By searching for hydrogen bonds between amino acids using PyMOL software, we propose that the H-bonds of the last six amino acids (or less) of FlhAC may stop the export of flagellar biosynthesis protein and affect the expression of the flagella (Figure 5C). Several temperature-sensitive Salmonella enterica serovar Typhimurium FlhAC missense mutants that cannot regrow flagella at restrictive temperatures (42 °C) have been reported. These mutants have been extensively analyzed using the recently reported S. Typhimurium FlhAC structure. The positions of these mutations are shown in Supplementary Figure S2 (available in www.besjournal.com), and two of the mutations (green circles) have already been reported in relation to the conserved salt bridges at the interface between the helical and C-terminal domains[11,26]. Campylobacter jejuni FlhA I719Y mutation, marked with a red square, was located at the same site as S. Typhimurium.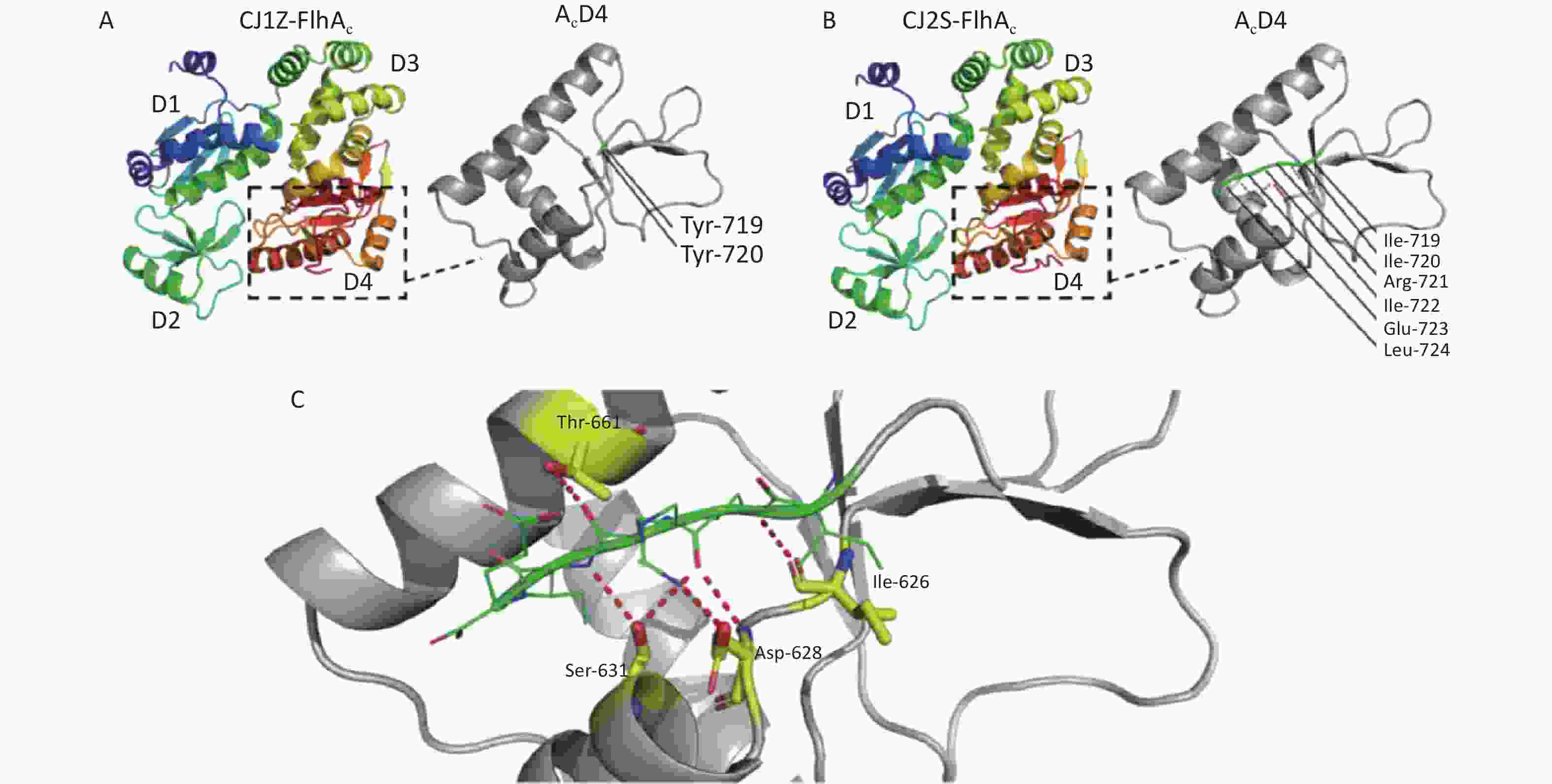
Figure 5. Location of the mutations in FlhA. (A, B) Crystal structure of C. jejuni FlhAC, which consists of four domains, D1, D2, D3, and D4. Strain CJ1Z revealed a native nucleotide insertion at amino acid position 719 (flhA719) that leads to the substitution of (A) Ile-719, Ile-720, Arg-721, Ile-722, Glu-723, and Leu-724 to (B) Tyr-719 and Tyr-720. (C) The last six amino acids (or less) of FlhA in strain CJ1Z exhibited hydrogen bonds with amino acid residues of FlhA.
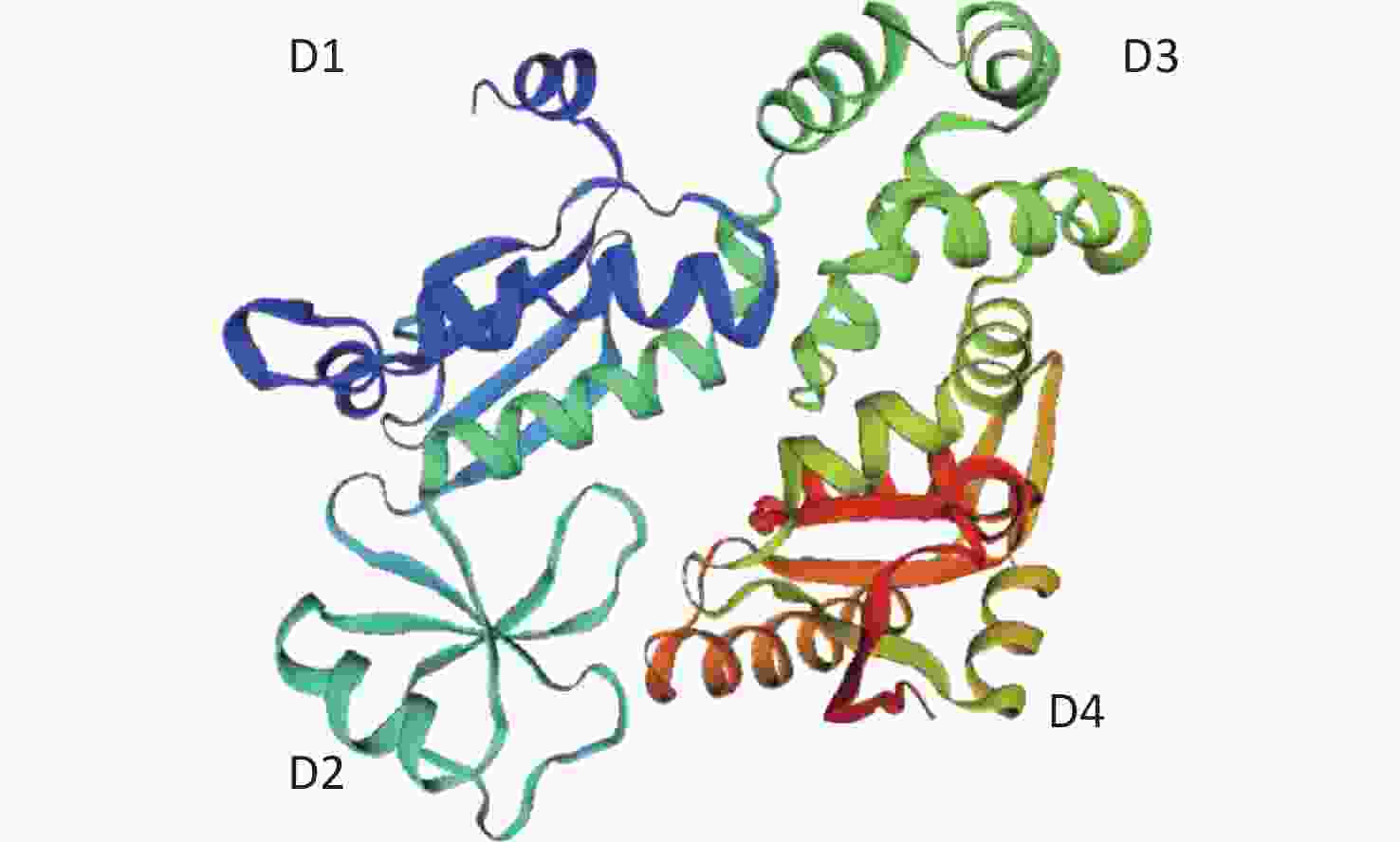
