

 下载:
下载:




-
Lung cancer is one of the most frequently diagnosed cancers in terms of incidence and mortality, with approximately 1.38 million deaths every year worldwide, while breast cancer is the most prevalent cancer globally among women[1]. Radiotherapy is a widely utilized treatment modality for malignancies, especially in lung and breast cancer, but one of the main obstacles in these tumor types is radioresistance.
It is widely recognized that epigenetic dysregulation is an important mechanism underlying cancer development and progression. Histone post-translational modifications (PTMs), such as methylation, phosphorylation, acetylation, ubiquitination, and other epigenetic mechanisms, appear to function together in establishing and maintaining gene activity states, thus PTMs are essential for cell-fate determination and development[2]. Histone methylation in a lysine or arginine residue is highly conserved as it acts in a coordinated and orderly fashion to regulate cellular processes, including gene transcription, DNA replication, and DNA repair[3].
SET8, a member of the SET domain-containing family, is the only known lysine methyltransferase (KMT) that monomethylates lysine 20 of histone H4 (H4K20me1) in vivo[4]. SET8 methyltransferase activity has been closely connected with carcinogenesis. SET8-mediated p53K382me1 suppresses p53-dependent transcription activation in cancer cells[5]. SET8 degradation inhibits H4K20me1 and suppresses epithelial-mesenchymal transition (EMT) and invasion of breast cancer cells[6]. SET8 has also been implicated in many essential cellular processes, including cell cycle regulation, DNA replication, and the DNA damage response[7]. Moreover, disruption of endogenous SET8, accompanied by suppression of H4K20me1, leads to cell cycle defects, chromatin decondensation, and enlarged nuclei, indicating the essential role(s) of SET8 in DNA repair[8-10]. Although the evidence regarding the impact of SET8 on radiation treatment is inconclusive, there is significant interest in the precise effect of SET8 on radiotherapy.
Our clinical data analysis suggested that low levels of SET8 contribute to radiotherapy efficacy in patients with lung adenocarcinoma (LUAD) and invasive breast carcinoma (BRCA). Single-cell RNA sequencing (ScRNA-seq) revealed that low SET8 expression is associated with a reduced capacity for DNA repair. To further elucidate our observations, we used the SET8-deficient A549 and MCF7 cell lines exposed to radiation. SET8 deficiency was shown to enhance radiosensitivity by blocking DNA damage repair.
-
All patient treatment and clinical data were obtained from the cBioPortal database (
https://www.cbioportal.org )[11], which includes 32 studies consisting of approximately 10,000 specimens and representing 33 types of cancer from The Cancer Genome Atlas (TCGA)[12]. Published clinical and genomic data from the TCGA PanCancer LUAD cohort, including 61 and 402 late-stage cancer patients treated and not treated with radiotherapy, respectively. Five hundred forty-eight and 433 cancer patients were included in the BRCA cohort who were treated and not treated with radiotherapy, respectively. Kaplan-Meier survival curves were generated using the statistical software, GraphPad Prism. The non-parametric Mantel-Cox log-rank test was used to determine the statistical differences among different patient groups. The Pearson correlation coefficients (R) of gene expression relevance were calculated by GraphPad Prism. -
A549 (human lung carcinoma), MCF7 (human breast carcinoma), LLC1 (mouse lung carcinoma), and HEK 293T cells (human embryonic kidney) were obtained from the American Type Culture Collection (Manassas, VA, USA). A549 cells were cultured in Ham's F-12K medium (Gibco, Waltham, MA, USA), and MCF7, LLC1, and HEK 293T cells were cultured in Dulbecco’ Modified Eagle’s Medium (DMEM; Gibco) supplemented with 10% (v/v) fetal bovine serum (FBS; Hyclone, Logan, UT, USA) and 1% penicillin/streptomycin. Cells were cultured at 37 °C in a humidified atmosphere containing 5% CO2.
-
X-ray irradiation was delivered in a Faxitron RX-650 system (100 kVp and 5 mA; Faxitron Bioptics, Marlborough, MA, USA) at room temperature. The target of this instrument is wolframium (W) and the dose was 0.76 Gy/min. Carbon ions irradiation (linear energy transfer [LET]: 80 KeV/μm) was delivered at the Heavy Ion Research Facility of Lanzhou, Institute of Modern Physics (HIRFL Lanzhou, China) with a dose of 0.4 Gy/min.
-
The knockout cells were generated using lentivirus-mediated CRISPR/Cas9 technology. Single guide RNA (sgRNA) sequences targeting human and mouse SET8 are listed in Supplementary Table S1, available in www.besjournal.com. Double-stranded oligos encoding the sgRNA sequences were cloned into the BsmBI- (Thermal Fisher Scientific, Waltham, MA, USA) digested plasmid, LentiCRISPRv2 (deposited by Dr. Feng Zhang of MIT to Addgene, Watertown, MA, USA), which co-expresses Cas9 and sgRNA in the same vector. CRISPR lentivirus vectors were then produced in HEK 293T cells by psPAX2 and pMD2.G plasmids according to an established protocol by the Zhang laboratory[13]. To generate the knockout cell lines, target cells were infected with lentivirus and selected in 1 µg/mL of puromycin.
Table S1. sgRNA used in CRISPR/Cas9 knockout experiments
Target Sequence (5'–3') Human-SET8-sg1 TTTAGGAGAACGTATTTACC Human-SET8-sg2 GCACGTGACTACCTGCAGCT Mouse-SET8-sg1 AGTATCTGAGCAAAACCTAC Mouse-SET8-sg1 TCCGGGAACTCCTCCGCACA SET8 overexpression and negative control vector were purchased from GeneCopoeia (Guangzhou, China). For transient transfection, cells were plated on the day before transfection at a confluence of 30%−50% performed with Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions. The medium was replaced with fresh medium 6 h post-transfection. Cells were used in the following experiments after 48 h.
-
Cells were lysed in RIPA buffer (Beyotime, Shanghai, China) with protease inhibitor cocktail tablets (Roche, Basel, Basel-Stadt, Switzerland). Lysate total protein concentrations were determined using a protein assay kit (Bio-Rad, Hercules, CA, USA). Equal amounts of protein were denatured with loading buffer (Beyotime) at 100 °C for 10 min, then loaded onto a 10% SDS-PAGE gel for electrophoresis and transferred to a methanol-activated polyvinylidene fluoride membrane (Millipore, Burlington, MA, USA). The membrane was blocked in Tris-buffered saline (TBS) containing 5% bovine serum albumin (MP Biomedical, Solon, OH, USA) for 2 h at room temperature, then incubated with primary antibodies overnight at 4 °C. The following primary antibodies were used: SET8 (1:1,000, catalog No 14063-1-AP, Proteintech, Rosemont, IL, USA); RNF8 (1:1,000, catalog No. 14112-1-AP); H4K20me1 (1:1,000, catalog No. 9724; CST, Danvers, MA, USA); and GAPDH (1:2,000, catalog No. TA-08; ZSGB-BIO, Beijing, China). After twice-washing with TBS, the membranes were incubated with the appropriate horseradish peroxidase (HRP)-labeled secondary antibody for 1 h at room temperature. Secondary antibodies conjugated with HRP included goat anti-rabbit IgG (1:5,000, catalog No. ZB-2301; ZSGB-BIO). Immunoblots were visualized using an enhanced chemiluminescence (ECL) detection system (Thermal Fisher Scientific) according to the manufacturer’s protocol, and autoradiography.
-
The animal experiments conducted in this study were approved by the Wenzhou Medical University Institutional Animal Use and Care Committee. C57BL/6J mice were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (Beijing, China). Prior to tumor cell injection, the right hindlimbs of age-matched 6-week-old female mice were shaved. LLC1 cells (2 × 105) were resuspended in 50 µL of PBS and injected subcutaneously into the shaved flanks with vector control or SET8 knockout tumor cells. After 7 days, tumors were X-irradiated using lead shielding with a single 8 Gy dose. Tumor volumes were measured every other day and calculated using the following formula: (length) x (width)2/2. The mice were sacrificed when tumor volumes reached 2,000 mm3. The Kaplan-Meier estimator and log-rank (Mantel-Cox) test were used for survival analysis among different tumor-bearing mice groups.
-
After irradiation, cells were washed with PBS buffer, trypsinized, and counted using a cell counter (Beckman, Indianapolis, IN, USA). An appropriate number of cells (0 Gy: 100; 1 Gy: 200; 2 Gy: 500; 4 Gy: 2,000; 6 Gy: 10,000) were seeded into each Φ60 dish in 5 mL of complete media. After 10 days of incubation, colonies were fixed with 10 mL of fresh Carnoy’s fluid and stained with 0.5% crystal violet for 20 min[14]. Colonies with > 50 remaining cells were recorded and counted manually under an inverted microscope. Plating efficiencies (PEs) were calculated as follows: number of colonies formed/number of cells plated. Survival ratios were calculated as follows: numbers of colonies formed/numbers of cells plated. Cell surviving fractions were calculated as follows: PE (irradiated)/PE (non-irradiated).
-
The micronucleus test is used in toxicologic screening for potential genotoxic targets[15]. Forty-eight hours after 4 Gy X-irradiation, cells were fixed with Carnoy’s fluid for 20 min at room temperature and stained with 20 μL of acridine orange in an aqueous solution (10 μg/mL). Micronuclei identification was established by the following criteria: diameter less than one-third of the primary nucleus; and color is the same as or lighter than the main nucleus[16]. Analyses were performed with an Axio Imager Z2 fluorescence microscope (Zeiss, Oberkochen, Germany) at 20 × magnification. At least 500 cells were scored for each sample.
-
Apoptosis was evaluated using an Annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) Apoptosis Detection Kit (Solarbio, Beijing, China) according to the manufacturer’s instructions. Forty-eight hours after 4 Gy X-irradiation, the culture medium was removed and the wells were washed with PBS, then the cells were harvested by trypsinization and resuspended in binding buffer at 1 × 106 cells/mL and stained with Annexin V-FITC and PI on ice for 15 min in the dark. Cell fluorescence was examined with an ImageSteam100 (Amnis, Austin, TX, USA).
-
Single-cell RNA sequencing analyses were performed using the CancerSEA database (http://biocc.hrbmu.edu.cn/CancerSEA), which is a cancer single-cell functional state atlas, involving 14 functional states of 41,900 single cancer cells from 25 cancer types[17]. The scores of 14 functional states, including DNA repaired across single cancer cells in each dataset, were evaluated using gene set variation analysis (GSVA). The cohorts analyzed include LUAD and BRCA single-cell RNA sequencing data, which were retrieved from published studies[18–21]. A non-parametric t-test was used to determine the statistical differences among different single-cell groups.
-
Extraction of total RNA from cells was performed using the E.Z.N.A.® Total RNA Kit (Omega, Norcross, GA, USA) following the manufacturer’s protocol. Reverse transcription was performed with the Transcriptor First Strand cDNA Synthesis Kit (Roche) and qRT-PCR was performed using SYBR Green PCR Master (Roche) to measure mRNA expression. The SET8 and GAPDH (internal control) primers were purchased from GeneCopoeia. The primers used for different genes are listed in Supplementary Table S2, available in www.besjournal.com. qRT-PCR was performed on samples using the Chromo4 System Real-Time PCR detector (Bio-Rad, USA). All procedures were performed according to the manufacturers’ protocols under the following conditions: initiation for 10 min at 95 °C; and 40 thermal cycles each at 95 °C for 10 s, 60 °C for 20 s, and 70 °C for 10 s. Relative fold-change in mRNA expression was calculated using the 2−ΔΔCt method with the following equation: relative quantitation (RQ) = 2-ΔΔCt.
Table S2. Oligonucleotide primers for qRT-PCR
Genes Upstream primer (5'–3') Downstream primer (5'–3') SET8 CGCAAACTTACGGATTTCT CGATGAGGTCAATCTTCATT
GAPDH
GTCTCCTCTGACTTCAACAGCG
ACCACCCTGTTGCTGTAGCCAA -
To detect 53BP1 and γ-H2AX foci that formed at the DSB sites and the level of RNF8 expression, cells were seeded at 1 × 104 on glass coverslips in each well of a 12-well tissue culture plate and cultured for 24 h prior to being irradiated according to the experimental requirements. Subsequent experimental procedures were ass previously described[22]. After 1 Gy X-irradiation, cells were fixed with 4% paraformaldehyde (PFA) for 20 min at room temperature and permeabilized with 0.5% Triton X-100 in PBS while on ice for 10 min. Non-specific binding sites were blocked with 5% bovine serum albumin (BSA) in PBS for 60 min at room temperature prior to probing with primary antibodies. The primary antibodies used for immunostaining included RNF8 (1:500), 53BP1 (1:3,000, catalog No. ab36823; Abcam, Cambridge, MA, USA), and γ-H2AX (1:1,000, catalog No. 80312; CST). Secondary antibodies (anti-mouse or rabbit conjugated with Alexa 488/647) were purchased from Beyotime (1:2,000, catalog Nos. A0423 and A0473, respectively). Then, the cells were incubated in the primary antibody diluted in 5% BSA/1 × PBS 1 h at room temperature. After incubation, cells were thrice-washed with PBS for 10 min each, then incubated with the appropriate Alexa Fluor secondary antibodies diluted in 5% BSA for 1 h. The cells were thrice-washed with PBS again for 10 min each. The nuclei were counterstained with DAPI (Beyotime). Digital image analysis was performed to determine the fluorescence intensity of RNF8 and the number of γH2AX and 53BP1 foci using an Axio Imager Z2 fluorescence microscope at 63× magnification and confirmed by visual inspection of images. Quantification of fluorescence intensity or foci per cell was determined from images of 50 cells for every time point from at least three independent experiments.
-
For RNA sequencing, samples were collected, snap-frozen, and maintained at –80 °C prior to RNA extraction. Total RNA was extracted with TRIzol reagent (Invitrogen). Sequencing was performed using the Illumina sequencing platform (Majorbio Co., Shanghai, China). The measured data were mapped to reference genomes after quality control. After counting the reads mapped on the gene and calculating the gene expression, the differences in gene expression were analyzed.
