

 下载:
下载:





-
Campylobacter jejuni (C. jejuni) is a major cause of human gastroenteritis worldwide. It commonly colonizes the intestinal tract of poultry and other livestock. The number of campylobacteriosis cases in China is estimated to be much higher than in Europe and the USA[1].
Lipooligosaccharide (LOS) is an important virulence factor that may play a role in microbial adhesion and invasion. In addition, the sialylated LOS is the determinant structure, which mimics human gangliosides. The LOS loci of classes A, B, and C are the major genetic types related to the Guillain-Barré syndrome (GBS)[2-4].
Capsular polysaccharide (CPS) is another major virulence factor and is the serodeterminant of the Penner serotyping scheme for C. jejuni[5]. Some specific serotypes of C. jejuni strains, such as HS1/44, HS2, HS4c, HS10, HS19, HS23/36, and HS41[6-8], are associated with GBS. Previous studies in China suggest that HS2, HS19, and HS41 are dominant C. jejuni strains that trigger GBS [9-12].
In this study, the genetic characteristics of LOS and CPS of C. jejuni strains from different sources in China were analyzed by whole-genome sequencing (WGS), and a multiplex real-time Polymerase chain reaction (PCR) assay was developed for the rapid identification of three crucial GBS-associated CPS types in China.
-
A total of 494 C. jejuni strains, including 260 strains from diarrhea patients, 13 strains from GBS patients, 70 strains from livestock, and 151 strains from poultry, were collected for this study. All strains were identified as C. jejuni using previously described methods[13]. All tested strains were grown on Campylobacter selective medium (Karmali; Oxoid, Basingstoke, UK) supplemented with 5% sheep blood at 37 °C under microaerobic conditions (5% O2, 10% CO2, and 85% N2) for 48 h. DNA was extracted from each strain using the QIAamp DNA mini kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions for WGS.
-
The genome of C. jejuni isolates was sequenced using an Illumina HiSeq 2000 sequencing platform at the Beijing Genomics Institution with a depth of 450 × coverage. The low-quality reads were removed if the quality scores of ≥ 3 consecutive bases were ≤ Q30. The reads were then assembled into contigs and scaffolds using SOAPdenovo v2.04 (
http://soap.genomics.org.cn/soapdenovo.html ). PCR amplification and Sanger sequencing were used to fill the gaps in the region of the CPS/LOS sequences. The CPS/LOS locus sequences were annotated using Prokka V1.13.3[14] with default parameters and the genus-specific database from RefSeq [15].According to a previous study [16], the integrated sequences between the conserved genes kpsF and kpsC were considered the CPS biosynthesis gene cluster, and the LOS biosynthesis gene cluster was recognized by the genes waaC and waaF[17].
OrthoMCL v2.0.9 was used to cluster all CPS/LOS genes into orthologous and paralogous groups[18] (the accession numbers of CPS/LOS reference sequences used in this study are listed in Supplementary Tables S1 and S2 available in www.besjournal.com). A database of translated coding sequences for C. jejuni LOS/CPS biosynthesis was assembled according to the orthologue nomenclature described in a previous study[19]. Reciprocal all-versus-all BLASTP was performed (E-value < 1 × 10−5), and the results were processed by OrthoMCL with default parameters (percent match length ≥ 50%). A database was created containing orthologues and paralogues. Orthologues are homologous sequences derived from speciation events, and paralogues are homologous sequences derived from duplication events[20].
Table S1. The accession numbers for CPS gene clusters of C. jejuni
CPS types Accession number HS1 BX545859 HS2 AL111168.1 HS3 HQ343268 HS4c HQ343269 HS5/31 KT868847 HS6/S7 NC_009839 HS8/17 HQ343270 HS9 KT868844 HS10 HQ343271 HS11 KT868845 HS12 KT868848 HS15/31/58 HQ343272 HS18 KT932997 HS19 BX545860 HS21 KT868849 HS22 KT893439 HS23/36 BX545858 HS27 KT893437 HS29 KT868846 HS32 KT893435 HS33/35 KT893436 HS37 KT893431 HS38 KT893430 HS40 KT893434 HS41 BX545857 HS42 HQ343274 HS44 JF496678 HS45/5/32/60 KT893432 HS52 KT893429 HS53 CP000025.1 HS55 KT893433 HS57 KT893428 HS58/32 KT893427 HS60 KT893426 HS63 KT893438 Table S2. The accession numbers for LOS gene clusters of C. jejuni
LOS class Accession number A AF215659 B AF401528 C AF400047 D AF400669 E AJ131360 F AY423554 G AY436358 H AY800272 I EU404107 J EU404104 K AY573819 L EU404111 M EF140720 N AY816330 O EF143352 P AY943308 Q EU404112 R AY962325 S EU404110 T AIOC00000000 U AIOU00000000 V AIOP00000000 W CP001900 CDC1-10 reference[24] CDC11-19 In this study -
A serotype-specific sequence database for 33 CPS types (HS1, HS2, HS4 complex, HS5, HS6/7, HS8/17, HS9, HS10, HS11, HS12, HS15, HS18, HS19, HS21, HS22, HS27, HS29, HS31, HS32, HS33, HS37, HS38, HS40, HS41, HS42, HS44, HS45, HS52, HS53, HS55, HS57, HS58, and HS60) was created. Serotype-specific genes in each serotype were identified by a shell script run on CentOS release 6.9. This shell script requires several preinstalled bioinformatics software programs, including Samtools[21], BWA[22], Wgsim, and Megahit[23]. Briefly, Wgsim was used to simulate sequencing reads from each CPS reference sequence, and the reads were then mapped to other CPS reference sequences with BWA. The unmapped reads were obtained with Samtools and subsequently assembled with Megahit. The assembled contigs were subjected to BLAST in the NCBI database to eliminate the sequences mapped onto the CPS loci of other known serotypes. The serotype-specific sequence database was established through the above process. The CPS genotype for each strain in this study was determined based on searching and BLASTing with serotype-specific sequences in the database.
-
The optimized primers and probes for this study were designed using Primer-BLAST (Table 1). The reaction volume was 25 µL in total, containing 12.5 µL 2 × qPCR SuperMix, 2 μmol/L HS2-F and HS2-R, 3 μmol/L HS19-F and HS19-R, 3 μmol/L HS41-F and HS41-R, 1 μmol/L probe HS2, 1.5 μmol/L probe HS19, 1.5 μmol/L probe HS41 and 1 µL of DNA template. The thermocycling conditions were optimized as follows: initial denaturation at 94 °C for 30 s, followed by 40 cycles of 94 °C for 10 s and 60 °C for 30 s. This assay was further validated with 109 strains of known serotype strains. The DNA template of each strain was prepared after lysing whole bacterial cells by boiling. The Ct value of ≤ 35 was considered positive, and the result was considered negative when the Ct value was more than 40 or no amplification curve was obtained. Sensitivity and specificity were calculated for this multiplex real-time PCR assay. Sensitivity was calculated as the number of true positives divided by the sum of true positives and false negatives (true positive was defined as the Penner phenotypic serotyping results). Specificity was calculated as the number of true negatives divided by the number of true negatives and false positives.
Table 1. Primers and probes for real-time PCR detection of HS2, HS19, and HS41
Capsular type Primers and probes Nucleotide sequence (5’ to 3’) HS2 HS2-F AACCAACCATCCACTATTTTCATCT HS2-R ACTCTTTGATTTTCCAATGCAATGTT HS2-P FAM-CGCACCTTCCAATGCAACCAAAGAGC-BHQ1 HS19 HS19-F CTGAAATATCAACACAGGGAAAATG HS19-R TGTTGGAGGTAAAAGACATGGTG HS19-P HEX-TCCAATCCTGTTGCTCATAACGCATTCA-BHQ1 HS41 HS41-F CACCCTGATTTCTATTAACTCTACCAC HS41-R GAGTTATTCTTGATGCTGAAAATGGA HS41-P CY5-AAACGGGATAAGATCGCTTGGGGGAATA-BHQ2 Note. PCR, Polymerase chain reaction. -
Among the HS2, HS19, and HS41 strains identified by multiplex real-time PCR, the integrated CPS loci were selected for further analysis of genetic variation. CPS loci from six HS19 strains and three HS41 strains were selected. The phenotypic serotype was analyzed previously. Eleven CPS loci from HS2 strains were also selected for genetic variation analysis. The sequences of each CPS locus from the strains with the same CPS type were aligned with mafft v7.471. Variant calling was performed using the Find Variations/SNPs program on Geneious v2021.03 with the default settings.