Figure S1. Structure of the cytoplasmic domain of FlhA from Helicobacter pylori (3my.1.A).
-
NCTC11168 is a C. jejuni strain that was isolated from a human infection in 1977[27] and was the first C. jejuni to be genome-sequenced[1,28]. It has been used as a standard strain for basic research on flagellar biosynthesis genes[7,8,19].
The present study aimed to determine the reasons for the loss of flagellar motility of C. jejuni strain NCTC11168 by experimental laboratory evolution. According to the Sher study, at early time points, the motility defect was often reversible; however, after 35 days of serial culture, motility was irreversibly lost in most cells in five independently evolved populations[9]. By observing the phenotypic variation of the strains, we found that C. jejuni strain CJ1Z serially passaged in a rich medium exhibited a lack of flagella. This is accompanied by a loss of motility, decrease in the ability to form biofilms, and autoagglutination. Flagella is essential for motility[7,8] and plays a significant role in biofilm formation by C. jejuni[8,29]. AAG is often a preliminary step in the formation of microcolonies, and in C. jejuni, AAG has been shown to be highly associated with flagellar expression[30,31]. In addition, the role of flagella in the virulence of Campylobacter spp. is clearly far more complex and intricate than simple motility[32].
The phenotypes of the flhA mutants in C. jejuni strain CJ1Z indicate that the last six amino acids (or less) of FlhA are required for flagellar assembly. Sequence analysis of the flhA gene in strain CJ1Z revealed a native nucleotide insertion at amino acid position 719 (flhA719) that led to the loss of Ile-719, Ile-720, Arg-721, Ile-722, Glu-723, and Leu-724 at the end of the cytosolic domain at its C terminal (FlhAC). This portion of FlhA was predicted to contain a stretch of 391 aa residues at the C-terminal of the protein (Figure 5). FlhAC belongs to the “flagellum/hypersensitive response/invasion” (FHIPEP) family of bacterial export proteins [33,34]. The hydrophilic C-terminal domain of FlhA extends into the cytoplasm[35,36] and is involved in the energy-dependent secretion of flagellar protein[37]. FlhAC interacts with FliHIJ and FlhB and with the substrate-chaperone complex (hook–filament junction protein FlgK/FlgN, filament capping protein FliD/FliT, and flagellin FliC/FliS[34,38-40]). Disruption of the flhA gene in gram-positive or gram-negative bacteria leads to non-motile cells, which lack flagella and are unable to export flagellar proteins[41-43]. Hara identified a non-motile isolate containing a D208A mutation in FlhA and suggested that this charged residue may be directly involved in PMF-driven protein export[44]. A mutation in the FlhA-binding site for substrate–chaperone complexes (A489E) was found to reduce the binding affinity of these complexes. We speculate that the FlhA719 mutation described in this study may influence the assembly or conformation of other components of the export apparatus, which in turn could modulate the expression of flagella.