-
All in vitro experiments were performed in triplicate and repeated at least three times. Statistical significance (P values) of differences in means between two samples were evaluated using unpaired t-tests. Data are represented as individual values or as the mean ± sem. Group sizes (n) and applied statistical tests are indicated in the figure legends. For in vivo experiments and clinic data analysis, log-rank (Mantel-Cox) tests were used for survival analysis and tumor volume significances were assessed by two-way ANOVA among different groups. Statistics were calculated using GraphPad Prism 8.2.1. A P < 0.05 was considered statistically significant (*), P < 0.01 as highly significant (**), and P < 0.001 (***) as extremely significant.
-
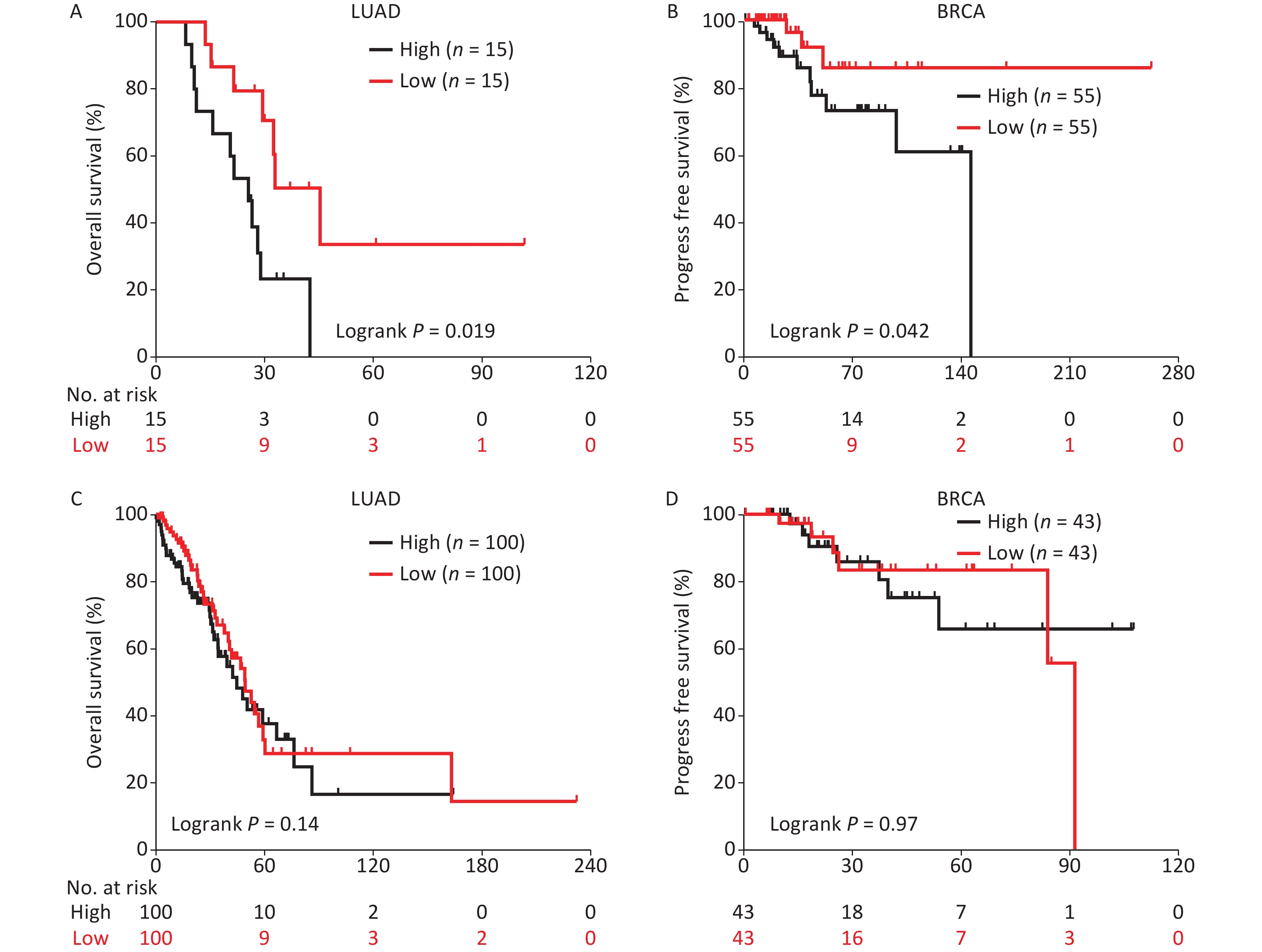
To determine the potential association between SET8 and radiotherapy in patients with cancer, we analyzed SET8 expression and efficacy of radiotherapy in LUAD and BRCA cancer patients known to have undergone radiotherapy[11]. Our analysis indicated that LUAD patients with low SET8 expression had a significant advantage in overall survival (OS) when compared with high SET8 expression. Specifically, the median duration of OS was 45 and 26 months in the low and high SET8 expression groups, respectively (P = 0.018, log-rank test; Figure 1A). BRCA patients with low SET8 expression had a significantly better probability of progress-free survival (PFS) than the high-expression group; specifically, the median duration of PFS in the low-expression group was undefined compared to 146 months in the high-expression group (P = 0.042, Figure 1B). Importantly, the OS in the LUAD patients without radiotherapy was not significantly different between the SET8 low- and high-expression groups (P = 0.41, Figure 1C). Similarly, the PFS of BRCA patients without radiotherapy was not significantly different between the SET8 low- and high-expression groups (P = 0.97, Figure 1D).

Figure 1. Kaplan-Meier survival curves of radiotherapy treated LUAD and BRCA patients with different SET8 expression. (A) Kaplan-Meier survival curves of radiotherapy-treated LUAD patients with low or high SET8 expression. (B) Kaplan-Meier survival curves of radiotherapy-treated BRCA patients with low or high SET8 expression. (C) Kaplan-Meier survival curves of radiotherapy untreated LUAD patients with low or high SET8 expression. (D) Kaplan-Meier survival curves of radiotherapy untreated BRCA patients with low or high SET8 expression.The cut-off values of high and low are top 25% and lowest 25% in LUAD, top 10% and lowest 10% in BRCA. P-values calculated by use of logrank test.
Therefore, these data clearly demonstrated that low SET8 expression has significant advantages in cancer radiotherapy and it is directly responsible for the observed prognostic benefit in radiotherapy-treated patients among the TCGA PanCancer cohorts.
-
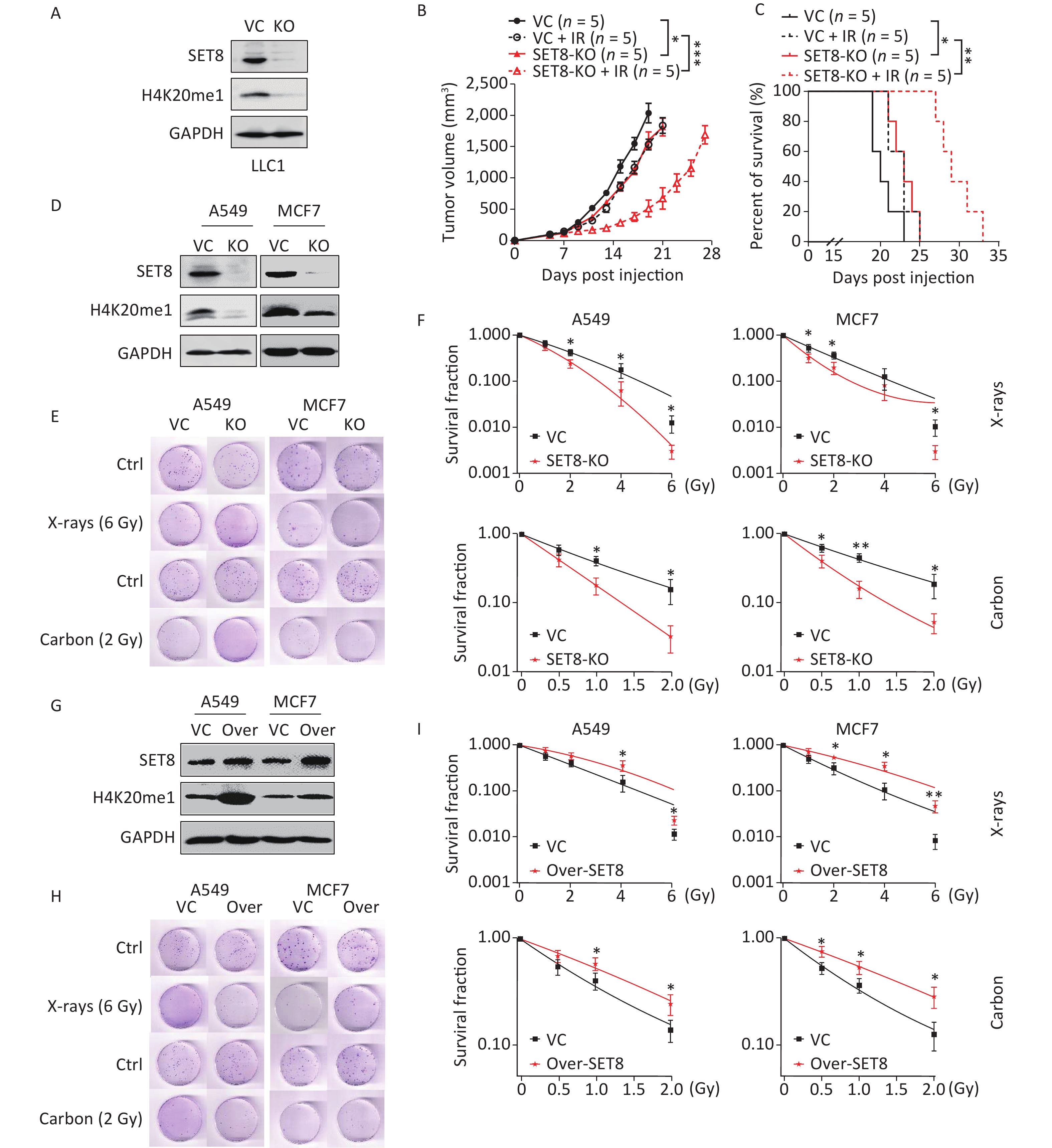
In consideration of low SET8 expression benefiting radiotherapy, we hypothesized that SET8 deficiency sensitizes tumor cells to radiation. To confirm our hypothesis, we generated SET8 knockout LLC1 tumor cells using CRISPR/Cas9 (Figure 2A). Vector control and SET8 KO LLC1 cells were then used to establish subcutaneous tumors, which were then treated with 8 Gy of X-ray radiotherapy after 7 d. Our data indicated that SET8-deficient tumors grew slower than controls. Furthermore, SET8-deficient tumors responded to radiotherapy significantly better than vector controls (Figure 2B). Mice bearing SET8 KO tumors also survived significantly longer after radiotherapy (Figure 2C).
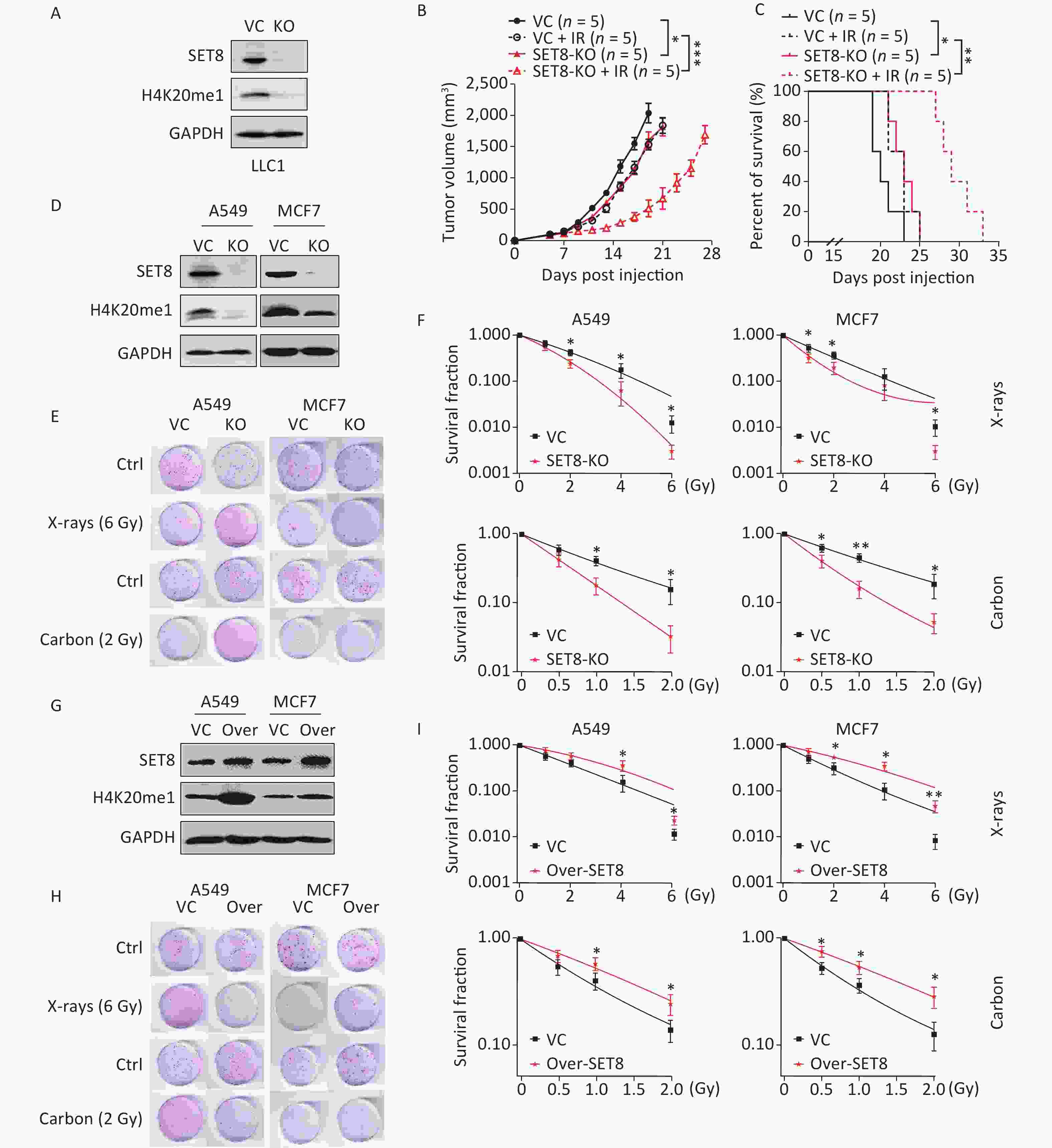
Figure 2. Radiosensitivity in the SET8 alteration A549 and MCF7 cells. (A) Western blot analysis of SET8 levels in vector control (VC) and SET8 KO groups B16F10 cells. (B) & (C) Tumor growth delay and Kaplan-Meier survival curves of C57BL/6 mice inoculated with 1 x 105 control or SE8-KO LLC1 cells. Radiotherapy was conducted 1 x at 8 Gy at 7 days post inoculation of the tumor cells. (D) Western blot assay of SET8 and H4K20me1 protein levels in SET8-KO or control A549 and MCF7 cells. (E) & (F) Colony formation assay of SET8-KO and control A549 and MCF7 cells were exposed to X rays or Carbon. (G) Western blot assay of SET8 and H4K20me1 protein level in SET8 overexpression (Over-SET8) and control A549 and MCF7 cells. (H) & (I) Colony formation assay of Over-SET8 and control A549 and MCF7 cells were exposed to X rays or Carbon. Significance was determined by 2-way ANOVA in (B), logrank test in c and unpaired t-test in (F) and (H) respectively. *P < 0.05; **P < 0.01, ***P < 0.001.