-
The Chi-square test was used for all of the statistical analyses throughout the whole study. A P-value < 0.05 was considered to be statistically significant. All analyses were conducted with a statistical software package (IBM SPSS Statistics 19.0, Chicago, IL, USA).
-
A panel of 26 different capsular types was identified in 448 C. jejuni strains. The CPS genotypes of 46 strains (9.31%) could not be determined using serotype-specific sequences. The number of different CPS genotypes and the distribution of different CPS genotypes in strains from different sources are presented in Figure 1. The five most common CPS types, which accounted for 58.71% (263/448) of all the tested strains, were HS2 (90 strains, 20.08%), HS4c (70 strains, 15.63%), HS5/31 (40 strains, 8.93%), HS19 (35 strains, 7.81%), and HS8/17 (28 strains, 6.25%).
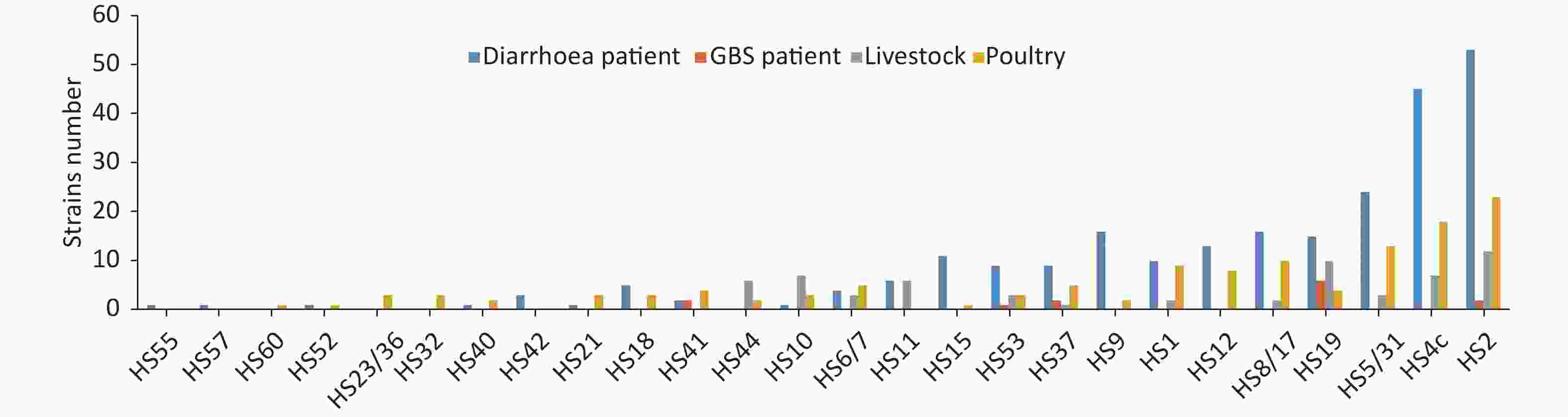
Figure 1. Distribution of different CPS types in strains from different sources: The CPS genotype for each strain was determined based on the different serospecific sequences. The top five CPS types were HS2 (90 strains, 20.1%), HS4c (70 strains, 15.6%), HS5/31 (40 strains, 8.9%), HS19 (35 strains, 7.8%), and HS8/17 (28 strains, 6.3%), which accounted for 58.7% (263/448) of the tested strains. CPS, capsular polysaccharide
The CPS genotypes of 12 strains (4.62%) from diarrhea patients could not be determined. The remaining 248 strains were classified into 22 CPS genotypes. The five most common CPS genotypes of the strains from diarrhea patients were HS2 (53 strains, 21.4%), HS4c (45 strains, 18.1%), HS5/31 (24 strains, 9.7%), HS19 (16 strains, 6.5%), and HS8/17 (16 strains, 6.5%). The CPS genotype of only one of the strains (7.7%) from GBS patients could not be determined. The remaining 12 GBS-associated strains consisted of five CPS types (HS19, HS37, HS2, HS53, and HS41). The CPS genotypes could not be determined for 11.43% (8/70) and 16.56% (25/151) of the strains from livestock and poultry, respectively. The five most common CPS genotypes for the strains from livestock were HS2 (12 strains, 17.14%), HS19 (10 strains, 14.29%), HS4c (7 strains, 10.0%), HS10 (7 strains, 10.0%), and HS11 (6 strains, 8.57%), and the five most common CPS types for the strains from poultry were HS2 (23 strains, 15.23%), HS4c (18 strains, 11.92%), HS5/31 (13 strains, 8.61%), HS8/17 (10 strains, 6.62%), and HS1 (9 strains, 5.96%). Except for the strains from GBS patients, HS2 and HS4c were the most frequent CPS types among the tested strains from different sources.
In total, 38 LOS classes were identified in 494 C. jejuni strains. Twenty-nine were previously confirmed LOS [3,24-26]. Nine novel classes were identified in 11 strains (9 from poultry and 2 from diarrhea patients) and were named CDC11–CDC19 (Figure 2). Four poultry strains (classified as CDC11, CDC15, CDC16, and CDC19) possessed the sialic acid synthesis genes ORF1 (cgtA/NeuA), ORF37 (cstII), ORF41 (NeuB), and ORF38 (NeuC) in the LOS locus. The distribution of the LOS classes in strains from different sources is presented in Table 2. LOS classes A, B, and C accounted for 12.35% (61/494), 28.54% (141/494), and 7.29% (36/494) of the tested strains, respectively. These three classes accounted for the largest proportion (48.18%, 238/494). Classes P, H, CDC5, F, CDC6, R, CDC2, W, CDC1, and G accounted for 38.46% (190/494) of the tested strains.
Table 2. The distribution of the LOS classes in strains from different sources
LOS class Diarrhoea
patientsGBS
patientsLivestock Poultry In total
(%)A 33 9 15 4 61 (12.35) B 73 0 27 41 141 (28.54) C 19 0 7 10 36 (7.29) D 0 0 0 1 1 (0.20) E 3 0 3 1 7 (1.42) F 9 1 6 2 18 (3.64) G 10 0 0 2 12 (2.43) H 24 1 2 4 31 (6.28) I 0 0 2 0 2 (0.40) J 2 0 0 0 2 (0.40) K 5 0 0 3 8 (1.62) M 0 0 0 1 1 (0.20) O 2 0 0 0 2 (0.40) P 21 2 2 8 33 (6.68) R 1 0 0 14 15 (3.04) S 5 0 3 1 9 (1.82) T 1 0 0 1 2 (0.40) V 1 0 1 2 4 (0.81) W 4 0 0 10 14 (2.83) CDC1 1 0 1 11 13 (2.63) CDC2 12 0 0 2 14 (2.83) CDC3 3 0 0 1 4 (0.81) CDC4 5 0 0 0 5 (1.01) CDC5 18 0 0 5 23 (4.66) CDC6 2 0 1 14 17 (3.44) CDC7 0 0 0 1 1 (0.20) CDC8 3 0 0 0 3 (0.61) CDC9 0 0 0 1 1 (0.20) CDC10 1 0 0 2 3 (0.61) CDC11 0 0 0 1 1 (0.20) CDC12 2 0 0 1 3 (0.61) CDC13 0 0 0 1 1 (0.20) CDC14 0 0 0 1 1 (0.20) CDC15 0 0 0 1 1 (0.20) CDC16 0 0 0 1 1 (0.20) CDC17 0 0 0 1 1 (0.02) CDC18 0 0 0 1 1 (0.02) CDC19 0 0 0 1 1 (0.02) Note. LOS, lipooligosaccharide, GBS, Guillain-Barré syndrome. 
Figure 2. Newly confirmed LOS classes (CDC11–CDC19) OrthoMCL was used to cluster all LOS genes into orthologous and paralogous groups and visualized in Geneious software. Arrows represent orthologues and paralogues. Numbers are orthologue ID numbers, and “N” are paralogues. (Supplementary Table S3 available in www.besjournal.com). LOS, lipooligosaccharide.
Twenty-five LOS classes were identified in 260 strains from diarrhea patients. The five most common LOS classes were class B (28.08%, 73/260), A (12.69%, 33/260), H (9.23%, 24/260), P (8.08%, 21/260), and C (7.31%, 19/260). A total of 9 of 13 (69.23%) GBS-associated strains had class A LOS, and another 4 strains had LOS class P (2 strains), F (1 strain), and H (1 strain). Twelve LOS classes were identified in 70 livestock strains, and the five most abundant classes were class B (38.57%, 27/70), A (21.43%, 15/70), C (10%, 7/70), F (8.57%, 6/70), and S (4.29%, 3/70). Thirty-three LOS classes were identified in 151 strains from poultry. The five most common classes were class B (27.15%, 41/151), CDC6 (9.27%, 14/151), R (9.27%, 14/151), CDC1 (7.28%, 11/151), and W (6.62%, 10/151).
-
In Table 3, 17 CPS types of strains (strain number > 5) were analyzed. Strains of HS9, HS10, HS11, and HS37 were classified into LOS class CDC5, B, F, and P, respectively. And strains of the other 13 CPS types also had one or two prevalent LOS classes (bold in Table 3, P < 0.05). Four GBS-related genes (NeuA, NeuB, NseuC, and cst) involved in the sialic acid biosynthesis pathway were detected in 279 strains (56.48%, 279/494). In these strains, 14 CPS genotypes (HS1, HS2, HS4c, HS5/31, HS10, HS12, HS8/17, HS19, HS40, HS41, HS42, HS44, HS52, and HS60) were detected. The prevalence of GBS-related genes was higher (90%–100%) in strains HS1, HS2, HS10, HS19, and HS41 than the average (56.48%, P < 0.01). All strains of HS19 and HS41 contained GBS-related genes. Among HS19 strains, LOS class A was dominant (33/35), and the remaining 2 strains were classified into class R (1/35) and CDC11 (1/35). Strains of HS41 were classified into LOS class A (3/8), R (4/8), and CDC2 (1/8). LOS class A, R, CDC2, and CDC11 all possess GBS-related genes.
Table 3. Distribution characteristics of CPS/LOS types for 494 isolates of C. jejuni from China
CPS types Num LOS classes (Num) Proportion of strains
possessed sialic acid
synthesis genes in LOS (%)aHS1 21 B (8), C (10), F (1), V (1), CDC15 (1) 90.5 HS2 90 A (21), B (54), H (1), P (6), R (4), CDC6 (2), CDC8 (2) 90.0 HS4c 70 A (2), B (35), C (2), G (2), H (20), O (2), P (1), R (5), V (1) 64.3 HS5/31 40 A (2), B (24), D (1), F (3), K (6), V (1), CDC10 (3) 67.5 HS6/7 12 E (6), T (1), CDC1 (4), CDC5 (1) 0 HS8/17 28 B (1), C (17), G (7), V (1), CDC13 (1), CDC18 (1) 67.9 HS9 18 CDC5 (18) 0 HS10 11 B (11) 100 HS11 12 F (12) 0 HS12 21 W (8), CDC2 (13) 61.9 HS15 12 G (1), H (2), P (5), CDC3 (4) 0 HS18 8 T (1), CDC4 (5), CDC7 (1), CDC17 (1) 0 HS19 35 A (33), R (1), CDC11 (1) 100 HS21 4 W (4) 0 HS23/36 3 P (2), W (1) 0 HS32 3 CDC6 (3) 0 HS37 17 P (17) 0 HS40 3 B (1), H (1), M (1) 66.7 HS41 8 A (3), R (4), CDC2 (1) 100 HS42 3 C (1), K (1), W (1) 33.3 HS44 8 C (4), F (1), I (2), K (1) 50.0 HS52 2 B (1), CDC9 (1) / HS53 16 F (1), J (2), S (9), CDC5 (3), CDC6 (1) 0 HS55 1 E (1) / HS57 1 H (1) / HS60 1 CDC19 (1) / ND 46 B (6), C (2), G (2), H (6), P (2), R (1), CDC1 (9),
CDC5 (1), CDC6 (11), CDC8 (1),
CDC12 (3), CDC14 (1), CDC16 (1)21.7 In total 494 / 56.5 Note. aLOS classes A, B, C, M, R, V, CDC2, CDC8, CDC11, CDC15, CDC16, or CDC19. LOS: lipooligosaccharide; CPS: capsular polysaccharide. -
Among the tested 109 serotype known C. jejuni strains, all of the 11 HS2 strains, 9 HS19 strains and 8 HS41 strains were detected as positive, and 81 other serotype strains were all negative. The results were consistent with previously identified serotype results. The sensitivity and specificity of this multiplex real-time PCR assay were 100%, respectively.
-
Among the six HS19 strains, all had identical phenotypic serotypes. From the alignment, each of the CPS loci in the HS19 strain was 19 kb in length and contained 15 ORFs (Figure 3). The sequence identity of these six CPS loci was 98.2%. Taking the CPS locus sequences from strain HB_CJGB_ZHX (phenotypic serotype HS19) as the reference, the sequences of the CPS locus of strains HB_CJGB_LXC and HB-CJGB-ZB were 100% identical. However, 185 SNPs and one indel (insertion/deletion) were found in the CPS loci in strain NH_A12 (Figure 3 and Table 4), which had the same phenotypic serotype as strain HB_CJGB_ZHX. One SNP was found at the same location in ORF30 in each of the CPS loci in strains HBJ_CJGB_96G25 and BJ-CJGB96114. The CPS locus sequences of these two strains were 99.99% identical to the reference.
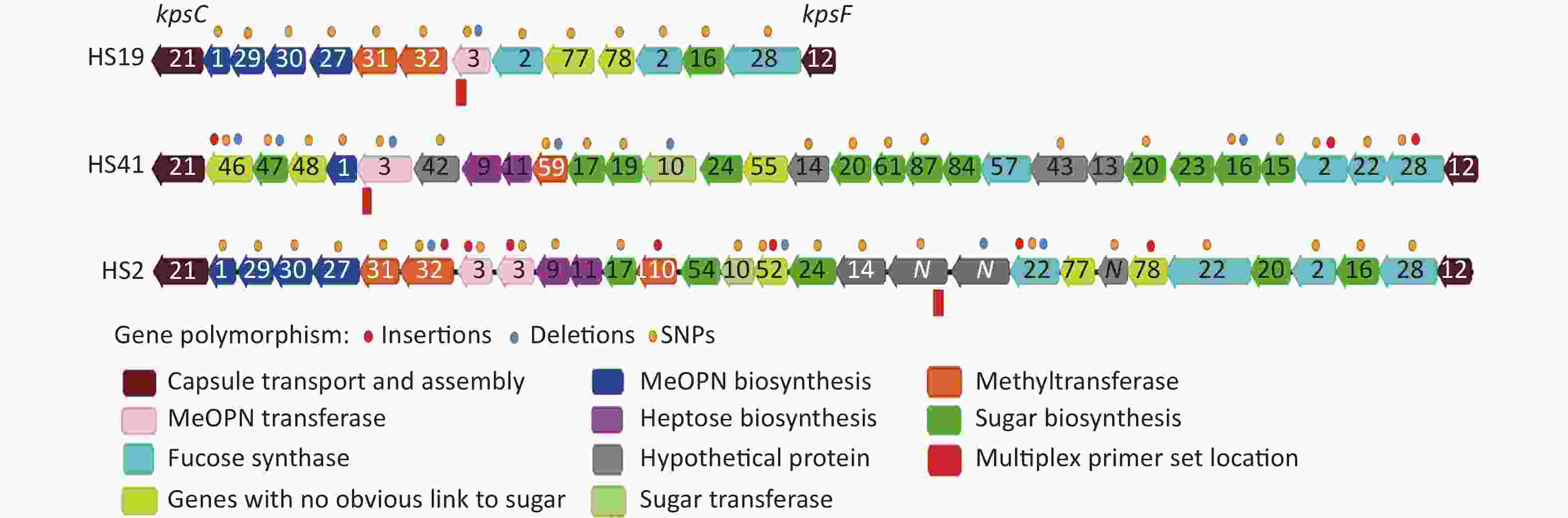
Figure 3. Schematic of CPS loci from HS2/HS19/HS41 and polymorphism distribution: OrthoMCL was used to cluster all CPS genes into orthologous and paralogous groups and visualized in Geneious software. Arrows represent orthologues and paralogues. Numbers are orthologue IDs, and “N” are paralogues. (Supplementary Table S4 available in www.besjournal.com) The different coloured dots represent gene polymorphism types. CPS: capsular polysaccharide.
Table 4. Genetic variation in the same CPS types
Strains CPS type Phenotypic serotype Sequence identity to reference (%) Polymorphisms in ORFs Sense mutations in ORFs HB_CJGB_LXC HS19 HS19 100 / / HB-CJGB-ZB HS19 HS19 100 / / HBJ_CJGB_96G25 HS19 HS19 99.99 1 SNP (30) / BJ-CJGB96114 HS19 HS19 99.99 1 SNP (30) / NH_A12 HS19 HS19 98.0 185 SNPs (1, 2, 3, 16, 27, 28, 29, 30, 31, 32, 77, 78 and a noncoding region); 1 deletion (3); 1 insertion (in a noncoding region) 56 substitutions (1, 2, 3, 16, 27, 28
, 29, 30, 31, 32, 78); 1 deletion (3)NH_F47 HS41 HS41 98.9 208 SNPs (1, 2, 3, 14, 15, 16, 17, 20, 22, 28, 42, 43, 46, 47, 48, 59, 61, 87 and three noncoding regions); 6 deletions (3, 10, 16, 46, 47, 59); 4 insertions (2, 28, 46, and one noncoding region) 99 substitutions (1, 2, 3, 14, 15, 16, 20, 22, 28, 42, 43, 46, 47, 48, 61, 87); 2 extensions (3, 46, 47); 7 frame shifts (3, 10, 16, 42, 46, 47, 59); 2 insertions (2, 28) ICDCCJ07004 HS41 NTa 99.9 11 SNPs (3, 42, 46, 61, 87 and a noncoding region); 1 insertion (46);
7 deletions (3, 10, 16, 46, 47, 59, 87)8 substitutions (3, 42, 46, 61, 87); 1 truncation (46); 2 extensions (3, 46); 7 frame shifts (3, 10, 16, 46, 47, 59, 87) Note. aThe phenotypic serotype could not be determined using a commercial kit. CPS: capsular polysaccharide. Two of the three HS41 strains had the same phenotypic serotypes (ICDCCJ07001 and NH_F47). The phenotypic serotypes of strain ICDCCJ07004 could not be determined using the same commercial phenotypic serotyping kit. The CPS locus of HS41 was 36 kb in length and contained 31 ORFs. Taking the CPS locus from ICDCCJ07001 as the reference, the sequence identity of these three CPS loci was 98.9% with the reference. In total, 208 SNPs and 6 indels (insertions/deletions) were found in NH_F47, and 11 SNPs and 8 indels were identified in ICDCCJ07004. Among the 11 HS2 strains, CPS loci were 37 kb in length and contained 30 ORFs. Taking the CPS locus from NCTC11168 as the reference, the sequence identity of 11 CPS gene clusters was 98.5%, and the polymorphisms among these strains included 212 SNPs and 18 indels. The genetic variations and schematics of the ORFs for each CPS type are presented in Figure 3 and Table 4.
-
In this study, we identified five major capsule types (> 5%): HS2, HS4c, HS5/31, HS19, and HS8/17 in 448 C. jejuni strains isolated from different sources in China. Forty-six strains could not be genotyped, possibly due to incomplete CPS loci or because their CPS types were not included in the database. Five capsule types (HS2, HS19, HS37, HS41, and HS53) were identified in 13 GBS-associated strains. Capsular-type HS19 was detected in 5.8% (35/259) of the enteritis strains but accounted for 42.9% (6/14) of the GBS-associated strains. Capsular-type HS41 was detected in 0.8% (2/259) of the enteritis strains but accounted for 14.3% (2/14) of the GBS-associated strains. We suggest that capsular types HS19 and HS41 might be associated with GBS.
In this study, we found that certain CPS types and LOS locus classes tended to appear in combination in C. jejuni strains. As described in Table 3, strains of most CPS types involved in this study had one or two prevalent LOS classes (Bold in Table 3); however, some CPS types could not be analyzed because the number of strains was too small (< 5). We speculate that certain combinations of CPS/LOS types might be related to the pathogenicity and environmental adaptation of C. jejuni.
The LOS classes A, B, C, M, R, V, CDC2, CDC8, CDC11, CDC15, CDC16, and CDC19 possess GBS-related genes (NeuA, NeuB, NeuC, and cst) involved in the sialic acid biosynthesis pathway. Approximately 90%–100% of strains of GBS-associated CPS types in this study (HS1, HS2, HS10, HS19, HS41) have LOS loci containing GBS-related genes, a prevalence that is much higher than the average (56.48%) (Table 3). All strains with capsular types HS19 and HS41 were classified into LOS classes A, R, CDC2, and CDC11. Interestingly, LOS classes A, R, CDC2, and CDC11 possess the sialylation genes necessary to produce ganglioside mimics. This ability may partly explain why HS19 and HS41 are the most prevalent serotypes among GBS-associated strains in China. All strains of capsular-type HS10 were identified as LOS class B, and HS10 was also reported as a Miller-Fisher syndrome-associated CPS type[7]; Miller-Fisher syndrome is a clinical variant of GBS.
A major weakness of this study is that our findings were not comprehensive. HS23/36 and HS37 were reported as GBS-associated serotypes, but strains of these capsular types do not possess sialylation genes. In this study, most strains of HS23/36 and HS37 were identified as LOS class P, which is a nonsialylated LOS with N-acetyl quinovosamine[27]. Further research is required to obtain evidence for the mechanism underlying the specific combination of capsular types and LOS classes resulting in synergistic effects on GBS risk.
The integrated sequence alignment revealed that the strains had identical CPS locus sequences and were classified into the same serotype (strains HB_CJGB_ZHX, HB_CJGB_LXC, HB-CJGB-ZB). Some strains had only a few SNPs and almost 99.99% similarity with the reference (strains HBJ_CJGB_96G25 and BJ-CJGB96114) and had the same phenotypic serotypes as the reference. Some other ORFs in the CPS locus had sense mutations, and the phenotypic serotype remained the same as the reference in strain NH_A12. We speculated that these mutations might not have affected the antigen structure of the CPS for the specific serotyping antibodies. For the same reason, the CPS locus in strain NH_F47 also had many SNPs, indels, extensions and frameshifts in the ORF regions, and none of these variations affected the phenotypic serotype. Eight nonsynonymous SNPs in ORFs 3/42/46/61/87, one truncation in ORF46, 7 frameshifts in ORFs 3/10/16/46/47/59/87, and two extensions in ORFs 3/46 were identified in the CPS locus from strain ICDCCJ07004. These mutations in the CPS locus caused a phenotypic variation in the serotype. Further investigation is needed to determine which mutation is crucial or causes the phase change in different serotypes.
The phenotypic serotyping method is time-consuming and labor-intensive. In this study, multiplex real-time PCR for the CPS types HS19, HS41, and HS2 was developed based on serotype-specific sequences. The multiplex PCR results were consistent with the phenotypic serotype results, which indicated the DNA variations in the CPS types HS19, HS41, and HS2 were not in the locations of the primers and probes. This approach offered a rapid screening strategy, which could not be affected by the expression of the CPS and focused on the rapid identification of the highly GBS-associated strains. However, the specific phenotypic serotypes of these strains need to be confirmed with specific serotypic antibodies.
-
The analysis of LOS and CPS typing characteristics contributes to recognizing the prevalence of virulence gene clusters of C. jejuni in China. The LOS class CDC11–CDC19 was first identified in this study. CDC11, CDC15, CDC16, and CDC19 possess genes for sialic acid synthesis and translocation, which are worthy of attention in strain monitoring. Strains of most capsular types had one or two prevalent LOS classes, supporting the hypothesis that capsular types and LOS locus classes tend to appear as a combination. The incidence of GBS is rare, and it is difficult to isolate Campylobacter strains once the neurological sign occurs; more GBS-related strains are needed for further confirmation study in the future. The multiplex real-time PCR developed in this study will be helpful for the identification of GBS-associated CPS types in China.
-
Table S3. The annotations of ORFs in novel LOS classes of C. jejuni
LOS class ORF Products (annotated by prokka) Length (bp) CDC11 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,071 39 hypothetical protein 780 7 hypothetical protein 933 37 hypothetical protein 929 41 N,N'-diacetyllegionaminic acid synthase 1,041 38 UDP-N-acetylglucosamine 2-epimerase 1,119 1 N-acylneuraminate cytidylyltransferase 666 53 2,3,4,5-tetrahydropyridine-2,6-dicarboxylate N-acetyltransferase 831 4 putative glycosyltransferase EpsJ 813 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC12 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 888 6 hypothetical protein 1,551 2 hypothetical protein 1,158 50 hypothetical protein 981 56 hypothetical protein 1,121 37 hypothetical protein 891 N hypothetical protein 489 N hypothetical protein 1,168 N hypothetical protein 782 N hypothetical protein 1,128 4 Putative glycosyltransferase EpsH 819 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC13 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,071 39 hypothetical protein 780 7 hypothetical protein 933 14 hypothetical protein 1,386 1 hypothetical protein 1,074 26 hypothetical protein 822 4 putative glycosyltransferase EpsJ 810 60 ADP-heptose--LPS heptosyltransferase 2 939 CDC14 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,071 39 hypothetical protein 780 7 hypothetical protein 936 2 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 1,584 26 hypothetical protein 2,022 4 putative glycosyltransferase EpsJ 810 60 ADP-heptose--LPS heptosyltransferase 2 939 CDC15 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,027 39 hypothetical protein 780 7 hypothetical protein 936 2 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 1,420 7 hypothetical protein 936 2 hypothetical protein 1,281 37 hypothetical protein 944 41 N,N'-diacetyllegionaminic acid synthase 1,041 38 UDP-N-acetylglucosamine 2-epimerase 1,119 1 N-acylneuraminate cytidylyltransferase 666 53 2,3,4,5-tetrahydropyridine-2,6-dicarboxylate N-acetyltransferase 755 4 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 758 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC16 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 2 hypothetical protein 1,224 N hypothetical protein 1,066 7 hypothetical protein 872 37 hypothetical protein 918 41 N,N'-diacetyllegionaminic acid synthase 1,032 38 UDP-N-acetylglucosamine 2-epimerase 1,116 1 hypothetical protein 954 1 N-acylneuraminate cytidylyltransferase 663 26 hypothetical protein 705 4 putative glycosyltransferase EpsJ 825 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC17 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 2 hypothetical protein 1,224 6 hypothetical protein 1,601 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,068 13 hypothetical protein 1,148 2 hypothetical protein 1,203 1 hypothetical protein 1,058 1 N-acylneuraminate cytidylyltransferase 663 26 hypothetical protein 843 4 putative glycosyltransferase EpsJ 825 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC18 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase811 39 hypothetical protein 966 7 hypothetical protein 936 26 hypothetical protein 891 4 putative glycosyltransferase EpsJ 810 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC19 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 2 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 1,275 6 hypothetical protein 1,560 37 hypothetical protein 935 41 N,N'-diacetyllegionaminic acid synthase 1,041 38 UDP-N-acetylglucosamine 2-epimerase 1,119 1 N-acylneuraminate cytidylyltransferase 798 53 Streptogramin A acetyltransferase 615 4 putative glycosyltransferase EpsJ 813 60 ADP-heptose--LPS heptosyltransferase 2 944 Table S4. The annotations of ORFs in CPS gene clusters of HS2, HS19, and HS41 of C. jejuni
CPS types ORF Product (annotated by prokka) Length (bp) HS2 21 Capsule polysaccharide modification protein 2,067 1 adenylyl-sulfate kinase 513 29 sugar nucleotidyltransferase 762 30 amidotransferase 603 27 hypothetical protein 2,340 31 methyltransferase 762 32 methyltransferase 774 3 sugar transferase 1,839 3 sugar transferase 1,878 9 D-glycero-D-manno-heptose 1-phosphate guanosyltransferase 666 11 phosphoheptose isomerase 606 17 D-glycero-D-manno-heptose 7-phosphate kinase 1,020 110 methyltransferase family protein 852 54 sugar-nucleotide epimerase/dehydratease 942 10 GDP-L-fucose synthetase 1,041 52 hypothetical protein 927 24 dTDP-4-dehydrorhamnose 3,5-epimerase 546 14 capsular polysaccharide heptosyltransferase 1,749 N sugar transferase 3,096 N hypothetical protein 1,107 22 sugar transferase 1,335 77 phosphatase 636 N aminotransferase 1,230 78 aminotransferase 1,104 22 sugar transferase 2,328 20 UDP-galactopyranose mutase 1,107 2 sugar transferase 1,224 16 UDP-glucose 6-dehydrogenase 1,182 28 sugar transferase 1,635 12 D-arabinose 5-phosphate isomerase 495 HS19 21 Capsule polysaccharide modification protein 2,067 1 adenylyl-sulfate kinase 513 29 sugar nucleotidyltransferase 762 30 amidotransferase 603 27 hypothetical protein 2,340 31 methyltransferase 762 32 methyltransferase 774 3 sugar transferase 1,833 2 hypothetical protein 1,509 77 phosphatase 633 78 aminotransferase 1,077 2 hypothetical protein 2,499 16 UDP-glucose 6-dehydrogenase 1,176 28 hypothetical protein 2,124 12 D-arabinose 5-phosphate isomerase 495 HS41 21 Capsule polysaccharide modification protein 2,067 46 Na(+)/H(+)-K(+) antiporter GerN 1,254 47 pyruvate kinase 960 48 sulfate adenylyltransferase 1,047 1 adenylyl-sulfate kinase 534 3 sugar transferase 1,854 42 hypothetical protein 1,749 9 D-glycero-D-manno-heptose 1-phosphate guanosyltransferase 666 11 phosphoheptose isomerase 606 59 hypothetical protein 1,236 17 D-glycero-D-manno-heptose 7-phosphate kinase 1,020 19 NAD-dependent 4,6-dehydratase 1,032 10 GDP-L-fucose synthase 2,214 24 dTDP-4-dehydrorhamnose 3,5-epimerase 546 55 hypothetical protein 1,251 14 capsular polysaccharide heptosyltransferase 1,608 20 UDP-galactopyranose mutase 1,131 61 NAD-dependent 4,6-dehydratase 885 87 dTDP-4-dehydrorhamnose 3,5-epimerase 630 84 NAD-dependent 4,6-dehydratase 1,071 57 Glucose-1-phosphate cytidylyltransferase 801 43 hypothetical protein 2,157 13 hypothetical protein 1,380 20 UDP-galactopyranose mutase 1,170 23 UDP-GlcNAc/Glc 4-epimerase 975 16 UDP-glucose 6-dehydrogenase 1,299 15 NAD-dependent 4,6-dehydratase 957 2 hypothetical protein 1,371 22 sugar transferase 1,050 28 sugar transferase 1,632 12 D-arabinose 5-phosphate isomerase 495
doi: 10.3967/bes2022.140
Genetic Characteristics of Lipooligosaccharide and Capsular Polysaccharide of Campylobacter jejuni from Different Sources in China
-
Abstract:
Objective To determine the distribution of two important virulence factors [lipooligosaccharide (LOS) and capsular polysaccharide (CPS)] in Campylobacter jejuni (C. jejuni) isolated from different sources in China and to develop a rapid screening method for Guillain–Barré syndrome (GBS)-associated strains. Methods Whole-genome sequencing was carried out for 494 C. jejuni strains. The OrthoMCL software was used to define the LOS/CPS gene clusters. CPS genotyping was performed with serotype-specific sequence alignment using the BLAST software. Real-time Polymerase chain reaction (PCR) was developed with the unique sequences of specific CPS types. Results Nine novel and 29 previously confirmed LOS classes were identified. LOS classes A, B, and C were the most common (48.2%, 238/494) among the 494 strains. Twenty-six capsular types were identified in 448 strains. HS2, HS4c, HS5/31, HS19, and HS8/17 were the most frequent CPS genotypes (58.7%, 263/448). Strains of 17 CPS genotypes (strain number > 5) had one or two prevalent LOS classes (P < 0.05). Multiplex real-time PCR for rapid identification of HS2, HS19, and HS41 was developed and validated with strains of known serotypes. Conclusion Our results describe the genetic characteristics of the important virulence factors in C. jejuni strains in China. The multiplex real-time PCR developed in this study will facilitate enhanced surveillance of GBS-associated strains in China. -
Key words:
- Campylobacter jejuni /
- Lipooligosaccharide /
- Capsular polysaccharide /
- Multiplex real-time PCR
The authors declare no conflicts of interest.
Availability: Shell scripts for identifying specific serotype sequences are available from the authors upon request.
注释:1) AUTHORS’ CONTRIBUTIONS: 2) CONFLICTS OF INTEREST: 3) CONFLICTS OF INTEREST: -
Figure 1. Distribution of different CPS types in strains from different sources: The CPS genotype for each strain was determined based on the different serospecific sequences. The top five CPS types were HS2 (90 strains, 20.1%), HS4c (70 strains, 15.6%), HS5/31 (40 strains, 8.9%), HS19 (35 strains, 7.8%), and HS8/17 (28 strains, 6.3%), which accounted for 58.7% (263/448) of the tested strains. CPS, capsular polysaccharide
Figure 2. Newly confirmed LOS classes (CDC11–CDC19) OrthoMCL was used to cluster all LOS genes into orthologous and paralogous groups and visualized in Geneious software. Arrows represent orthologues and paralogues. Numbers are orthologue ID numbers, and “N” are paralogues. (Supplementary Table S3 available in www.besjournal.com). LOS, lipooligosaccharide.
Figure 3. Schematic of CPS loci from HS2/HS19/HS41 and polymorphism distribution: OrthoMCL was used to cluster all CPS genes into orthologous and paralogous groups and visualized in Geneious software. Arrows represent orthologues and paralogues. Numbers are orthologue IDs, and “N” are paralogues. (Supplementary Table S4 available in www.besjournal.com) The different coloured dots represent gene polymorphism types. CPS: capsular polysaccharide.
S1. The accession numbers for CPS gene clusters of C. jejuni
CPS types Accession number HS1 BX545859 HS2 AL111168.1 HS3 HQ343268 HS4c HQ343269 HS5/31 KT868847 HS6/S7 NC_009839 HS8/17 HQ343270 HS9 KT868844 HS10 HQ343271 HS11 KT868845 HS12 KT868848 HS15/31/58 HQ343272 HS18 KT932997 HS19 BX545860 HS21 KT868849 HS22 KT893439 HS23/36 BX545858 HS27 KT893437 HS29 KT868846 HS32 KT893435 HS33/35 KT893436 HS37 KT893431 HS38 KT893430 HS40 KT893434 HS41 BX545857 HS42 HQ343274 HS44 JF496678 HS45/5/32/60 KT893432 HS52 KT893429 HS53 CP000025.1 HS55 KT893433 HS57 KT893428 HS58/32 KT893427 HS60 KT893426 HS63 KT893438  下载: 导出CSV
下载: 导出CSV
S2. The accession numbers for LOS gene clusters of C. jejuni
LOS class Accession number A AF215659 B AF401528 C AF400047 D AF400669 E AJ131360 F AY423554 G AY436358 H AY800272 I EU404107 J EU404104 K AY573819 L EU404111 M EF140720 N AY816330 O EF143352 P AY943308 Q EU404112 R AY962325 S EU404110 T AIOC00000000 U AIOU00000000 V AIOP00000000 W CP001900 CDC1-10 reference[24] CDC11-19 In this study
下载: 导出CSV
Table 1. Primers and probes for real-time PCR detection of HS2, HS19, and HS41
Capsular type Primers and probes Nucleotide sequence (5’ to 3’) HS2 HS2-F AACCAACCATCCACTATTTTCATCT HS2-R ACTCTTTGATTTTCCAATGCAATGTT HS2-P FAM-CGCACCTTCCAATGCAACCAAAGAGC-BHQ1 HS19 HS19-F CTGAAATATCAACACAGGGAAAATG HS19-R TGTTGGAGGTAAAAGACATGGTG HS19-P HEX-TCCAATCCTGTTGCTCATAACGCATTCA-BHQ1 HS41 HS41-F CACCCTGATTTCTATTAACTCTACCAC HS41-R GAGTTATTCTTGATGCTGAAAATGGA HS41-P CY5-AAACGGGATAAGATCGCTTGGGGGAATA-BHQ2 Note. PCR, Polymerase chain reaction.
下载: 导出CSV
Table 2. The distribution of the LOS classes in strains from different sources
LOS class Diarrhoea
patientsGBS
patientsLivestock Poultry In total
(%)A 33 9 15 4 61 (12.35) B 73 0 27 41 141 (28.54) C 19 0 7 10 36 (7.29) D 0 0 0 1 1 (0.20) E 3 0 3 1 7 (1.42) F 9 1 6 2 18 (3.64) G 10 0 0 2 12 (2.43) H 24 1 2 4 31 (6.28) I 0 0 2 0 2 (0.40) J 2 0 0 0 2 (0.40) K 5 0 0 3 8 (1.62) M 0 0 0 1 1 (0.20) O 2 0 0 0 2 (0.40) P 21 2 2 8 33 (6.68) R 1 0 0 14 15 (3.04) S 5 0 3 1 9 (1.82) T 1 0 0 1 2 (0.40) V 1 0 1 2 4 (0.81) W 4 0 0 10 14 (2.83) CDC1 1 0 1 11 13 (2.63) CDC2 12 0 0 2 14 (2.83) CDC3 3 0 0 1 4 (0.81) CDC4 5 0 0 0 5 (1.01) CDC5 18 0 0 5 23 (4.66) CDC6 2 0 1 14 17 (3.44) CDC7 0 0 0 1 1 (0.20) CDC8 3 0 0 0 3 (0.61) CDC9 0 0 0 1 1 (0.20) CDC10 1 0 0 2 3 (0.61) CDC11 0 0 0 1 1 (0.20) CDC12 2 0 0 1 3 (0.61) CDC13 0 0 0 1 1 (0.20) CDC14 0 0 0 1 1 (0.20) CDC15 0 0 0 1 1 (0.20) CDC16 0 0 0 1 1 (0.20) CDC17 0 0 0 1 1 (0.02) CDC18 0 0 0 1 1 (0.02) CDC19 0 0 0 1 1 (0.02) Note. LOS, lipooligosaccharide, GBS, Guillain-Barré syndrome.
下载: 导出CSV
Table 3. Distribution characteristics of CPS/LOS types for 494 isolates of C. jejuni from China
CPS types Num LOS classes (Num) Proportion of strains
possessed sialic acid
synthesis genes in LOS (%)aHS1 21 B (8), C (10), F (1), V (1), CDC15 (1) 90.5 HS2 90 A (21), B (54), H (1), P (6), R (4), CDC6 (2), CDC8 (2) 90.0 HS4c 70 A (2), B (35), C (2), G (2), H (20), O (2), P (1), R (5), V (1) 64.3 HS5/31 40 A (2), B (24), D (1), F (3), K (6), V (1), CDC10 (3) 67.5 HS6/7 12 E (6), T (1), CDC1 (4), CDC5 (1) 0 HS8/17 28 B (1), C (17), G (7), V (1), CDC13 (1), CDC18 (1) 67.9 HS9 18 CDC5 (18) 0 HS10 11 B (11) 100 HS11 12 F (12) 0 HS12 21 W (8), CDC2 (13) 61.9 HS15 12 G (1), H (2), P (5), CDC3 (4) 0 HS18 8 T (1), CDC4 (5), CDC7 (1), CDC17 (1) 0 HS19 35 A (33), R (1), CDC11 (1) 100 HS21 4 W (4) 0 HS23/36 3 P (2), W (1) 0 HS32 3 CDC6 (3) 0 HS37 17 P (17) 0 HS40 3 B (1), H (1), M (1) 66.7 HS41 8 A (3), R (4), CDC2 (1) 100 HS42 3 C (1), K (1), W (1) 33.3 HS44 8 C (4), F (1), I (2), K (1) 50.0 HS52 2 B (1), CDC9 (1) / HS53 16 F (1), J (2), S (9), CDC5 (3), CDC6 (1) 0 HS55 1 E (1) / HS57 1 H (1) / HS60 1 CDC19 (1) / ND 46 B (6), C (2), G (2), H (6), P (2), R (1), CDC1 (9),
CDC5 (1), CDC6 (11), CDC8 (1),
CDC12 (3), CDC14 (1), CDC16 (1)21.7 In total 494 / 56.5 Note. aLOS classes A, B, C, M, R, V, CDC2, CDC8, CDC11, CDC15, CDC16, or CDC19. LOS: lipooligosaccharide; CPS: capsular polysaccharide.
下载: 导出CSV
Table 4. Genetic variation in the same CPS types
Strains CPS type Phenotypic serotype Sequence identity to reference (%) Polymorphisms in ORFs Sense mutations in ORFs HB_CJGB_LXC HS19 HS19 100 / / HB-CJGB-ZB HS19 HS19 100 / / HBJ_CJGB_96G25 HS19 HS19 99.99 1 SNP (30) / BJ-CJGB96114 HS19 HS19 99.99 1 SNP (30) / NH_A12 HS19 HS19 98.0 185 SNPs (1, 2, 3, 16, 27, 28, 29, 30, 31, 32, 77, 78 and a noncoding region); 1 deletion (3); 1 insertion (in a noncoding region) 56 substitutions (1, 2, 3, 16, 27, 28
, 29, 30, 31, 32, 78); 1 deletion (3)NH_F47 HS41 HS41 98.9 208 SNPs (1, 2, 3, 14, 15, 16, 17, 20, 22, 28, 42, 43, 46, 47, 48, 59, 61, 87 and three noncoding regions); 6 deletions (3, 10, 16, 46, 47, 59); 4 insertions (2, 28, 46, and one noncoding region) 99 substitutions (1, 2, 3, 14, 15, 16, 20, 22, 28, 42, 43, 46, 47, 48, 61, 87); 2 extensions (3, 46, 47); 7 frame shifts (3, 10, 16, 42, 46, 47, 59); 2 insertions (2, 28) ICDCCJ07004 HS41 NTa 99.9 11 SNPs (3, 42, 46, 61, 87 and a noncoding region); 1 insertion (46);
7 deletions (3, 10, 16, 46, 47, 59, 87)8 substitutions (3, 42, 46, 61, 87); 1 truncation (46); 2 extensions (3, 46); 7 frame shifts (3, 10, 16, 46, 47, 59, 87) Note. aThe phenotypic serotype could not be determined using a commercial kit. CPS: capsular polysaccharide.
下载: 导出CSV
S3. The annotations of ORFs in novel LOS classes of C. jejuni
LOS class ORF Products (annotated by prokka) Length (bp) CDC11 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,071 39 hypothetical protein 780 7 hypothetical protein 933 37 hypothetical protein 929 41 N,N'-diacetyllegionaminic acid synthase 1,041 38 UDP-N-acetylglucosamine 2-epimerase 1,119 1 N-acylneuraminate cytidylyltransferase 666 53 2,3,4,5-tetrahydropyridine-2,6-dicarboxylate N-acetyltransferase 831 4 putative glycosyltransferase EpsJ 813 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC12 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 888 6 hypothetical protein 1,551 2 hypothetical protein 1,158 50 hypothetical protein 981 56 hypothetical protein 1,121 37 hypothetical protein 891 N hypothetical protein 489 N hypothetical protein 1,168 N hypothetical protein 782 N hypothetical protein 1,128 4 Putative glycosyltransferase EpsH 819 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC13 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,071 39 hypothetical protein 780 7 hypothetical protein 933 14 hypothetical protein 1,386 1 hypothetical protein 1,074 26 hypothetical protein 822 4 putative glycosyltransferase EpsJ 810 60 ADP-heptose--LPS heptosyltransferase 2 939 CDC14 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,071 39 hypothetical protein 780 7 hypothetical protein 936 2 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 1,584 26 hypothetical protein 2,022 4 putative glycosyltransferase EpsJ 810 60 ADP-heptose--LPS heptosyltransferase 2 939 CDC15 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,027 39 hypothetical protein 780 7 hypothetical protein 936 2 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 1,420 7 hypothetical protein 936 2 hypothetical protein 1,281 37 hypothetical protein 944 41 N,N'-diacetyllegionaminic acid synthase 1,041 38 UDP-N-acetylglucosamine 2-epimerase 1,119 1 N-acylneuraminate cytidylyltransferase 666 53 2,3,4,5-tetrahydropyridine-2,6-dicarboxylate N-acetyltransferase 755 4 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 758 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC16 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 2 hypothetical protein 1,224 N hypothetical protein 1,066 7 hypothetical protein 872 37 hypothetical protein 918 41 N,N'-diacetyllegionaminic acid synthase 1,032 38 UDP-N-acetylglucosamine 2-epimerase 1,116 1 hypothetical protein 954 1 N-acylneuraminate cytidylyltransferase 663 26 hypothetical protein 705 4 putative glycosyltransferase EpsJ 825 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC17 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 2 hypothetical protein 1,224 6 hypothetical protein 1,601 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase1,068 13 hypothetical protein 1,148 2 hypothetical protein 1,203 1 hypothetical protein 1,058 1 N-acylneuraminate cytidylyltransferase 663 26 hypothetical protein 843 4 putative glycosyltransferase EpsJ 825 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC18 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 6 hypothetical protein 702 18 GalNAc-alpha-(1->4)-GalNAc-alpha-(1->3)- diNAcBac-PP-undecaprenol
alpha-1,4-N-acetyl-D-galactosaminyltransferase811 39 hypothetical protein 966 7 hypothetical protein 936 26 hypothetical protein 891 4 putative glycosyltransferase EpsJ 810 60 ADP-heptose--LPS heptosyltransferase 2 944 CDC19 8 Lipopolysaccharide heptosyltransferase 1 1,029 5 Lipid A biosynthesis lauroyltransferase 879 2 Undecaprenyl-phosphate 4-deoxy-4-formamido-L-arabinose transferase 1,275 6 hypothetical protein 1,560 37 hypothetical protein 935 41 N,N'-diacetyllegionaminic acid synthase 1,041 38 UDP-N-acetylglucosamine 2-epimerase 1,119 1 N-acylneuraminate cytidylyltransferase 798 53 Streptogramin A acetyltransferase 615 4 putative glycosyltransferase EpsJ 813 60 ADP-heptose--LPS heptosyltransferase 2 944
下载: 导出CSV
S4. The annotations of ORFs in CPS gene clusters of HS2, HS19, and HS41 of C. jejuni
CPS types ORF Product (annotated by prokka) Length (bp) HS2 21 Capsule polysaccharide modification protein 2,067 1 adenylyl-sulfate kinase 513 29 sugar nucleotidyltransferase 762 30 amidotransferase 603 27 hypothetical protein 2,340 31 methyltransferase 762 32 methyltransferase 774 3 sugar transferase 1,839 3 sugar transferase 1,878 9 D-glycero-D-manno-heptose 1-phosphate guanosyltransferase 666 11 phosphoheptose isomerase 606 17 D-glycero-D-manno-heptose 7-phosphate kinase 1,020 110 methyltransferase family protein 852 54 sugar-nucleotide epimerase/dehydratease 942 10 GDP-L-fucose synthetase 1,041 52 hypothetical protein 927 24 dTDP-4-dehydrorhamnose 3,5-epimerase 546 14 capsular polysaccharide heptosyltransferase 1,749 N sugar transferase 3,096 N hypothetical protein 1,107 22 sugar transferase 1,335 77 phosphatase 636 N aminotransferase 1,230 78 aminotransferase 1,104 22 sugar transferase 2,328 20 UDP-galactopyranose mutase 1,107 2 sugar transferase 1,224 16 UDP-glucose 6-dehydrogenase 1,182 28 sugar transferase 1,635 12 D-arabinose 5-phosphate isomerase 495 HS19 21 Capsule polysaccharide modification protein 2,067 1 adenylyl-sulfate kinase 513 29 sugar nucleotidyltransferase 762 30 amidotransferase 603 27 hypothetical protein 2,340 31 methyltransferase 762 32 methyltransferase 774 3 sugar transferase 1,833 2 hypothetical protein 1,509 77 phosphatase 633 78 aminotransferase 1,077 2 hypothetical protein 2,499 16 UDP-glucose 6-dehydrogenase 1,176 28 hypothetical protein 2,124 12 D-arabinose 5-phosphate isomerase 495 HS41 21 Capsule polysaccharide modification protein 2,067 46 Na(+)/H(+)-K(+) antiporter GerN 1,254 47 pyruvate kinase 960 48 sulfate adenylyltransferase 1,047 1 adenylyl-sulfate kinase 534 3 sugar transferase 1,854 42 hypothetical protein 1,749 9 D-glycero-D-manno-heptose 1-phosphate guanosyltransferase 666 11 phosphoheptose isomerase 606 59 hypothetical protein 1,236 17 D-glycero-D-manno-heptose 7-phosphate kinase 1,020 19 NAD-dependent 4,6-dehydratase 1,032 10 GDP-L-fucose synthase 2,214 24 dTDP-4-dehydrorhamnose 3,5-epimerase 546 55 hypothetical protein 1,251 14 capsular polysaccharide heptosyltransferase 1,608 20 UDP-galactopyranose mutase 1,131 61 NAD-dependent 4,6-dehydratase 885 87 dTDP-4-dehydrorhamnose 3,5-epimerase 630 84 NAD-dependent 4,6-dehydratase 1,071 57 Glucose-1-phosphate cytidylyltransferase 801 43 hypothetical protein 2,157 13 hypothetical protein 1,380 20 UDP-galactopyranose mutase 1,170 23 UDP-GlcNAc/Glc 4-epimerase 975 16 UDP-glucose 6-dehydrogenase 1,299 15 NAD-dependent 4,6-dehydratase 957 2 hypothetical protein 1,371 22 sugar transferase 1,050 28 sugar transferase 1,632 12 D-arabinose 5-phosphate isomerase 495
下载: 导出CSV
-
[1] Kaakoush NO, Castaño-Rodríguez N, Mitchell HM, et al. Global epidemiology of Campylobacter infection. Clin Microbiol Rev, 2015; 28, 687−720. doi: 10.1128/CMR.00006-15 [2] Koga M, Gilbert M, Takahashi M, et al. Comprehensive analysis of bacterial risk factors for the development of Guillain-Barre syndrome after Campylobacter jejuni enteritis. J Infect Dis, 2006; 193, 547−55. doi: 10.1086/499969 [3] Godschalk PCR, Heikema AP, Gilbert M, et al. The crucial role of Campylobacter jejuni genes in anti-ganglioside antibody induction in Guillain-Barré syndrome. J Clin Invest, 2004; 114, 1659−65. doi: 10.1172/JCI200415707 [4] Yuki N. Campylobacter sialyltransferase gene polymorphism directs clinical features of Guillain-Barré syndrome. J Neurochem, 2007; 103 Suppl 1, 150−8. [5] Karlyshev AV, Linton D, Gregson NA, et al. Genetic and biochemical evidence of a Campylobacter jejuni capsular polysaccharide that accounts for Penner serotype specificity. Mol Microbiol, 2000; 35, 529−41. [6] Heikema AP, Islam Z, Horst-Kreft D, et al. Campylobacter jejuni capsular genotypes are related to Guillain-Barré syndrome. Clin Microbiol Infect, 2015; 21, 852. e1−9. [7] Salloway S, Mermel LA, Seamans M, et al. Miller-Fisher syndrome associated with Campylobacter jejuni bearing lipopolysaccharide molecules that mimic human ganglioside GD3. Infect Immun, 1996; 64, 2945−9. doi: 10.1128/iai.64.8.2945-2949.1996 [8] Takahashi M, Koga M, Yokoyama K, et al. Epidemiology of Campylobacter jejuni isolated from patients with Guillain-Barre and Fisher syndromes in Japan. J Clin Microbiol, 2005; 43, 335−9. doi: 10.1128/JCM.43.1.335-339.2005 [9] Zhang MJ, Li Q, He LH, et al. Association study between an outbreak of Guillain-Barre syndrome in Jilin, China, and preceding Campylobacter jejuni infection. Foodborne Pathog Dis, 2010; 7, 913−9. doi: 10.1089/fpd.2009.0493 [10] Zhang MJ, He LH, Li Q, et al. Genomic characterization of the Guillain-Barre syndrome-associated Campylobacter jejuni ICDCCJ07001 Isolate. PLoS One, 2010; 5, e15060. doi: 10.1371/journal.pone.0015060 [11] Yuki N. Molecular mimicry between gangliosides and lipopolysaccharides of Campylobacter jejuni isolated from patients with Guillain-Barré syndrome and Miller Fisher syndrome. J Infect Dis, 1997; 176 Suppl 2, S150−3. [12] Yuki N, Takahashi M, Tagawa Y, et al. Association of Campylobacter jejuni serotype with antiganglioside antibody in Guillain-Barré syndrome and Fisher's syndrome. Ann Neurol, 1997; 42, 28−33. doi: 10.1002/ana.410420107 [13] Liang H, Wen ZY, Li Y, et al. Comparison of the filtration culture and multiple real-time PCR examination for Campylobacter spp. from stool specimens in diarrheal patients. Front Microbiol, 2018; 9, 2995. doi: 10.3389/fmicb.2018.02995 [14] Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics, 2014; 30, 2068−9. doi: 10.1093/bioinformatics/btu153 [15] Pruitt KD, Tatusova T, Brown GR, et al. NCBI Reference Sequences (RefSeq): current status, new features and genome annotation policy. Nucleic Acids Res, 2012; 40, D130−5. doi: 10.1093/nar/gkr1079 [16] Karlyshev AV, Champion OL, Churcher C, et al. Analysis of Campylobacter jejuni capsular loci reveals multiple mechanisms for the generation of structural diversity and the ability to form complex heptoses. Mol Microbiol, 2005; 55, 90−103. [17] Klena JD, Gray SA, Konkel ME. Cloning, sequencing, and characterization of the lipopolysaccharide biosynthetic enzyme heptosyltransferase I gene (waaC) from Campylobacter jejuni and Campylobacter coli. Gene, 1998; 222, 177−85. doi: 10.1016/S0378-1119(98)00501-0 [18] Li L, Stoeckert CJ Jr, Roos DS. OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res, 2003; 13, 2178−89. doi: 10.1101/gr.1224503 [19] Richards VP, Lefébure T, Pavinski Bitar PD, et al. Comparative characterization of the virulence gene clusters (lipooligosaccharide and capsular polysaccharide) for Campylobacter coli, Campylobacter jejuni subsp. jejuni and related Campylobacter species. Infect Genet Evol, 2013; 14, 200−13. doi: 10.1016/j.meegid.2012.12.010 [20] Kuzniar A, van Ham RCHJ, Pongor S, et al. The quest for orthologs: finding the corresponding gene across genomes. Trends Genet, 2008; 24, 539−51. doi: 10.1016/j.tig.2008.08.009 [21] Li H, Handsaker B, Wysoker A, et al. The sequence alignment/map format and SAMtools. Bioinformatics, 2009; 25, 2078−9. doi: 10.1093/bioinformatics/btp352 [22] Li H. Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. Preprint at arXiv https://arxiv.org/abs/1303.3997, 2013. [2022-3-12]. [23] Li DH, Liu CM, Luo RB, et al. MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics, 2015; 31, 1674−6. doi: 10.1093/bioinformatics/btv033 [24] Wang JQ, Gu YX, Zhou GL, et al. Genetic diversity of lipooligosaccharide core biosynthesis gene clusters in Campylobacter jejuni. Dis Surveill, 2020; 35, 11−5. (In Chinese [25] Gilbert M, Karwaski MF, Bernatchez S, et al. The genetic bases for the variation in the lipo-oligosaccharide of the mucosal pathogen, Campylobacter jejuni: biosynthesis of sialylated ganglioside mimics in the core oligosaccharide. J Biol Chem, 2002; 277, 327−37. doi: 10.1074/jbc.M108452200 [26] Parker CT, Gilbert M, Yuki N, et al. Characterization of lipooligosaccharide-biosynthetic loci of Campylobacter jejuni reveals new lipooligosaccharide classes: evidence of mosaic organizations. J Bacteriol, 2008; 190, 5681−9. doi: 10.1128/JB.00254-08 [27] Poly F, Read TD, Chen YH, et al. Characterization of two Campylobacter jejuni strains for use in volunteer experimental-infection studies. Infect Immun, 2008; 76, 5655−67. doi: 10.1128/IAI.00780-08 -
 22159Supplementary Materials.pdf
22159Supplementary Materials.pdf

-

 点击查看大图
点击查看大图
计量
- 文章访问数: 601
- HTML全文浏览量: 267
- PDF下载量: 73
- 被引次数: 0

 Quick Links
Quick Links