Sher et al. demonstrated that the C. jejuni strain was rapidly attenuated by experimental laboratory evolution and C. jejuni in experimental laboratory evolution experiments undergo genomic instability with rapid gene attenuation, including insertions and deletions in the flhA gene. In addition, the loss of flagella observed in the present study in strain CJ1Z occurred in the flhA gene (FlhA I719Y mutation), which is inconsistent with the mutation described by Sher et al.[9] The binding site for the substrate–chaperone complex consists of α-helix and β-sheet structures and hydrogen bonds (H-bonds) between backbone N−H donors and CO acceptors[45]. H-bonds between the backbone N−H and CO moieties are ubiquitous in proteins, and their contribution to the structure is undisputed[46]. In this study, the last six amino acids (or less) of FlhAC in strain CJ1Z formed hydrogen bonds with the amino acid residues of FlhA. The absence of H-bonds may cause changes in the binding site that affect flagellar expression.
Carrillo[7] et al. provided evidence that the inactivation of flhA inhibits the transcription of both class II and class III flagellar genes and potential virulence factors regulated by σ28 and σ54 promoters, which are involved in virulence, flagellar biosynthesis, and carbohydrate modification. However, no differences were observed between the two variants in the gene sequence of flhA or other known regulators of flagellar biosynthesis. As reported in the literature, both the N-terminal and C-terminal domains of FlhA homologues have been conserved throughout evolution, and the cytoplasmic domains of FlhA and FlhB form complexes with FliH (regulator of FliI), FliI (ATPase), FliJ (general chaperone), and the substrate to be exported. Subsequently, through ATP hydrolysis by FliI, the substrate is translocated across the cytoplasmic membrane through the membrane-spanning domains of FlhA and FlhB, followed by the dissociation of FliH, FliI, and FliJ from the export apparatus[47]. In contrast, in this study, we compared the two variants of the genome-sequenced strains, CJ1Z and CJ2S, which indicated that the motility defect of CJ1Z was largely due to a change in the expression of flhA, the C-terminal domain of FlhA. Future work will focus on how the C-terminal domains of FlhA interact with other known regulators of flagellar biosynthesis to understand flhA regulation of flagellar assembly and virulence in Campylobacter jejuni. NCTC11168 is commonly used as a reference strain for studying the genes of the flagellar regulon and is accompanied by the loss of flagella after in vitro passage. The flagella-lacking strain had a truncated flhA gene and exhibited reduced biofilm formation, AGG, and different colony morphologies. In conclusion, flhA is more than a flagellar biosynthesis gene and plays an important role in controlling the vital functions of C. jejuni. Therefore, future evolutionary studies should be conducted to determine whether cell morphology is preserved. The effect of mutations on flagellar expression should be investigated to study the functions of genes related to flagellar regulation.