To evaluate the influence of SET8 on the intrinsic radiosensitivity of cancer cells, we generated SET8 knockout A549 and MCF7 cells using CRISPR/Cas9 (Figure 2D). We then performed a colony-forming assay after exposing the cells to different doses of X-rays. Our results indicated that SET8 deficiency significantly decreased the clonogenic abilities of A549 and MCF7 cells, especially at higher radiation doses (Figure 2E–F). Furthermore, the survival fraction increased significantly in the irradiated SET8 overexpression group compared with controls (Figure 2G–I).
We also measured the levels of SET8 and H4K20me1 expression in A549 cells exposed to low LET (X rays) and high LET (carbon ions, a promising radiotherapy particle) radiation at a series of time points using qRT-PCR and Western blotting, respectively (extended data, Supplementary Figure S1, available in www.besjournal.com). Our results showed that SET8 and H4K20me1 expression decreased from 4–48 h in carbon ion X-irradiated A549 cells.

Figure S1. SET8 expressions and H4K20me1 dynamic changes after X-ray or carbon ion radiation. (A) Relative SET8 mRNA expression was measured by qRT-PCR at indicated time points in A549 cells after 4 Gy X-ray irradiation. GAPDH was used as internal controls. (B) SET8 and H4K20me1 levels in A549 cells at indicated time points after 4 Gy X-ray radiation were measured by western blot assay. (C) Relative SET8 mRNA expression was measured by qRT-PCR at indicated time points in A549 cells after 2 Gy Carbon irradiation. (D) SET8 and H4K20me1 levels in A549 cells at indicated time points after 2 Gy Carbon radiation were measured by western blot assay. Significance was determined by unpaired t-test. *P < 0.05; **P < 0.01.
Taken together, these results indicate that SET8 expression alteration affected radiotherapy in vivo and in vitro and responded to ionizing radiation.
-
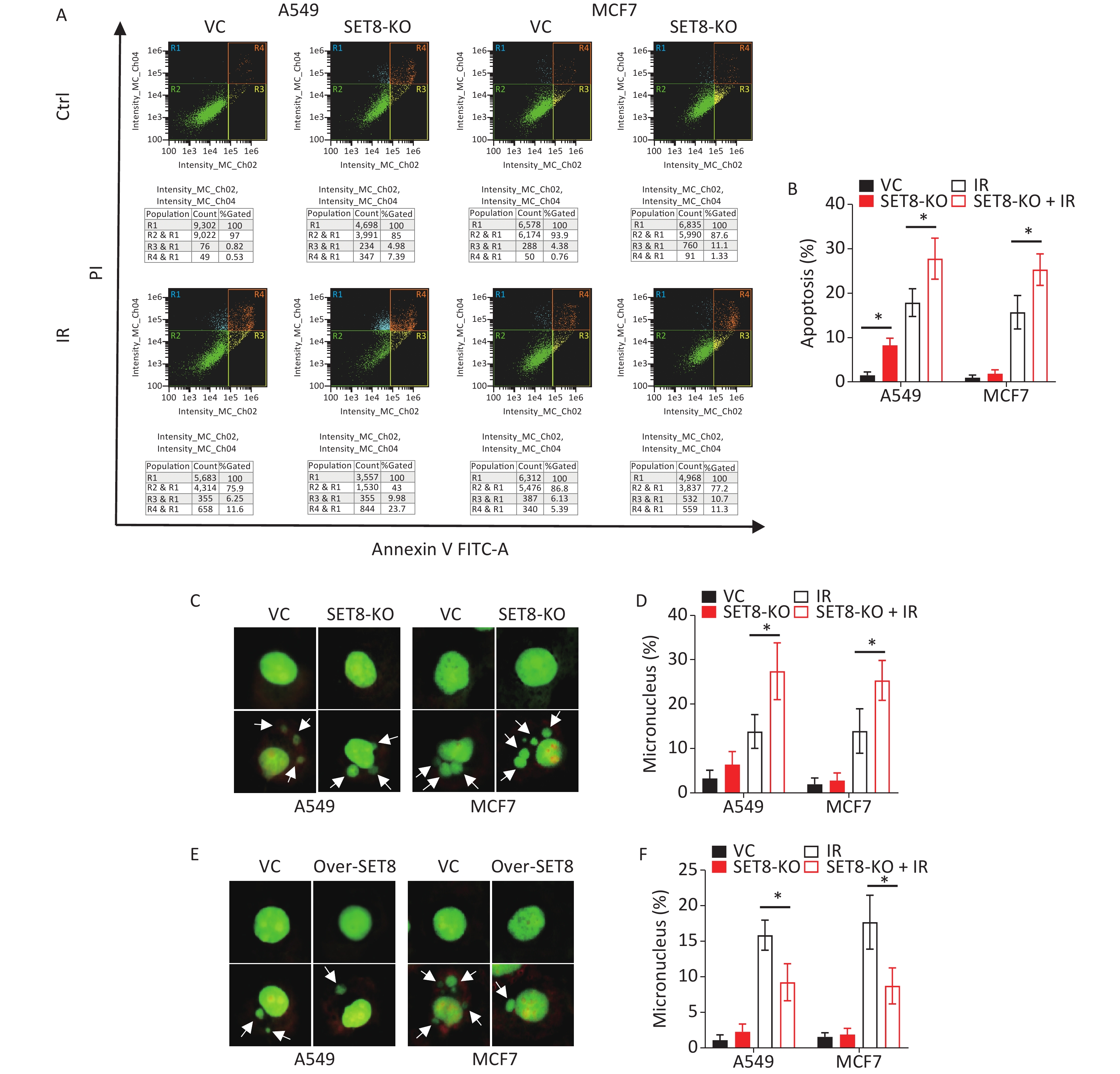
Next, we attempted to determine if SET8 expression regulates radiation-induced cellular apoptosis and nuclear injury. Figure 3A–B show that SET8 knockout increased the proportion of radiation-induced apoptosis in A549 and MCF7 cells. Furthermore, SET8 knockout increased the frequency of micronuclei in irradiated cells (Figure 3C–D). Conversely, SET8 overexpression decreased the frequency of micronuclei (Figure 3E–F). These results further indicate that SET8 is positively correlated with radiation resistance.

Figure 3. Cellular apoptosis and nucleus injury in irradiated SET8 alteration A549 and MCF7 cells. (A) & (B) Apoptosis proportion of SET8-KO or control A549 and MCF7 cells were exposed to 4 Gy X rays 48 h later. (C) & (D) Micronucleus frequency of SET8-KO or control A549 and MCF7 cells were exposed to 4 Gy X rays 48 h later. (E) & (F) Micronucleus frequency of Over-SET8 and control A549 and MCF7 cells were exposed to 4 Gy X rays 48 h later. Significance was determined by unpaired t-test. *P < 0.05. IR, irradiation. VC, vector control.
-
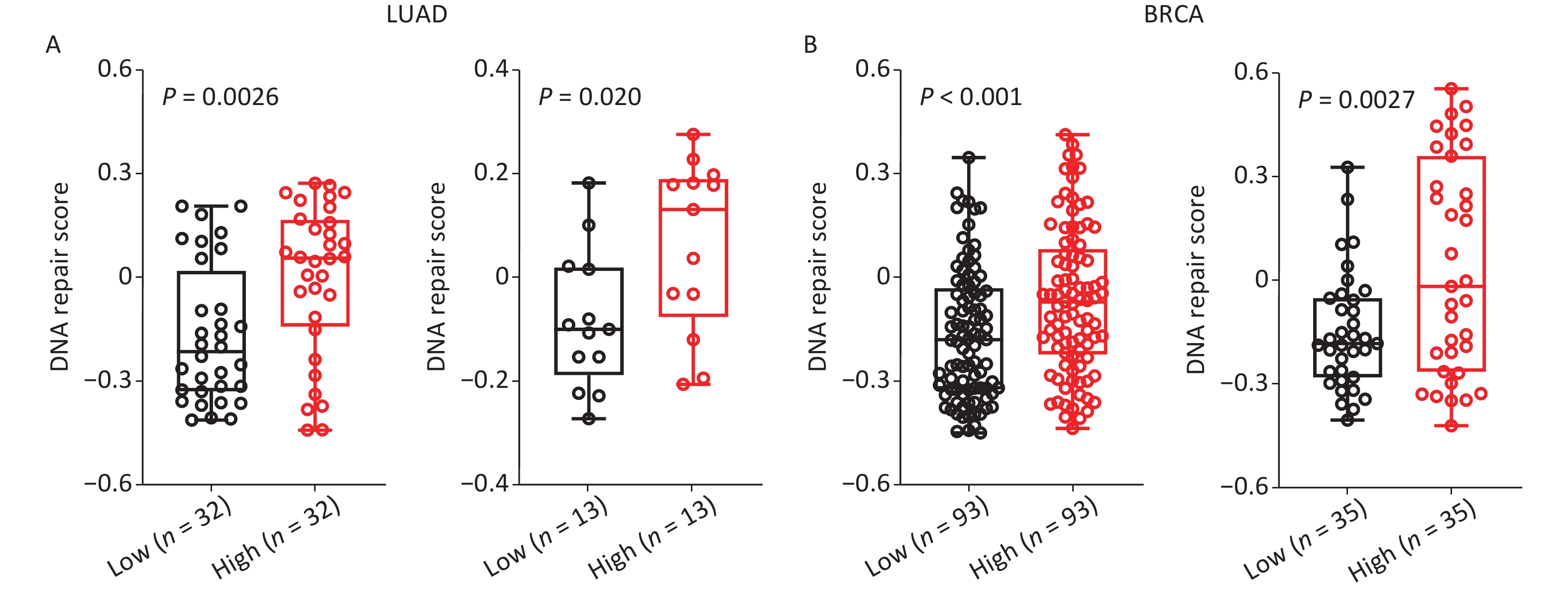
Further analysis of the role of SET8 in DNA repair was carried out utilizing ScRNA-seq to identify the mechanism underlying the SET8 increase in radiotherapy efficacy. The scores of DNA repaired across single cancer cells in each dataset were evaluated using the CancerSEA database. Comparing the SET8 low- and high-expression groups, there were significant positive correlations between SET8 expression and DNA repair scores in the two independent LUAD cohorts, including 126 and 42 samples, respectively (Figure 4A)[18,19]. Similar results were obtained in the two independent BRCA cohorts that included 369 and 70 samples (Figure 4B)[20,21]. Thus, the above analyses proved that there is a positive association between SET8 expression and DNA repair.
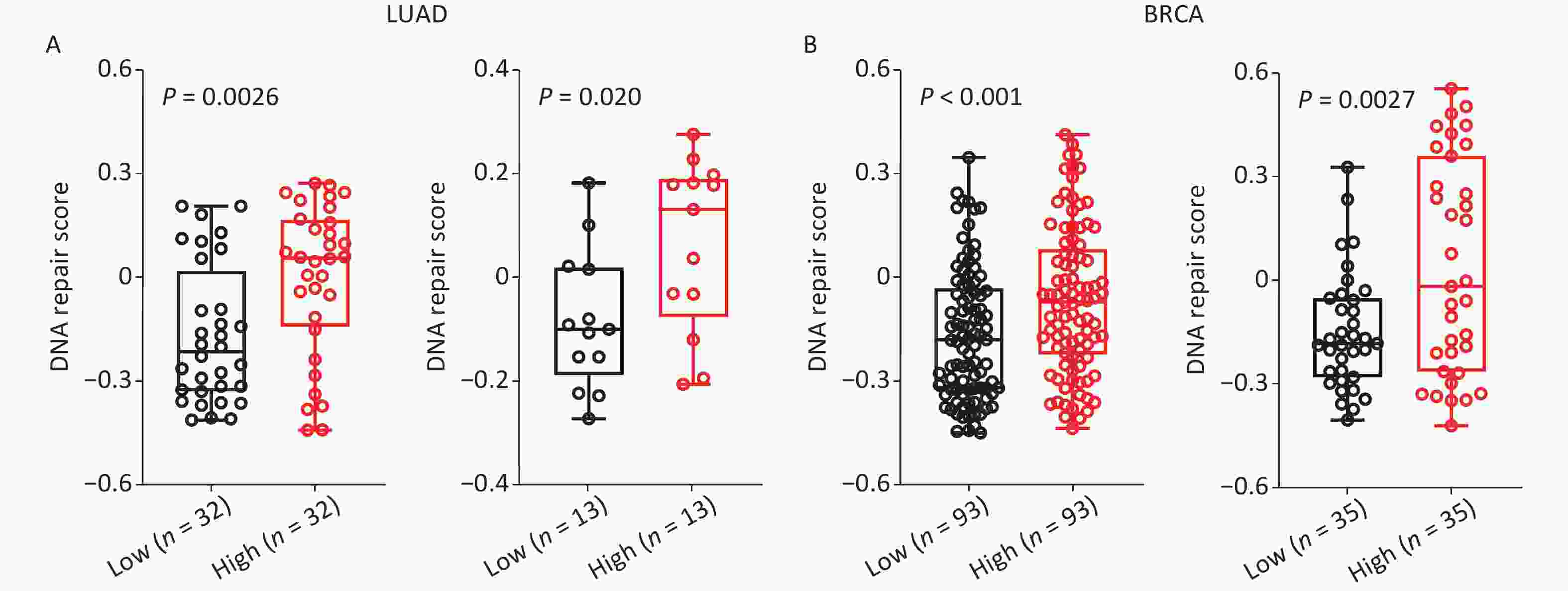
Figure 4. Single-cell RNA sequencing analyses between SET8 expression and DNA repair scores in LUAD and BRCA. (A) Scores of DNA repaired through single-cell RNA sequencing analyses in LUAD with SET8 expression low or high signature. (B) Scores of DNA repaired through single-cell RNA sequencing analyses in BRCA with SET8 expression low or high signature. The cut-off values of high and low are top 25% and lowest 25% in LUAD and BRCA . P-values calculated by use of unpaired t-test .
-
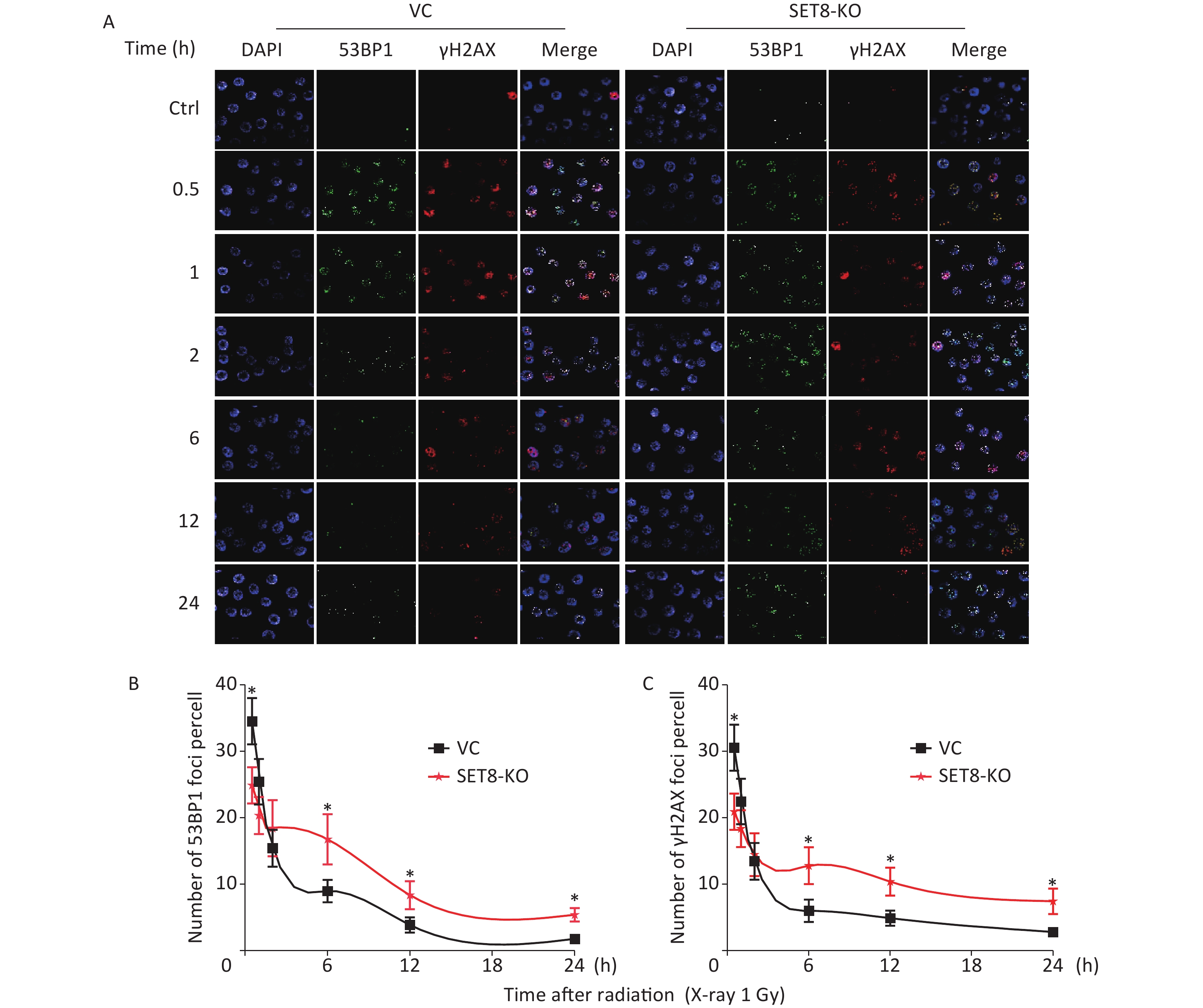
Because we have determined that SET8 expression is associated with DNA repair by ScRNA-seq, we subsequently speculated that SET8 inhibition enhanced radiotherapy by blocking the DNA damage repair process. To confirm this assumption, we directly visualized 53BP1 and γH2AX foci, which are surrogate markers for DNA damage repair, by immunofluorescence. First, significantly lower numbers of 53BP1 and γH2AX foci were observed 0.5 h after 1 Gy X-irradiation in SET8-deficient A549 cells (Figure 5). Further, the kinetics of foci dissolution of both two markers were delayed in SET8-deficient cells than in SET8-negative cells from 6–24 h. The above results indicated that SET8 deficiency hindered the recruitment of the DNA repair complex and DNA damage was refractory to repair in SET8-deficient A549 cells, which is consistent with previous reports[6,7]. The above evidence suggests that SET8 deficiency blocked the kinetics process of DNA damage repair.
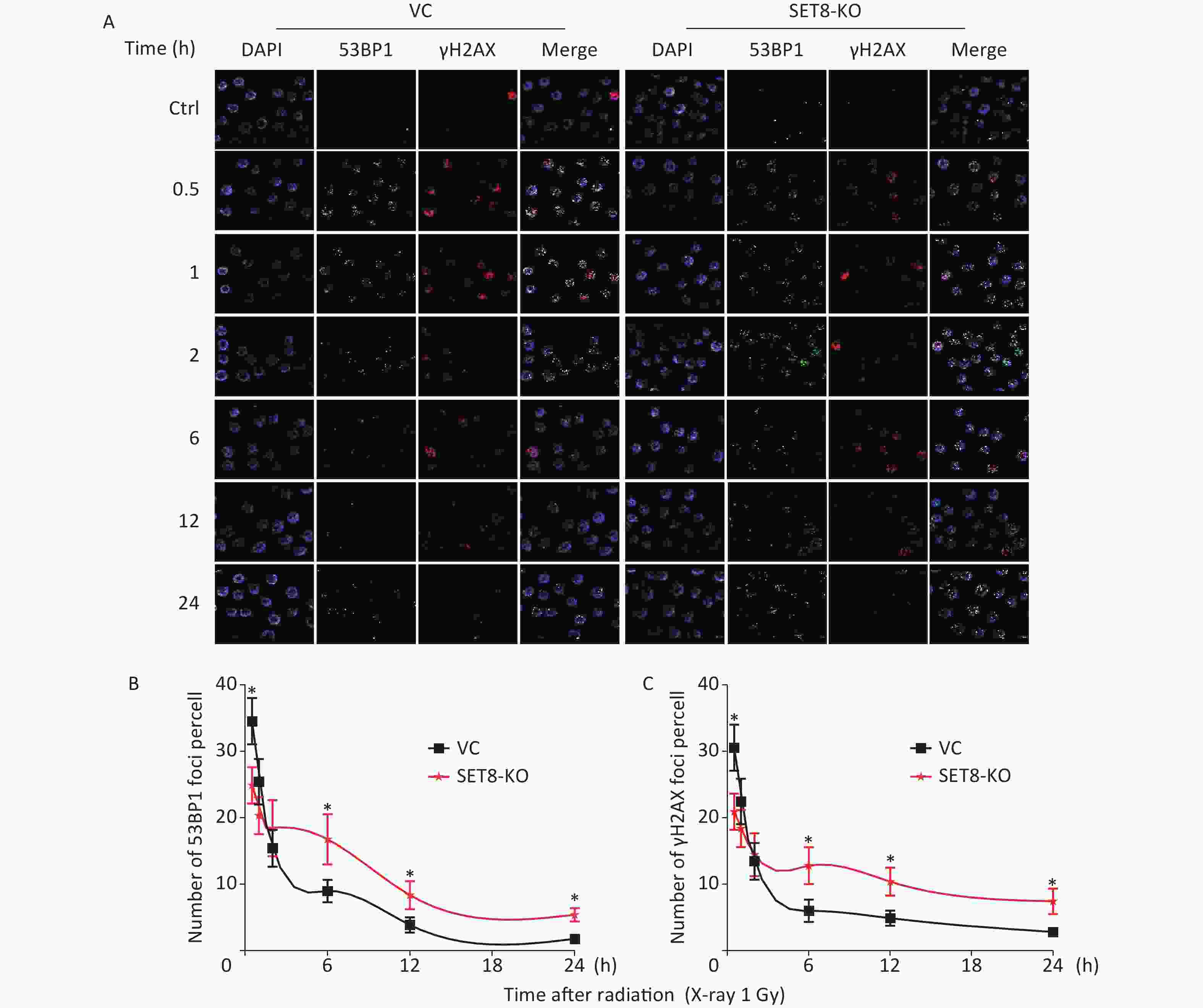
Figure 5. Kinetics process of DNA damage repair in SET8 knockdown A549 cells. (A) Graph shows quantification of 53BP1 and γH2AX foci in SET8-KO or control A549 cells exposed to 1 Gy X rays. (B) The numbers of 53BP1 foci in 50 cells of each group were counted for each time point. (C) The numbers of γH2AX foci in 50 cells of each group were counted for each time point. Significance was determined by unpaired t-test . *P < 0.05. VC, vector control.
-
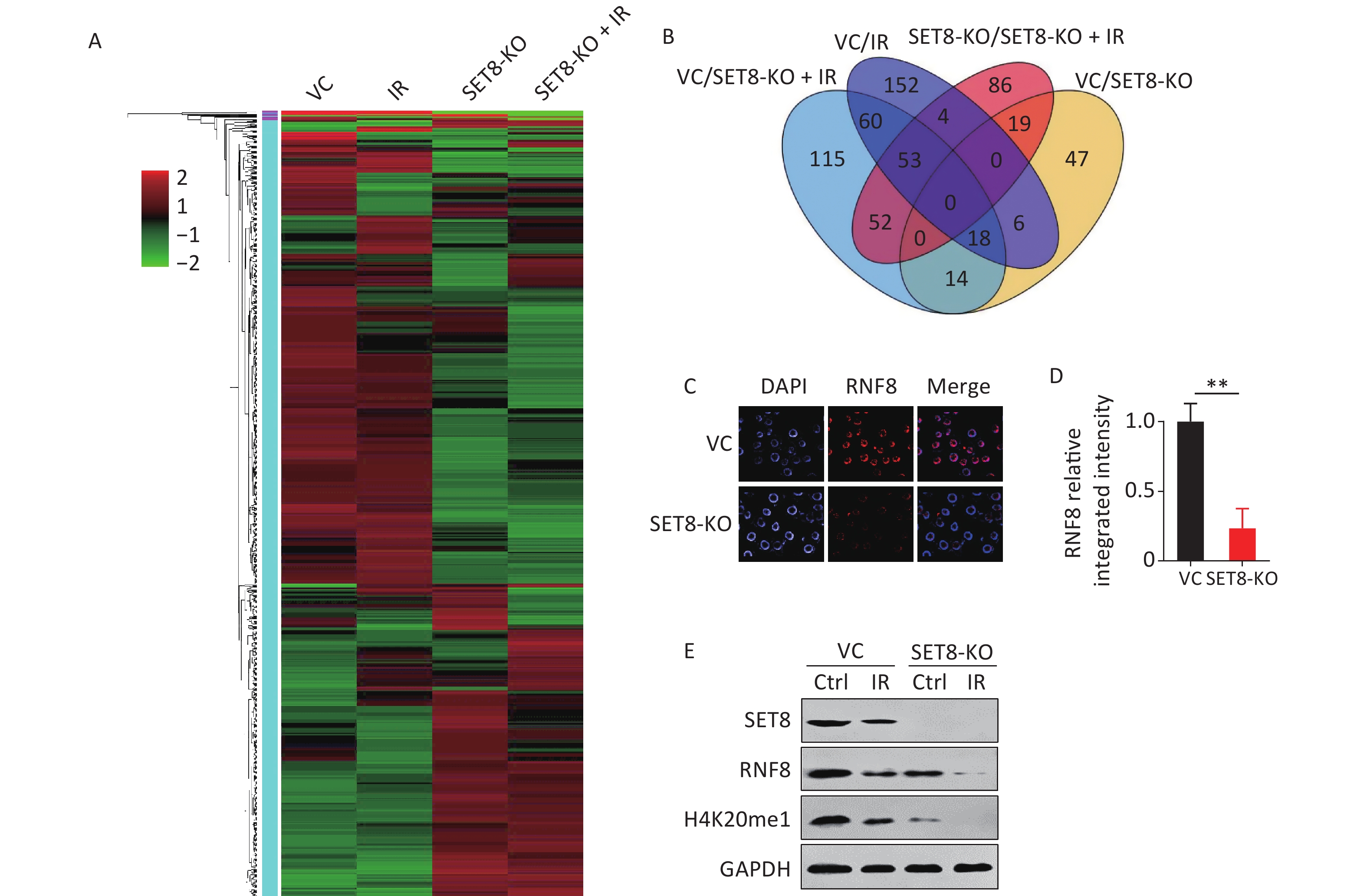
We further investigated the functional connection between SET8 and other underlying genes to determine the mechanism underlying SET8-regulated radiotherapy efficacy. Before RNA sequencing, SET8 knockout and control A549 cells were exposed to 0 and 4 Gy of X-rays and collected 12 h after irradiation. Duplicate samples were in each group for subsequent investigation. Gene expression relative fold-changes were calculated via RNA sequencing data. The set of 312 genes with > 2 fold-changes was analyzed using hierarchical clustering heatmap to reveal expression signatures (Figure 6A). Among the 312 genes, expression of 104 were significantly different between the SET8 knockout and control groups (43 were upregulated and 61 were downregulated; Figure 6B, Supplementary Table S3, available in www.besjournal.com). Ring finger protein 8 (RNF8) is involved in the aforementioned genes. RNF8 is an E3 ubiquitin (Ub) ligase with RING finger and has key roles in DNA damage and repair[23]. Meanwhile, Zhou et al.[22] reported that silencing of RNF8 enhances radiosensitivity in A549. Because RNF8 had persistent low expression in the SET8-KO groups that were independent of radiation by RNA sequencing, we explored the correlation between SET8 and RNF8. Figure 6C and d showed that the relative fluorescence intensity was low in the SET8 inhibition group. Our data further confirmed that RNF8 was downregulated in SET8 inhibition cells (Figure 6E). Above all, these results implied that SET8 inhibition induced radiation sensitization that depended on impeding RNF8 expression.
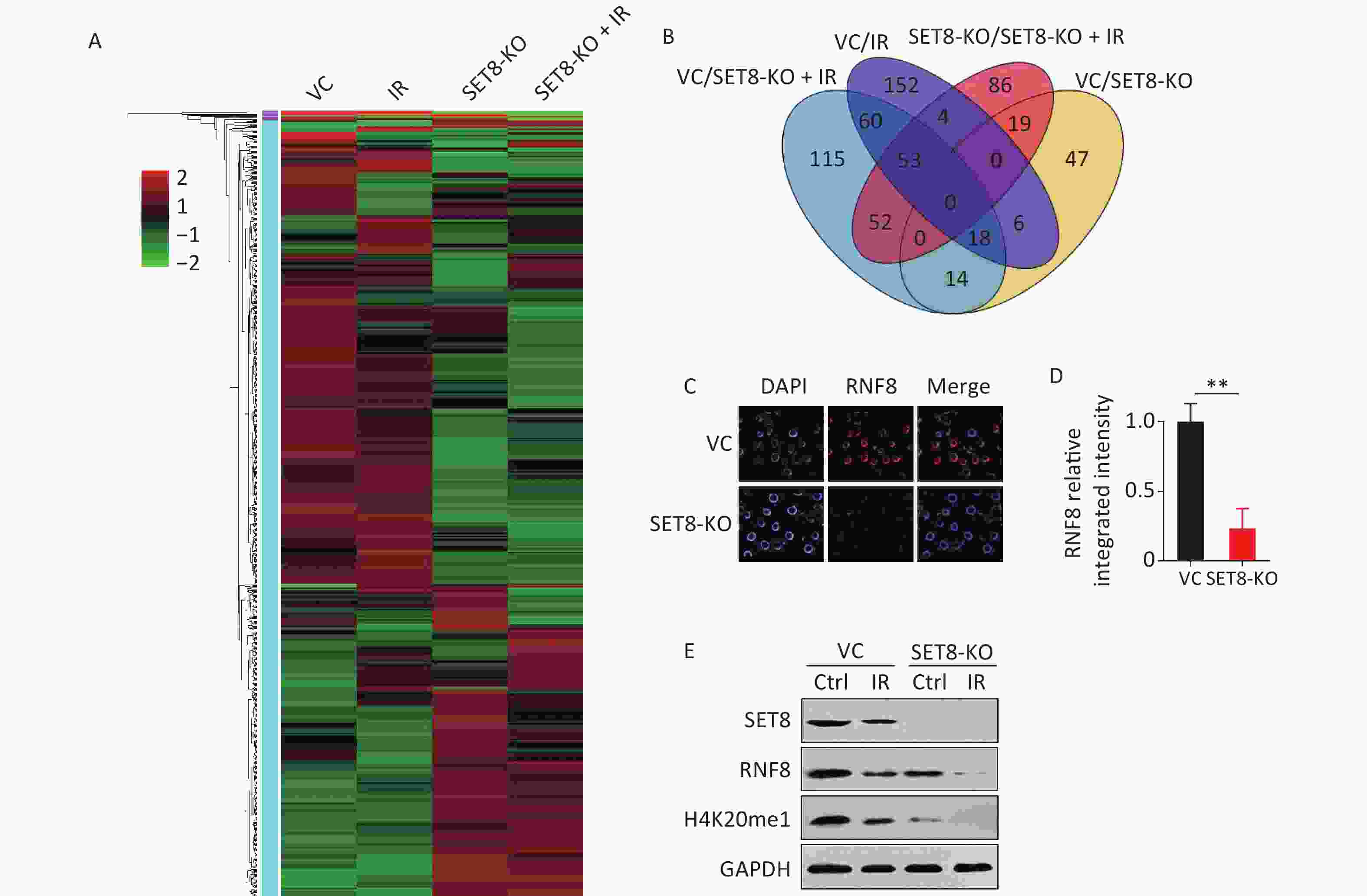
Figure 6. Radiosensitivity in the SET8 alteration A549 and MCF7 cells. (A) Heat map of RNA sequnece analysis in vector control (VC) and SET8 KO groups A549 cells, 12 h after 4Gy X-ray radiation. Genes that were significantly differentially expressed (P < 0.05) and had greater than 2-fold difference in expression were identified. (B) Venn diagram showing the total number of gene comparisons between different treatments groups. (C) & (D) Representative immunofluorescence images and RNF8 relative integrated immunofluorescence intensity in control and SET8-KO A549 cells. (E) Western blot assay of SET8, RNF8 and H4K20me1 protein levels in SET8-KO or control A549 and MCF7 cells exposed to 4 Gy X rays after 8 h. Significance was determined by unpaired t-test in (D). **P < 0.01. IR, irradiation
-
SET8 has been implicated in many essential cellular processes, including cell cycle regulation, DNA replication, and the DNA damage response.[7] There is no conclusive evidence about the contribution of SET8 to radiation treatment. The cBioPortal database provided abundant clinical data from TCGA, which provided the most direct evidence between genomic data and cancer prognosis in a variety of therapeutic strategies. Meanwhile, the genome-wide expression profiling of cancer tissues is an extremely useful method to explore the correlation of the genes in vivo[11,12]. Our clinical and genomic data analysis in radiation-treated LUAD and BRCA patients offered theoretical proof that SET8 deficiency contributed to better radiotherapy efficiency. Subsequently, the current experiments clearly demonstrated that genetic deletion of SET8 significantly enhanced radiotherapy in a murine tumor model, and A549 and MCF7 cells in vitro, and SET8 overexpression was effective in decreasing radiosensitivity, which suggests the benefit of low SET8 expression to radiotherapy.
Emerging pieces of evidence have revealed that radiation-induced alterations in histone methylation affect the cellular response to radiation damage in the DNA[24–29]. It is therefore important to characterize radiation-induced alterations in histone methylation patterns and the associated factors. Our study showed that ionizing radiation induced the recession of SET8 and H4K20me1 in the short term after irradiation. SET8/H4K20me1 oscillates during the cell cycle, and maintenance of H4K20me1 levels is critical for proper cell cycle progression through its role in protecting genome stability, DNA replication, mitosis, and transcription[7], which indicates that SET8/H4K20me1 may have an identical function in low and high LET radiation-induced DNA damage. SET8 directly modulates chromatin compaction and may be involved in the recruitment of signaling proteins, such as 53BP1, to the site of double-strand DNA breaks (DSBs)[30,31]. Furthermore, when chromatin reorganization at DSBs is blocked, DNA repair is inhibited and cells exhibit increased sensitivity to radiation that creates DSBs[32]. Our subsequent experiments revealed that SET8/H4K20me1 also affected radiosensitivity by blocking the kinetics process of DNA damage repair based on the reduction in 53BP1 and γ-H2AX foci.
RNF8 plays a key role in cellular life processes that promote the recruitment of several proteins, such as 53BP1, Rad51, and BRCA1, at the sites of DNA damage by regulating the ubiquitination of H2AX to facilitate DNA repair. Previous studies have reported that aberrantly-expressed RNF8 may disrupt the DNA damage repair process[33,34], especially in the A549 cells[35]. Combined with our results, SET8 inhibition induced radiation sensitization involving the RNF8 pathway.
A series of inhibitors of SET8 or H4K20me1 have been synthesized and exhibit crucial biological functions in cell proliferation and DNA repair[36–38]. SPS8I1–3 are small molecule inhibitors of SET8 that can result in cell-cycle defects at the S and G2/M phases, and the phenomenon can be recapitulated by RNAi-mediated knockdown of SET8[36]. The H4K20me1 inhibitor, A-196, inhibits 53BP1 foci formation upon ionizing radiation and reduces NHEJ-mediated DNA repair[37]. In addition, the inhibitor of SET8 (UNC-0379) blocks H4K20 methylation and augments radiosensitivity by reducing recruitment of the 53BP1 protein to DSBs is an effective radiosensitizer of human glioma cells[38]. Developing SET8/H4K20me1 inhibitors is an effective step toward elucidating the roles of SET8 in DNA repair and appears to be a promising prospect on combination radiotherapy via convenient pharmacologic perturbation.
In conclusion, our work and existing reports suggest that the histone methylation pathway, SET8/H4K20me1, which is required for DNA repair, is an important and novel target for the development of radiosensitizers with therapeutic relevance. As new epigenetic inhibitors are synthesized and tested in preclinical and clinical settings, it will be important to consider combining the epigenetic inhibitors with radiation or other types of DNA-damaging chemotherapy for precise radiotherapy.
-
BH and DP contributed to the conception and design of the study. DP, RD, RL, and AS conducted the investigation. DP and BH wrote the first draft of the manuscript. RD, XL, and CL wrote sections and reviewed the manuscript. All authors contributed to the manuscript revision, and read and approved the submitted version.
-
We also appreciate the staff of the Heavy Ion Research Facility in Lanzhou for the offer of 12C6+ ion.
Table S3. Gene expression signatures in SET8-KO A549 cells
Seq_id (VC) fpkm (KO)3_7_fpkm log2FC(KO/VC) Regulate Description ENSG00000264148 0 49.1764 5.61989422 up − ENSG00000236991 0.330355 3.76492 3.510530124 up − ENSG00000088448 41.5497 364.193 3.131793228 up ankyrin repeat domain 10 [Source:HGNC Symbol;Acc:20265] ENSG00000185437 0 8.08694 3.015593908 up SH3 domain binding glutamic acid-rich protein [Source:HGNC Symbol;Acc:10822] ENSG00000228950 0 7.66067 2.937470575 up − ENSG00000271267 0 5.72202 2.51652454 up − ENSG00000232859 1.14869 6.19551 2.43123354 up LYR motif containing 9 [Source:HGNC Symbol;Acc:27314] ENSG00000172339 1.08544 5.63913 2.377192622 up asparagine-linked glycosylation 14 homolog (S. cerevisiae) [Source:HGNC Symbol;Acc:28287] ENSG00000213420 1.27423 5.78869 2.183611189 up glypican 2 [Source:HGNC Symbol;Acc:4450] ENSG00000131095 0.961888 4.15931 2.112403391 up glial fibrillary acidic protein [Source:HGNC Symbol;Acc:4235] ENSG00000185920 3.25398 13.6198 2.065428229 up patched 1 [Source:HGNC Symbol;Acc:9585] ENSG00000234160 0 3.97046 1.989306161 up − ENSG00000241769 1.82417 7.01286 1.942762742 up − ENSG00000153291 1.65069 6.24826 1.92038528 up solute carrier family 25, member 27 [Source:HGNC Symbol;Acc:21065] ENSG00000254335 0 3.74532 1.905088988 up cadherin 12 (N-cadherin 2) pseudogene 1 [Source:HGNC Symbol;Acc:37698] ENSG00000230438 1.00366 3.68563 1.876640622 up − ENSG00000170899 1.78022 5.97155 1.746049911 up glutathione S-transferase alpha 4 [Source:HGNC Symbol;Acc:4629] ENSG00000104205 1.80144 5.94424 1.722341765 up serum/glucocorticoid regulated kinase family, member 3 [Source:HGNC Symbol;Acc:10812] ENSG00000139946 1.0563 3.48076 1.72038271 up pellino E3 ubiquitin protein ligase family member 2 [Source:HGNC Symbol;Acc:8828] ENSG00000137843 1.61001 5.23319 1.70062099 up p21 protein (Cdc42/Rac)-activated kinase 6 [Source:HGNC Symbol;Acc:16061] ENSG00000246273 6.43651 18.854 1.550520085 up SBF2 antisense RNA 1 [Source:HGNC Symbol;Acc:27438] ENSG00000176371 3.07709 8.49372 1.464829909 up zinc finger and SCAN domain containing 2 [Source:HGNC Symbol;Acc:20994] ENSG00000080298 1.3528 3.72538 1.461439027 up regulatory factor X, 3 (influences HLA class II expression) [Source:HGNC Symbol;Acc:9984] ENSG00000133794 5.24692 13.2361 1.334935398 up aryl hydrocarbon receptor nuclear translocator-like [Source:HGNC Symbol;Acc:701] ENSG00000111850 2.28058 5.72774 1.328565227 up small integral membrane protein 8 [Source:HGNC Symbol;Acc:21401] ENSG00000126070 4.95836 12.1416 1.292023623 up eukaryotic translation initiation factor 2C, 3 [Source:HGNC Symbol;Acc:18421] ENSG00000154781 6.58946 15.389 1.223667339 up coiled-coil domain containing 174 [Source:HGNC Symbol;Acc:28033] ENSG00000111490 5.02417 11.7049 1.220155423 up TBC1 domain family, member 30 [Source:HGNC Symbol;Acc:29164] ENSG00000112183 2.14386 4.84626 1.17666111 up RNA binding motif protein 24 [Source:HGNC Symbol;Acc:21539] ENSG00000162433 5.88931 13.2923 1.174420239 up adenylate kinase 4 [Source:HGNC Symbol;Acc:363] ENSG00000189144 3.3689 7.60021 1.173761677 up zinc finger protein 573 [Source:HGNC Symbol;Acc:26420] ENSG00000267100 1.88424 4.19986 1.156358501 up ILF3 antisense RNA 1 (head to head) [Source:HGNC Symbol;Acc:27115] ENSG00000147654 6.32929 13.8479 1.129551635 up estrogen receptor binding site associated, antigen, 9 [Source:HGNC Symbol;Acc:3123] ENSG00000180447 1.91579 4.12663 1.107024662 up growth arrest-specific 1 [Source:HGNC Symbol;Acc:4165] ENSG00000254870 17.3676 37.3119 1.103237423 up ATP6V1G2-DDX39B readthrough (non-protein coding) [Source:HGNC Symbol;Acc:41999] ENSG00000143622 14.5221 30.7865 1.08404777 up Ras-like without CAAX 1 [Source:HGNC Symbol;Acc:10023] ENSG00000168283 5.71889 12.116 1.083106422 up BMI1 polycomb ring finger oncogene [Source:HGNC Symbol;Acc:1066] ENSG00000178537 8.94604 18.9266 1.081094153 up solute carrier family 25 (carnitine/acylcarnitine translocase), member 20 [Source:HGNC Symbol;Acc:1421] ENSG00000047617 6.34087 13.2084 1.058703012 up anoctamin 2 [Source:HGNC Symbol;Acc:1183] ENSG00000160404 9.5724 19.6562 1.038031853 up torsin family 2, member A [Source:HGNC Symbol;Acc:11996] ENSG00000116711 2.30078 4.72253 1.037436921 up phospholipase A2, group IVA (cytosolic, calcium-dependent) [Source:HGNC Symbol;Acc:9035] ENSG00000183579 1.75972 3.58008 1.024645935 up zinc and ring finger 3 [Source:HGNC Symbol;Acc:18126] ENSG00000197415 5.96185 12.0142 1.010908603 up ventricular zone expressed PH domain-containing 1 [Source:HGNC Symbol;Acc:25735] ENSG00000198088 17.2525 8.51158 −1.019306564 down nucleoporin 62kDa C-terminal like [Source:HGNC Symbol;Acc:25960] ENSG00000179046 10.0612 4.93927 −1.026432646 down tripartite motif family-like 2 [Source:HGNC Symbol;Acc:26378] ENSG00000085117 16.3935 8.02257 −1.030987524 down CD82 molecule [Source:HGNC Symbol;Acc:6210] ENSG00000270882 13.1691 6.38904 −1.043485676 down − ENSG00000167535 11.2491 5.40623 −1.057114786 down calcium channel, voltage-dependent, beta 3 subunit [Source:HGNC Symbol;Acc:1403] ENSG00000187764 5.71625 2.7422 −1.059735217 down sema domain, immunoglobulin domain (Ig), transmembrane domain (TM) and short cytoplasmic domain, (semaphorin) 4D [Source:HGNC Symbol;Acc:10732] ENSG00000115421 11.6833 5.54962 −1.073986933 down poly(A) polymerase gamma [Source:HGNC Symbol;Acc:14982] ENSG00000115947 25.359 11.9667 −1.083472494 down origin recognition complex, subunit 4 [Source:HGNC Symbol;Acc:8490] ENSG00000168143 4.12583 1.93689 −1.090942355 down family with sequence similarity 83, member B [Source:HGNC Symbol;Acc:21357] ENSG00000137868 4.36322 2.03399 −1.101080632 down stimulated by retinoic acid 6 [Source:HGNC Symbol;Acc:30650] ENSG00000184517 11.179 5.18427 −1.108578378 down ZFP1 zinc finger protein [Source:HGNC Symbol;Acc:23328] ENSG00000166025 8.62556 3.98626 −1.113582313 down angiomotin like 1 [Source:HGNC Symbol;Acc:17811] ENSG00000164506 6.06679 2.79463 −1.118276086 down syntaxin binding protein 5 (tomosyn) [Source:HGNC Symbol;Acc:19665] ENSG00000131374 8.96638 4.11688 −1.122974246 down TBC1 domain family, member 5 [Source:HGNC Symbol;Acc:19166] ENSG00000157514 13.3161 6.03102 −1.142697685 down TSC22 domain family, member 3 [Source:HGNC Symbol;Acc:3051] ENSG00000204396 11.6324 5.11676 −1.184846314 down von Willebrand factor A domain containing 7 [Source:HGNC Symbol;Acc:13939] ENSG00000114861 7.03474 3.06923 −1.196620342 down forkhead box P1 [Source:HGNC Symbol;Acc:3823] ENSG00000164808 46.0156 20.0548 −1.198175461 down KIAA0146 [Source:HGNC Symbol;Acc:28971] ENSG00000281181 16.7705 7.1703 −1.225820316 down − ENSG00000167081 8.3093 3.50186 −1.246605537 down pre-B-cell leukemia homeobox 3 [Source:HGNC Symbol;Acc:8634] ENSG00000214174 4.86578 2.03253 −1.259394446 down archaelysin family metallopeptidase 2 pseudogene 1 [Source:HGNC Symbol;Acc:26491] ENSG00000072195 8.30099 3.33036 −1.317605271 down SPEG complex locus [Source:HGNC Symbol;Acc:16901] ENSG00000109458 3.66758 1.44769 −1.34107573 down GRB2-associated binding protein 1 [Source:HGNC Symbol;Acc:4066] ENSG00000075043 5.86368 2.26188 −1.374283981 down potassium voltage-gated channel, KQT-like subfamily, member 2 [Source:HGNC Symbol;Acc:6296] ENSG00000131746 9.23984 3.52582 −1.389909046 down tensin 4 [Source:HGNC Symbol;Acc:24352] ENSG00000156453 4.01672 1.50392 −1.417290071 down protocadherin 1 [Source:HGNC Symbol;Acc:8655] ENSG00000188517 6.04896 2.24749 −1.428372424 down collagen, type XXV, alpha 1 [Source:HGNC Symbol;Acc:18603] ENSG00000265096 8.69038 3.22807 −1.428747399 down Uncharacterized protein [Source:UniProtKB/TrEMBL;Acc:J3KS61] ENSG00000151689 9.40788 3.47631 −1.436312918 down inositol polyphosphate-1-phosphatase [Source:HGNC Symbol;Acc:6071] ENSG00000088756 4.70196 1.68862 −1.477417559 down Rho GTPase activating protein 28 [Source:HGNC Symbol;Acc:25509] ENSG00000104081 5.96943 2.12103 −1.492828154 down Bcl2 modifying factor [Source:HGNC Symbol;Acc:24132] ENSG00000118194 4.6428 1.62065 −1.518422578 down troponin T type 2 (cardiac) [Source:HGNC Symbol;Acc:11949] ENSG00000188266 6.21225 2.05372 −1.596876388 down aminoglycoside phosphotransferase domain containing 1 [Source:HGNC Symbol;Acc:34403] ENSG00000172927 9.72759 3.20937 −1.599792299 down myeloma overexpressed (in a subset of t(11;14) positive multiple myelomas) [Source:HGNC Symbol;Acc:7563] ENSG00000278932 9.65933 3.14891 −1.617070599 down − ENSG00000107614 3.78983 1.11699 −1.762516865 down tRNA aspartic acid methyltransferase 1 [Source:HGNC Symbol;Acc:2977] ENSG00000152076 3.91793 1.15134 −1.766777686 down coiled-coil domain containing 74B [Source:HGNC Symbol;Acc:25267] ENSG00000159788 5.99872 1.73903 −1.786371872 down regulator of G-protein signaling 12 [Source:HGNC Symbol;Acc:9994] ENSG00000232973 5.70581 1.57831 −1.85405111 down CYP1B1 antisense RNA 1 [Source:HGNC Symbol;Acc:28543] ENSG00000067141 4.28714 1.16488 −1.879834187 down neogenin 1 [Source:HGNC Symbol;Acc:7754] ENSG00000203326 25.5951 6.93345 −1.884222339 down zinc finger protein 525 [Source:HGNC Symbol;Acc:29423] ENSG00000163995 4.71407 1.25699 −1.907000009 down actin binding LIM protein family, member 2 [Source:HGNC Symbol;Acc:19195] ENSG00000280130 3.76176 0 −1.911407808 down − ENSG00000240364 3.86536 0 −1.950602786 down ribosomal protein L31 pseudogene 59 [Source:HGNC Symbol;Acc:36975] ENSG00000174640 4.63741 1.17475 −1.980965515 down solute carrier organic anion transporter family, member 2A1 [Source:HGNC Symbol;Acc:10955] ENSG00000133401 4.00349 0.995203 −2.008195463 down PDZ domain containing 2 [Source:HGNC Symbol;Acc:18486] ENSG00000278903 11.7426 2.82235 −2.056783067 down − ENSG00000204406 6.34622 1.49977 −2.081156265 down methyl-CpG binding domain protein 5 [Source:HGNC Symbol;Acc:20444] ENSG00000280987 8.10572 1.90811 −2.086795997 down − ENSG00000108387 7.70484 1.56887 −2.296039185 down septin 4 [Source:HGNC Symbol;Acc:9165] ENSG00000253492 5.21647 0 −2.383073861 down cadherin 12 (N-cadherin 2) pseudogene 3 [Source:HGNC Symbol;Acc:37699] ENSG00000184271 7.99757 1.44707 −2.466427003 down POU class 6 homeobox 1 [Source:HGNC Symbol;Acc:9224] ENSG00000249684 5.82118 0 −2.541311629 down − ENSG00000185324 207.869 31.9115 −2.703526292 down cyclin-dependent kinase 10 [Source:HGNC Symbol;Acc:1770] ENSG00000251141 12.3399 1.88182 −2.71313016 down − ENSG00000130208 6.13461 0.690214 −3.151855987 down apolipoprotein C-I [Source:HGNC Symbol;Acc:607] ENSG00000270808 9.54955 0 −3.255432751 down − ENSG00000257341 6.31322 0.598144 −3.399811274 down − ENSG00000266210 11.7633 0 −3.556220936 down Metazoan signal recognition particle RNA [Source:RFAM;Acc:RF00017] ENSG00000189283 15.9633 0.425823 −5.228361236 down fragile histidine triad [Source:HGNC Symbol;Acc:3701] ENSG00000277739 85.2919 0 −6.414336833 down −
doi: 10.3967/bes2022.028
SET8 Inhibition Potentiates Radiotherapy by Suppressing DNA Damage Repair in Carcinomas
-
Abstract:
Objective SET8 is a member of the SET domain-containing family and the only known lysine methyltransferase (KMT) that monomethylates lysine 20 of histone H4 (H4K20me1). SET8 has been implicated in many essential cellular processes, including cell cycle regulation, DNA replication, DNA damage response, and carcinogenesis. There is no conclusive evidence, however, regarding the effect of SET8 on radiotherapy. In the current study we determined the efficacy of SET8 inhibition on radiotherapy of tumors and the underlying mechanism. Methods First, we explored the radiotherapy benefit of the SET8 expression signature by analyzing clinical data. Then, we measured a series of biological endpoints, including the xenograft tumor growth in mice and apoptosis, frequency of micronuclei, and foci of 53BP1 and γ-H2AX in cells to detect the SET8 effects on radiosensitivity. RNA sequencing and subsequent experiments were exploited to verify the mechanism underlying the SET8 effects on radiotherapy. Results Low expression of SET8 predicted a better benefit to radiotherapy in lung adenocarcinoma (LUAD) and invasive breast carcinoma (BRCA) patients. Furthermore, genetic deletion of SET8 significantly enhanced radiation treatment efficacy in a murine tumor model, and A549 and MCF7 cells; SET8 overexpression decreased the radiosensitivity. SET8 inhibition induced more apoptosis, the frequency of micronuclei, and blocked the kinetics process of DNA damage repair as 53BP1 and γ-H2AX foci remained in cells. Moreover, RNF8 was positively correlated with the SET8 impact on DNA damage repair. Conclusion Our results demonstrated that SET8 inhibition enhanced radiosensitivity by suppressing DNA damage repair, thus suggesting that SET8 potentiated radiotherapy of carcinomas. As new inhibitors of SET8 are synthesized and tested in preclinical and clinical settings, combining SET8 inhibitors with radiation warrants consideration for precise radiotherapy. -
Key words:
- SET8 /
- Histone methylation /
- DNA repair /
- Radiotherapy /
- Carcinoma
-
Figure 1. Kaplan-Meier survival curves of radiotherapy treated LUAD and BRCA patients with different SET8 expression. (A) Kaplan-Meier survival curves of radiotherapy-treated LUAD patients with low or high SET8 expression. (B) Kaplan-Meier survival curves of radiotherapy-treated BRCA patients with low or high SET8 expression. (C) Kaplan-Meier survival curves of radiotherapy untreated LUAD patients with low or high SET8 expression. (D) Kaplan-Meier survival curves of radiotherapy untreated BRCA patients with low or high SET8 expression.The cut-off values of high and low are top 25% and lowest 25% in LUAD, top 10% and lowest 10% in BRCA. P-values calculated by use of logrank test.
Figure 2. Radiosensitivity in the SET8 alteration A549 and MCF7 cells. (A) Western blot analysis of SET8 levels in vector control (VC) and SET8 KO groups B16F10 cells. (B) & (C) Tumor growth delay and Kaplan-Meier survival curves of C57BL/6 mice inoculated with 1 x 105 control or SE8-KO LLC1 cells. Radiotherapy was conducted 1 x at 8 Gy at 7 days post inoculation of the tumor cells. (D) Western blot assay of SET8 and H4K20me1 protein levels in SET8-KO or control A549 and MCF7 cells. (E) & (F) Colony formation assay of SET8-KO and control A549 and MCF7 cells were exposed to X rays or Carbon. (G) Western blot assay of SET8 and H4K20me1 protein level in SET8 overexpression (Over-SET8) and control A549 and MCF7 cells. (H) & (I) Colony formation assay of Over-SET8 and control A549 and MCF7 cells were exposed to X rays or Carbon. Significance was determined by 2-way ANOVA in (B), logrank test in c and unpaired t-test in (F) and (H) respectively. *P < 0.05; **P < 0.01, ***P < 0.001.
S1. SET8 expressions and H4K20me1 dynamic changes after X-ray or carbon ion radiation. (A) Relative SET8 mRNA expression was measured by qRT-PCR at indicated time points in A549 cells after 4 Gy X-ray irradiation. GAPDH was used as internal controls. (B) SET8 and H4K20me1 levels in A549 cells at indicated time points after 4 Gy X-ray radiation were measured by western blot assay. (C) Relative SET8 mRNA expression was measured by qRT-PCR at indicated time points in A549 cells after 2 Gy Carbon irradiation. (D) SET8 and H4K20me1 levels in A549 cells at indicated time points after 2 Gy Carbon radiation were measured by western blot assay. Significance was determined by unpaired t-test. *P < 0.05; **P < 0.01.
Figure 3. Cellular apoptosis and nucleus injury in irradiated SET8 alteration A549 and MCF7 cells. (A) & (B) Apoptosis proportion of SET8-KO or control A549 and MCF7 cells were exposed to 4 Gy X rays 48 h later. (C) & (D) Micronucleus frequency of SET8-KO or control A549 and MCF7 cells were exposed to 4 Gy X rays 48 h later. (E) & (F) Micronucleus frequency of Over-SET8 and control A549 and MCF7 cells were exposed to 4 Gy X rays 48 h later. Significance was determined by unpaired t-test. *P < 0.05. IR, irradiation. VC, vector control.
Figure 4. Single-cell RNA sequencing analyses between SET8 expression and DNA repair scores in LUAD and BRCA. (A) Scores of DNA repaired through single-cell RNA sequencing analyses in LUAD with SET8 expression low or high signature. (B) Scores of DNA repaired through single-cell RNA sequencing analyses in BRCA with SET8 expression low or high signature. The cut-off values of high and low are top 25% and lowest 25% in LUAD and BRCA . P-values calculated by use of unpaired t-test .
Figure 5. Kinetics process of DNA damage repair in SET8 knockdown A549 cells. (A) Graph shows quantification of 53BP1 and γH2AX foci in SET8-KO or control A549 cells exposed to 1 Gy X rays. (B) The numbers of 53BP1 foci in 50 cells of each group were counted for each time point. (C) The numbers of γH2AX foci in 50 cells of each group were counted for each time point. Significance was determined by unpaired t-test . *P < 0.05. VC, vector control.
Figure 6. Radiosensitivity in the SET8 alteration A549 and MCF7 cells. (A) Heat map of RNA sequnece analysis in vector control (VC) and SET8 KO groups A549 cells, 12 h after 4Gy X-ray radiation. Genes that were significantly differentially expressed (P < 0.05) and had greater than 2-fold difference in expression were identified. (B) Venn diagram showing the total number of gene comparisons between different treatments groups. (C) & (D) Representative immunofluorescence images and RNF8 relative integrated immunofluorescence intensity in control and SET8-KO A549 cells. (E) Western blot assay of SET8, RNF8 and H4K20me1 protein levels in SET8-KO or control A549 and MCF7 cells exposed to 4 Gy X rays after 8 h. Significance was determined by unpaired t-test in (D). **P < 0.01. IR, irradiation
S1. sgRNA used in CRISPR/Cas9 knockout experiments
Target Sequence (5'–3') Human-SET8-sg1 TTTAGGAGAACGTATTTACC Human-SET8-sg2 GCACGTGACTACCTGCAGCT Mouse-SET8-sg1 AGTATCTGAGCAAAACCTAC Mouse-SET8-sg1 TCCGGGAACTCCTCCGCACA  下载: 导出CSV
下载: 导出CSV
S2. Oligonucleotide primers for qRT-PCR
Genes Upstream primer (5'–3') Downstream primer (5'–3') SET8 CGCAAACTTACGGATTTCT CGATGAGGTCAATCTTCATT
GAPDH
GTCTCCTCTGACTTCAACAGCG
ACCACCCTGTTGCTGTAGCCAA
下载: 导出CSV
S3. Gene expression signatures in SET8-KO A549 cells
Seq_id (VC) fpkm (KO)3_7_fpkm log2FC(KO/VC) Regulate Description ENSG00000264148 0 49.1764 5.61989422 up − ENSG00000236991 0.330355 3.76492 3.510530124 up − ENSG00000088448 41.5497 364.193 3.131793228 up ankyrin repeat domain 10 [Source:HGNC Symbol;Acc:20265] ENSG00000185437 0 8.08694 3.015593908 up SH3 domain binding glutamic acid-rich protein [Source:HGNC Symbol;Acc:10822] ENSG00000228950 0 7.66067 2.937470575 up − ENSG00000271267 0 5.72202 2.51652454 up − ENSG00000232859 1.14869 6.19551 2.43123354 up LYR motif containing 9 [Source:HGNC Symbol;Acc:27314] ENSG00000172339 1.08544 5.63913 2.377192622 up asparagine-linked glycosylation 14 homolog (S. cerevisiae) [Source:HGNC Symbol;Acc:28287] ENSG00000213420 1.27423 5.78869 2.183611189 up glypican 2 [Source:HGNC Symbol;Acc:4450] ENSG00000131095 0.961888 4.15931 2.112403391 up glial fibrillary acidic protein [Source:HGNC Symbol;Acc:4235] ENSG00000185920 3.25398 13.6198 2.065428229 up patched 1 [Source:HGNC Symbol;Acc:9585] ENSG00000234160 0 3.97046 1.989306161 up − ENSG00000241769 1.82417 7.01286 1.942762742 up − ENSG00000153291 1.65069 6.24826 1.92038528 up solute carrier family 25, member 27 [Source:HGNC Symbol;Acc:21065] ENSG00000254335 0 3.74532 1.905088988 up cadherin 12 (N-cadherin 2) pseudogene 1 [Source:HGNC Symbol;Acc:37698] ENSG00000230438 1.00366 3.68563 1.876640622 up − ENSG00000170899 1.78022 5.97155 1.746049911 up glutathione S-transferase alpha 4 [Source:HGNC Symbol;Acc:4629] ENSG00000104205 1.80144 5.94424 1.722341765 up serum/glucocorticoid regulated kinase family, member 3 [Source:HGNC Symbol;Acc:10812] ENSG00000139946 1.0563 3.48076 1.72038271 up pellino E3 ubiquitin protein ligase family member 2 [Source:HGNC Symbol;Acc:8828] ENSG00000137843 1.61001 5.23319 1.70062099 up p21 protein (Cdc42/Rac)-activated kinase 6 [Source:HGNC Symbol;Acc:16061] ENSG00000246273 6.43651 18.854 1.550520085 up SBF2 antisense RNA 1 [Source:HGNC Symbol;Acc:27438] ENSG00000176371 3.07709 8.49372 1.464829909 up zinc finger and SCAN domain containing 2 [Source:HGNC Symbol;Acc:20994] ENSG00000080298 1.3528 3.72538 1.461439027 up regulatory factor X, 3 (influences HLA class II expression) [Source:HGNC Symbol;Acc:9984] ENSG00000133794 5.24692 13.2361 1.334935398 up aryl hydrocarbon receptor nuclear translocator-like [Source:HGNC Symbol;Acc:701] ENSG00000111850 2.28058 5.72774 1.328565227 up small integral membrane protein 8 [Source:HGNC Symbol;Acc:21401] ENSG00000126070 4.95836 12.1416 1.292023623 up eukaryotic translation initiation factor 2C, 3 [Source:HGNC Symbol;Acc:18421] ENSG00000154781 6.58946 15.389 1.223667339 up coiled-coil domain containing 174 [Source:HGNC Symbol;Acc:28033] ENSG00000111490 5.02417 11.7049 1.220155423 up TBC1 domain family, member 30 [Source:HGNC Symbol;Acc:29164] ENSG00000112183 2.14386 4.84626 1.17666111 up RNA binding motif protein 24 [Source:HGNC Symbol;Acc:21539] ENSG00000162433 5.88931 13.2923 1.174420239 up adenylate kinase 4 [Source:HGNC Symbol;Acc:363] ENSG00000189144 3.3689 7.60021 1.173761677 up zinc finger protein 573 [Source:HGNC Symbol;Acc:26420] ENSG00000267100 1.88424 4.19986 1.156358501 up ILF3 antisense RNA 1 (head to head) [Source:HGNC Symbol;Acc:27115] ENSG00000147654 6.32929 13.8479 1.129551635 up estrogen receptor binding site associated, antigen, 9 [Source:HGNC Symbol;Acc:3123] ENSG00000180447 1.91579 4.12663 1.107024662 up growth arrest-specific 1 [Source:HGNC Symbol;Acc:4165] ENSG00000254870 17.3676 37.3119 1.103237423 up ATP6V1G2-DDX39B readthrough (non-protein coding) [Source:HGNC Symbol;Acc:41999] ENSG00000143622 14.5221 30.7865 1.08404777 up Ras-like without CAAX 1 [Source:HGNC Symbol;Acc:10023] ENSG00000168283 5.71889 12.116 1.083106422 up BMI1 polycomb ring finger oncogene [Source:HGNC Symbol;Acc:1066] ENSG00000178537 8.94604 18.9266 1.081094153 up solute carrier family 25 (carnitine/acylcarnitine translocase), member 20 [Source:HGNC Symbol;Acc:1421] ENSG00000047617 6.34087 13.2084 1.058703012 up anoctamin 2 [Source:HGNC Symbol;Acc:1183] ENSG00000160404 9.5724 19.6562 1.038031853 up torsin family 2, member A [Source:HGNC Symbol;Acc:11996] ENSG00000116711 2.30078 4.72253 1.037436921 up phospholipase A2, group IVA (cytosolic, calcium-dependent) [Source:HGNC Symbol;Acc:9035] ENSG00000183579 1.75972 3.58008 1.024645935 up zinc and ring finger 3 [Source:HGNC Symbol;Acc:18126] ENSG00000197415 5.96185 12.0142 1.010908603 up ventricular zone expressed PH domain-containing 1 [Source:HGNC Symbol;Acc:25735] ENSG00000198088 17.2525 8.51158 −1.019306564 down nucleoporin 62kDa C-terminal like [Source:HGNC Symbol;Acc:25960] ENSG00000179046 10.0612 4.93927 −1.026432646 down tripartite motif family-like 2 [Source:HGNC Symbol;Acc:26378] ENSG00000085117 16.3935 8.02257 −1.030987524 down CD82 molecule [Source:HGNC Symbol;Acc:6210] ENSG00000270882 13.1691 6.38904 −1.043485676 down − ENSG00000167535 11.2491 5.40623 −1.057114786 down calcium channel, voltage-dependent, beta 3 subunit [Source:HGNC Symbol;Acc:1403] ENSG00000187764 5.71625 2.7422 −1.059735217 down sema domain, immunoglobulin domain (Ig), transmembrane domain (TM) and short cytoplasmic domain, (semaphorin) 4D [Source:HGNC Symbol;Acc:10732] ENSG00000115421 11.6833 5.54962 −1.073986933 down poly(A) polymerase gamma [Source:HGNC Symbol;Acc:14982] ENSG00000115947 25.359 11.9667 −1.083472494 down origin recognition complex, subunit 4 [Source:HGNC Symbol;Acc:8490] ENSG00000168143 4.12583 1.93689 −1.090942355 down family with sequence similarity 83, member B [Source:HGNC Symbol;Acc:21357] ENSG00000137868 4.36322 2.03399 −1.101080632 down stimulated by retinoic acid 6 [Source:HGNC Symbol;Acc:30650] ENSG00000184517 11.179 5.18427 −1.108578378 down ZFP1 zinc finger protein [Source:HGNC Symbol;Acc:23328] ENSG00000166025 8.62556 3.98626 −1.113582313 down angiomotin like 1 [Source:HGNC Symbol;Acc:17811] ENSG00000164506 6.06679 2.79463 −1.118276086 down syntaxin binding protein 5 (tomosyn) [Source:HGNC Symbol;Acc:19665] ENSG00000131374 8.96638 4.11688 −1.122974246 down TBC1 domain family, member 5 [Source:HGNC Symbol;Acc:19166] ENSG00000157514 13.3161 6.03102 −1.142697685 down TSC22 domain family, member 3 [Source:HGNC Symbol;Acc:3051] ENSG00000204396 11.6324 5.11676 −1.184846314 down von Willebrand factor A domain containing 7 [Source:HGNC Symbol;Acc:13939] ENSG00000114861 7.03474 3.06923 −1.196620342 down forkhead box P1 [Source:HGNC Symbol;Acc:3823] ENSG00000164808 46.0156 20.0548 −1.198175461 down KIAA0146 [Source:HGNC Symbol;Acc:28971] ENSG00000281181 16.7705 7.1703 −1.225820316 down − ENSG00000167081 8.3093 3.50186 −1.246605537 down pre-B-cell leukemia homeobox 3 [Source:HGNC Symbol;Acc:8634] ENSG00000214174 4.86578 2.03253 −1.259394446 down archaelysin family metallopeptidase 2 pseudogene 1 [Source:HGNC Symbol;Acc:26491] ENSG00000072195 8.30099 3.33036 −1.317605271 down SPEG complex locus [Source:HGNC Symbol;Acc:16901] ENSG00000109458 3.66758 1.44769 −1.34107573 down GRB2-associated binding protein 1 [Source:HGNC Symbol;Acc:4066] ENSG00000075043 5.86368 2.26188 −1.374283981 down potassium voltage-gated channel, KQT-like subfamily, member 2 [Source:HGNC Symbol;Acc:6296] ENSG00000131746 9.23984 3.52582 −1.389909046 down tensin 4 [Source:HGNC Symbol;Acc:24352] ENSG00000156453 4.01672 1.50392 −1.417290071 down protocadherin 1 [Source:HGNC Symbol;Acc:8655] ENSG00000188517 6.04896 2.24749 −1.428372424 down collagen, type XXV, alpha 1 [Source:HGNC Symbol;Acc:18603] ENSG00000265096 8.69038 3.22807 −1.428747399 down Uncharacterized protein [Source:UniProtKB/TrEMBL;Acc:J3KS61] ENSG00000151689 9.40788 3.47631 −1.436312918 down inositol polyphosphate-1-phosphatase [Source:HGNC Symbol;Acc:6071] ENSG00000088756 4.70196 1.68862 −1.477417559 down Rho GTPase activating protein 28 [Source:HGNC Symbol;Acc:25509] ENSG00000104081 5.96943 2.12103 −1.492828154 down Bcl2 modifying factor [Source:HGNC Symbol;Acc:24132] ENSG00000118194 4.6428 1.62065 −1.518422578 down troponin T type 2 (cardiac) [Source:HGNC Symbol;Acc:11949] ENSG00000188266 6.21225 2.05372 −1.596876388 down aminoglycoside phosphotransferase domain containing 1 [Source:HGNC Symbol;Acc:34403] ENSG00000172927 9.72759 3.20937 −1.599792299 down myeloma overexpressed (in a subset of t(11;14) positive multiple myelomas) [Source:HGNC Symbol;Acc:7563] ENSG00000278932 9.65933 3.14891 −1.617070599 down − ENSG00000107614 3.78983 1.11699 −1.762516865 down tRNA aspartic acid methyltransferase 1 [Source:HGNC Symbol;Acc:2977] ENSG00000152076 3.91793 1.15134 −1.766777686 down coiled-coil domain containing 74B [Source:HGNC Symbol;Acc:25267] ENSG00000159788 5.99872 1.73903 −1.786371872 down regulator of G-protein signaling 12 [Source:HGNC Symbol;Acc:9994] ENSG00000232973 5.70581 1.57831 −1.85405111 down CYP1B1 antisense RNA 1 [Source:HGNC Symbol;Acc:28543] ENSG00000067141 4.28714 1.16488 −1.879834187 down neogenin 1 [Source:HGNC Symbol;Acc:7754] ENSG00000203326 25.5951 6.93345 −1.884222339 down zinc finger protein 525 [Source:HGNC Symbol;Acc:29423] ENSG00000163995 4.71407 1.25699 −1.907000009 down actin binding LIM protein family, member 2 [Source:HGNC Symbol;Acc:19195] ENSG00000280130 3.76176 0 −1.911407808 down − ENSG00000240364 3.86536 0 −1.950602786 down ribosomal protein L31 pseudogene 59 [Source:HGNC Symbol;Acc:36975] ENSG00000174640 4.63741 1.17475 −1.980965515 down solute carrier organic anion transporter family, member 2A1 [Source:HGNC Symbol;Acc:10955] ENSG00000133401 4.00349 0.995203 −2.008195463 down PDZ domain containing 2 [Source:HGNC Symbol;Acc:18486] ENSG00000278903 11.7426 2.82235 −2.056783067 down − ENSG00000204406 6.34622 1.49977 −2.081156265 down methyl-CpG binding domain protein 5 [Source:HGNC Symbol;Acc:20444] ENSG00000280987 8.10572 1.90811 −2.086795997 down − ENSG00000108387 7.70484 1.56887 −2.296039185 down septin 4 [Source:HGNC Symbol;Acc:9165] ENSG00000253492 5.21647 0 −2.383073861 down cadherin 12 (N-cadherin 2) pseudogene 3 [Source:HGNC Symbol;Acc:37699] ENSG00000184271 7.99757 1.44707 −2.466427003 down POU class 6 homeobox 1 [Source:HGNC Symbol;Acc:9224] ENSG00000249684 5.82118 0 −2.541311629 down − ENSG00000185324 207.869 31.9115 −2.703526292 down cyclin-dependent kinase 10 [Source:HGNC Symbol;Acc:1770] ENSG00000251141 12.3399 1.88182 −2.71313016 down − ENSG00000130208 6.13461 0.690214 −3.151855987 down apolipoprotein C-I [Source:HGNC Symbol;Acc:607] ENSG00000270808 9.54955 0 −3.255432751 down − ENSG00000257341 6.31322 0.598144 −3.399811274 down − ENSG00000266210 11.7633 0 −3.556220936 down Metazoan signal recognition particle RNA [Source:RFAM;Acc:RF00017] ENSG00000189283 15.9633 0.425823 −5.228361236 down fragile histidine triad [Source:HGNC Symbol;Acc:3701] ENSG00000277739 85.2919 0 −6.414336833 down −
下载: 导出CSV
-
[1] Smith RA, Andrews KS, Brooks D, et al. Cancer screening in the United States, 2019: a review of current American Cancer Society guidelines and current issues in cancer screening. CA Cancer J Clin, 2019; 69, 184−210. doi: 10.3322/caac.21557 [2] Sawan C, Herceg Z. Histone modifications and cancer. Adv Genet, 2010; 70, 57−85. [3] McCabe MT, Mohammad HP, Barbash O, et al. Targeting histone methylation in cancer. Cancer J, 2017; 23, 292−301. doi: 10.1097/PPO.0000000000000283 [4] Nishioka K, Rice JC, Sarma K, et al. PR-Set7 is a nucleosome-specific methyltransferase that modifies lysine 20 of histone H4 and is associated with silent chromatin. Mol Cell, 2002; 9, 1201−13. doi: 10.1016/S1097-2765(02)00548-8 [5] Shi XB, Kachirskaia I, Yamaguchi H, et al. Modulation of p53 function by set8-mediated methylation at lysine 382. Mol Cell, 2007; 27, 636−46. doi: 10.1016/j.molcel.2007.07.012 [6] Yu N, Huangyang PW, Yang XH, et al. MicroRNA-7 suppresses the invasive potential of breast cancer cells and sensitizes cells to DNA damages by targeting histone methyltransferase SET8. J Biol Chem, 2013; 288, 19633−42. doi: 10.1074/jbc.M113.475657 [7] Beck DB, Oda H, Shen SS, et al. PR-Set7 and H4K20me1: at the crossroads of genome integrity, cell cycle, chromosome condensation, and transcription. Genes Dev, 2012; 26, 325−37. doi: 10.1101/gad.177444.111 [8] Huen MSY, Sy SMH, van Deursen JM, et al. Direct interaction between SET8 and proliferating cell nuclear antigen couples H4-K20 methylation with DNA replication. J Biol Chem, 2008; 283, 11073−7. doi: 10.1074/jbc.C700242200 [9] Houston SI, McManus KJ, Adams MM, et al. Catalytic function of the PR-Set7 histone H4 lysine 20 monomethyltransferase is essential for mitotic entry and genomic stability. J Biol Chem, 2008; 283, 19478−88. doi: 10.1074/jbc.M710579200 [10] Jørgensen S, Schotta G, Sørensen CS. Histone H4 lysine 20 methylation: key player in epigenetic regulation of genomic integrity. Nucleic Acids Res, 2013; 41, 2797−806. doi: 10.1093/nar/gkt012 [11] Cerami E, Gao JJ, Dogrusoz U, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov, 2012; 2, 401−4. doi: 10.1158/2159-8290.CD-12-0095 [12] Hoadley KA, Yau C, Hinoue T, et al. Cell-of-origin patterns dominate the molecular classification of 10, 000 tumors from 33 types of cancer. Cell, 2018; 173, 291−304.e6. doi: 10.1016/j.cell.2018.03.022 [13] Sanjana NE, Shalem O, Zhang F. Improved vectors and genome-wide libraries for CRISPR screening. Nat Methods, 2014; 11, 783−4. doi: 10.1038/nmeth.3047 [14] Puchtler H, Sweat Waldrop F, Conner HM, et al. Carnoy fixation: practical and theoretical considerations. Histochemie, 1968; 16, 361−71. doi: 10.1007/BF00306359 [15] Hayashi M. The micronucleus test—most widely used in vivo genotoxicity test—. Genes Environ, 2016; 38, 18. doi: 10.1186/s41021-016-0044-x [16] Savage JRK. A comment on the quantitative relationship between micronuclei and chromosomal aberrations. Mutat Res Lett, 1988; 207, 33−6. doi: 10.1016/0165-7992(88)90008-5 [17] Yuan HT, Yan M, Zhang GX, et al. CancerSEA: a cancer single-cell state atlas. Nucleic Acids Res, 2019; 47, D900−8. doi: 10.1093/nar/gky939 [18] Kim KT, Lee HW, Lee HO, et al. Single-cell mRNA sequencing identifies subclonal heterogeneity in anti-cancer drug responses of lung adenocarcinoma cells. Genome Biol, 2015; 16, 127. doi: 10.1186/s13059-015-0692-3 [19] Guillaumet-Adkins A, Rodríguez-Esteban G, Mereu E, et al. Single-cell transcriptome conservation in cryopreserved cells and tissues. Genome Biol, 2017; 18, 45. doi: 10.1186/s13059-017-1171-9 [20] Braune EB, Tsoi YL, Phoon YP, et al. Loss of CSL unlocks a hypoxic response and enhanced tumor growth potential in breast cancer cells. Stem Cell Reports, 2016; 6, 643−51. doi: 10.1016/j.stemcr.2016.03.004 [21] Jordan NV, Bardia A, Wittner BS, et al. HER2 expression identifies dynamic functional states within circulating breast cancer cells. Nature, 2016; 537, 102−6. doi: 10.1038/nature19328 [22] van Oorschot B, Hovingh SE, Moerland PD, et al. Reduced activity of double-strand break repair genes in prostate cancer patients with late normal tissue radiation toxicity. Int J Radiat Oncol Biol Phys, 2014; 88, 664−70. doi: 10.1016/j.ijrobp.2013.11.219 [23] Li F, Liu B, Zhou XL, et al. Silencing of E3 ubiquitin ligase RNF8 enhances ionizing radiation sensitivity of medulloblastoma cells by promoting the deubiquitination of PCNA. Oncol Res, 2018; 26, 1365−73. doi: 10.3727/096504018X15154085345907 [24] Pogribny I, Koturbash I, Tryndyak V, et al. Fractionated low-dose radiation exposure leads to accumulation of DNA damage and profound alterations in DNA and histone methylation in the murine thymus. Mol Cancer Res, 2005; 3, 553−61. doi: 10.1158/1541-7786.MCR-05-0074 [25] Maroschik B, Gürtler A, Krämer A, et al. Radiation-induced alterations of histone post-translational modification levels in lymphoblastoid cell lines. Radiat Oncol, 2014; 9, 15. doi: 10.1186/1748-717X-9-15 [26] Sak A, Kübler D, Bannik K, et al. Dependence of radiation-induced H2AX phosphorylation on histone methylation: evidence from the chromatin immunoprecipitation assay. Int J Radiat Biol, 2015; 91, 346−53. doi: 10.3109/09553002.2015.997895 [27] Game JC, Williamson MS, Spicakova T, et al. The RAD6/BRE1 histone modification pathway in saccharomyces confers radiation resistance through a RAD51-dependent process that is independent of RAD18. Genetics, 2006; 173, 1951−68. doi: 10.1534/genetics.106.057794 [28] Zhang BP, Li B, Cheng JY, et al. Anti-cancer effect of 20(S)-ginsenoside-Rh2 on oral squamous cell carcinoma cells via the decrease in ROS and downregulation of MMP-2 and VEGF. Biomed Environ Sci, 2020; 33, 713−7. [29] Dai XY, Song AY, Mu L, et al. Potential function of MMP3 gene in degradation of extracellular matrix complex in colorectal carcinoma. Biomed Environ Sci, 2021; 34, 66−70. [30] Lu X, Simon MD, Chodaparambil JV, et al. The effect of H3K79 dimethylation and H4K20 trimethylation on nucleosome and chromatin structure. Nat Struct Mol Biol, 2008; 15, 1122−4. doi: 10.1038/nsmb.1489 [31] Dulev S, Tkach J, Lin SC, et al. SET8 methyltransferase activity during the DNA double-strand break response is required for recruitment of 53BP1. EMBO Rep, 2014; 15, 1163−74. doi: 10.15252/embr.201439434 [32] Gursoy-Yuzugullu O, House N, Price BD. Patching broken DNA: nucleosome dynamics and the repair of DNA breaks. J Mol Biol, 2016; 428, 1846−60. doi: 10.1016/j.jmb.2015.11.021 [33] Zhao MJ, Song YF, Niu HT, et al. Adenovirus-mediated downregulation of the ubiquitin ligase RNF8 sensitizes bladder cancer to radiotherapy. Oncotarget, 2016; 7, 8956−67. doi: 10.18632/oncotarget.6909 [34] Tan XH, Peng J, Fu YB, et al. miR-638 mediated regulation of BRCA1affects DNA repair and sensitivity to UV and cisplatin in triple-negative breast cancer. Breast Cancer Res, 2014; 16, 435. doi: 10.1186/s13058-014-0435-5 [35] Zhou HX, Mu XQ, Chen J, et al. RNAi silencing targeting RNF8 enhances radiosensitivity of a non-small cell lung cancer cell line A549. Int J Radiat Biol, 2013; 89, 708−15. doi: 10.3109/09553002.2013.792964 [36] Blum G, Ibáñez G, Rao XJ, et al. Small-molecule inhibitors of SETD8 with cellular activity. ACS Chem Biol, 2014; 9, 2471−8. doi: 10.1021/cb500515r [37] Bromberg KD, Mitchell TRH, Upadhyay AK, et al. The SUV4-20 inhibitor A-196 verifies a role for epigenetics in genomic integrity. Nat Chem Biol, 2017; 13, 317−24. doi: 10.1038/nchembio.2282 [38] Gursoy-Yuzugullu O, Carman C, Serafim RB, et al. Epigenetic therapy with inhibitors of histone methylation suppresses DNA damage signaling and increases glioma cell radiosensitivity. Oncotarget, 2017; 8, 24518−32. doi: 10.18632/oncotarget.15543 -
 21300Supplementary Materials.pdf
21300Supplementary Materials.pdf

-

 点击查看大图
点击查看大图
计量
- 文章访问数: 1414
- HTML全文浏览量: 622
- PDF下载量: 96
- 被引次数: 0

 Quick Links
Quick Links