-
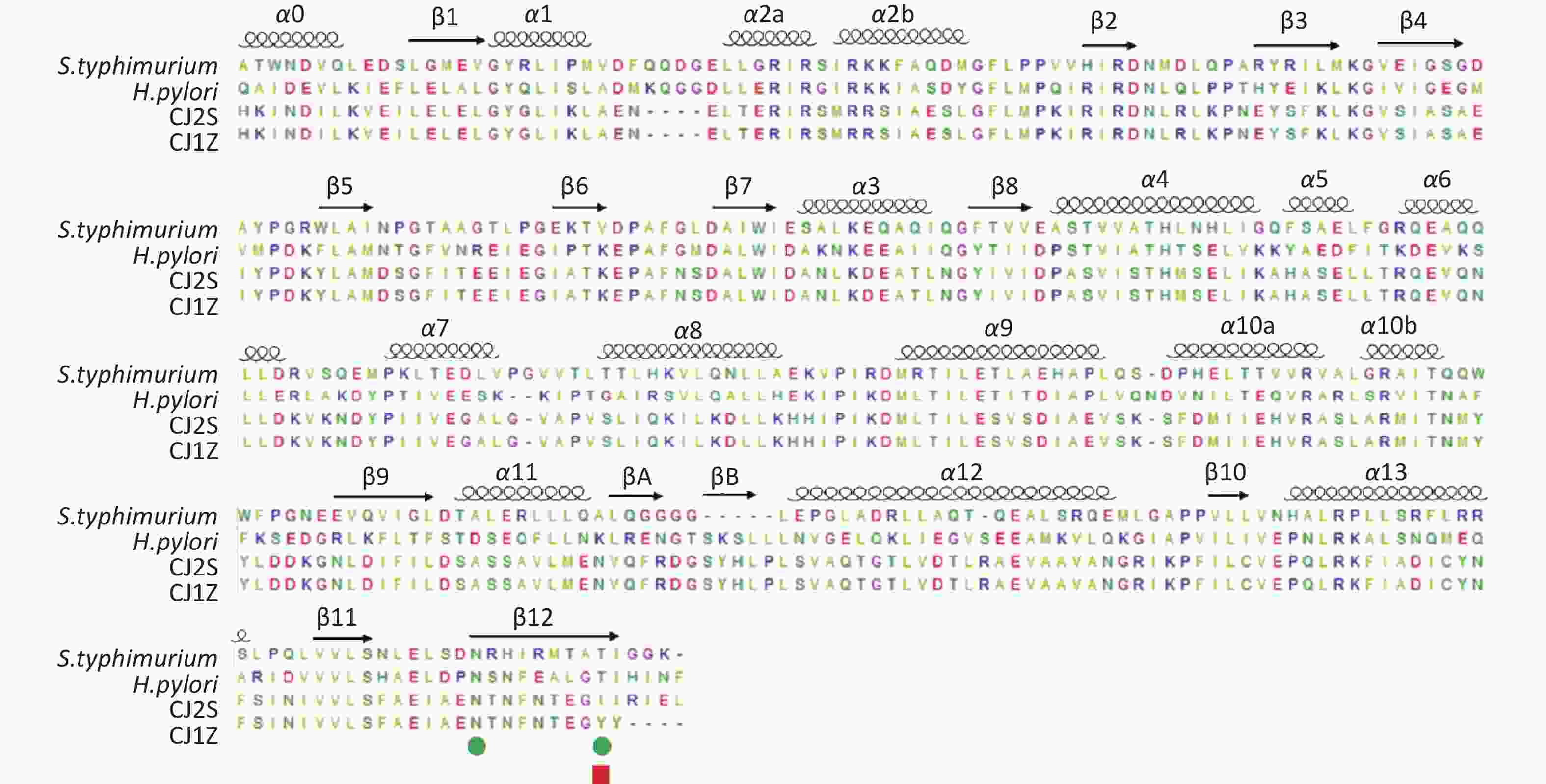
Figure S2. Structurally based alignment of FlhAC homologue sequences. These are labeled according to the Salmonella typhimutium and Helicobacter pylori FlhAC structure. Salmonella typhimutium FlhA deletion mutants are marked by green circles. Campylobacter jejuni FlhA I719Y mutation is marked with a red square.
HTML
Bacteria Strains and Growth Conditions
Motility Plates
Transmission Electron Microscopy
Growth Curves
Biofilm-formation Assay
Autoagglutination Assay
Whole Genome Sequencing for Strains CJ1Z and CJ2S
Variant Colony CJ1Z has no Flagellum by Transmission Electron Microscopy
Strain CJ1Z has a Motility Defect with Decreasing Autoagglutination and Biofilm-formation Abilities but no Defect in Growth Kinetics
Mutations were Detected in flhA Mutant and Complemented Strains
Different flhA Alleles in Strains CJ1Z and CJ2S
COMPETING INTERESTS The authors declare that they have no competing interests.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE Not applicable
 21317+Supplementary Materials.pdf
21317+Supplementary Materials.pdf
|

|

 Quick Links
Quick Links
 DownLoad:
DownLoad: