
-
With about one in ten cancer cases and fatalities, CRC is the third most common malignancy in the world and the second dominant cause of cancer-associated deaths[1]. Progress has been made in the fight against CRC over the past few decades, such as the widespread application of screening tests, and the development of immunotherapy, yet, CRC-related mortality and morbidity are continuously rising. Today, anticancer treatments concentrating on particular molecular targets have garnered the most interest. Clarifying the molecular pathways behind CRC occurrence and progression is therefore particularly significant.
As a form of non-coding single-stranded RNA with an average length of 20–22 nucleotides, microRNA (miRNA) is well known to target 3'-untranslated region (3'-UTR) in target mRNAs to silence gene expression at post-transcriptional levels[2]. Multiple reports have also indicated the capability of miRNAs to target protein-coding sequences (CDS)[3-5]. Depending on which potential oncogenes or tumor repressors they particularly target, miRNAs may functionally promote or repress a variety of cancers[6]. They are proven to be dysregulated in diverse cancer types including CRC, which shapes an advantageous microenvironment conducive to tumor progression[7,8]. MiR-224-5p is overexpressed and further involved in various kinds of tumorigenic pathways, according to several studies[9,10]. However, microRNA expression is known to be highly tissue-specific and time-specific[11]. It is yet unclear how miR-224-5p regulates colorectal cancer. Our previous research demonstrated that miR-224-5p facilitates CRC invasion and migration via targeting Beta-Transducin Repeat Containing E3 Ubiquitin Protein Ligase (BTRC)[12]. In this study, we verified ULK2 as another target gene of miR-224-5p and further discussed their effect on CRC tumor cell proliferation.
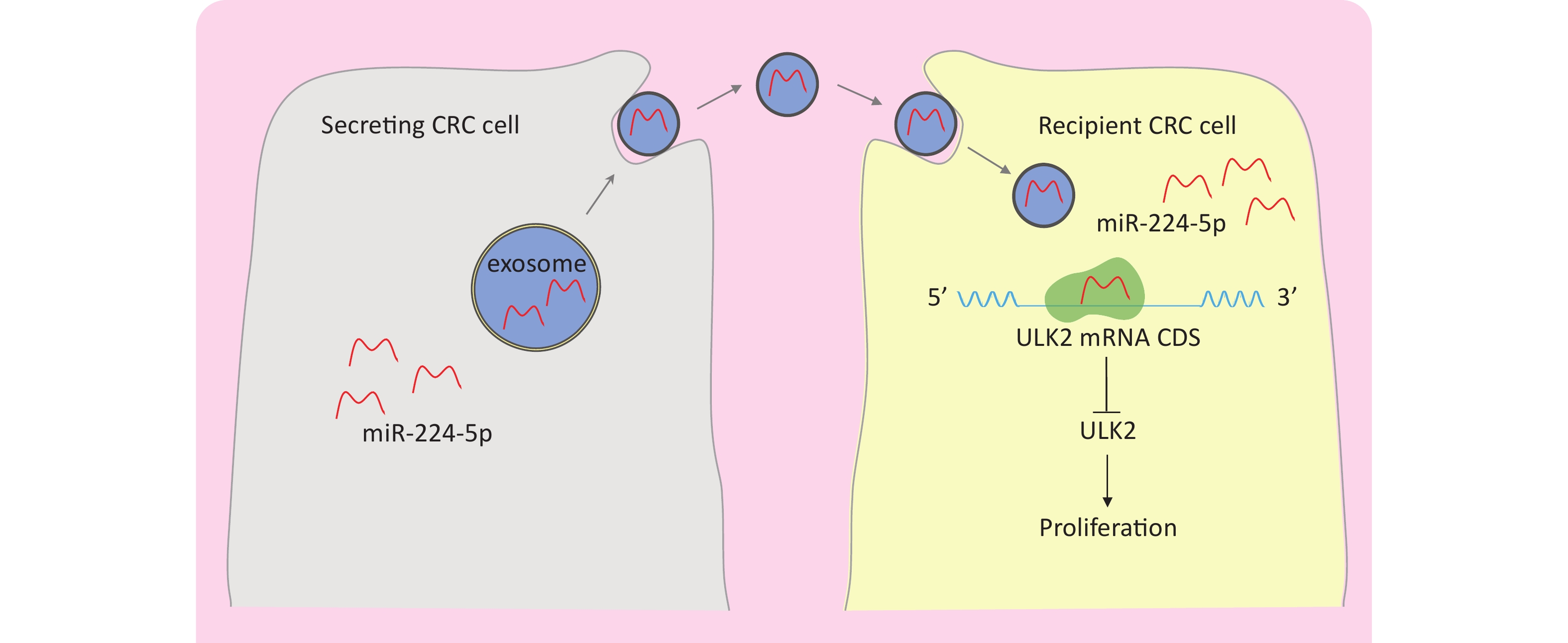
It is widely known that miRNAs primarily act within the cytoplasm. Intriguingly, research has demonstrated that they are also exported to extracellular space via exosomes released by practically all cell types[13]. The exosome is a kind of membranous vesicle with a lipid bilayer that ranges in size from 30 to 100 nm and contains cell-specific assemblages of bioactive molecules comprising proteins, lipids, and nucleic acids[14]. Cancer-derived exosomes contain a lot of abnormal miRNAs as well as their own mechanisms related to miRNA regulation[15]. They can be released from donor cells, then transmit their encapsulated specific miRNAs to nearby or distant cells and regulate target gene expression, thereby playing critical roles in proliferation, angiogenesis, and metastasis[16-18]. However, to date, it has not been reported whether miR-224-5p may be secreted into the tumor microenvironment through CRC-derived exosomes, then transferred to adjacent cells to directly interact with which target genes and play which regulatory roles.
It has been reported that about 40%–50% of sporadic CRC has p53 mutation, especially p53 R273H exists a high mutation rate and p53 mutation status is strongly correlated with the progression and outcome of CRC[19]. This arouses our great curiosity. However, to yet, whether the regulatory effect of exosomal miR-224-5p on CRC is related to p53 mutation still remains to be answered.
Overall, this paper makes a preliminary exploration for the above pending problems. We highlighted for the first time that CRC cell-derived miR-224-5p can be released into the tumor microenvironment via exosomes and consequently transferred to recipient cells in paracrine or autocrine form, regulating cell proliferation in a p53-dependent manner via directly targeting ULK2. This discovery may provide more effective targets for CRC patients and lay a solid theoretical foundation for developing novel treatment strategies.
-
The Shanghai Huashan Hospital affiliated with Fudan University offered us 5 pairs of paraffin tissues (carcinoma and corresponding para-carcinoma) from CRC patients. Based on our previous research[20], we selected 177 laser capture microdissected snap-frozen colorectal tissues from 66 CRC and 5 adenoma patients for this study. The samples included 71 normal tissues, 18 inflammatory polyps, 22 adenomas, and 66 carcinomas (15 Dukes’ A, 21 Dukes’ B, 19 Dukes’ C, 11 Dukes’ D). Among the 22 adenomas, seventeen were from patients who also had CRC. The study received approval from the Fudan University Basic Medical School Ethics Committee (approval no.2019-C018), which followed the principles of the Declaration of Helsinki. All patients were informed and provided written consent. Table 1 and Supplementary Table S1 (available in www.besjournal.com) provide details on CRC samples involved in this study.
Characteristics Case (n) Sex Male 41 Female 30 Age (y) ≤ 60 31 > 60 40 Tumor position Colon 42 Rectum 25 Colorectal junction 4 Tumor Size ≤ 3 cm 29 > 3 cm 42 Tumor stage Dukes’ A 15 Dukes’ B 21 Dukes’ C 19 Dukes’ D 11 Adenoma High-grade 2 Low-grade 3 p53 status Negative 18 Positive 40 Unknown 13 Lymphatic invasion No 46 Yes 25 Ki67 ≤ 50% 23 > 50% 35 Unknown 13 Table 1. The clinical-pathological features of 71 patients for microarray
-
The complete cell culture medium was made by combining 10% exosome-free FBS (System Biosciences, USA), 1% penicillin-streptomycin (Invitrogen, USA), and DMEM (Gibco, USA). The human embryonic kidney cell lines (HEK293T) and human colorectal cancer cell lines (HCT116, DLD1, SW480, HT29) purchased from the Chinese Academy of Sciences (Shanghai, China) were cultured at 37 °C in an incubator with 5% CO2 (volume fraction) and saturated humidity.
-
Total RNA was extracted from cells and exosomes in accordance with the guideline of miRNeasy Micro Kit (Qiagen, Germany). NanoDrop 2000 spectrophotometer (Thermo, USA) was utilized to detect RNA concentration. MiR-224-5p expression was determined by miRNA-specific primers (Ribobio, Guangzhou) and Mir-X miRNA qRT-PCR TB Green Kit (Takara, Japan) with U6 expression as its reference gene. ULK2 mRNA expression was measured by ChamQ Universal SYBR qPCR Master Mix and HiScript III RT SuperMix (Vazyme, Nanjing) with GAPDH expression as its reference gene. Supplementary Table S2 (available in www.besjournal.com) provides a list of the primer sequences for SYBR qRT-PCR. The expression difference between the control group and the experimental group was assessed using the 2-ΔΔCt method.
Target Primer sequences GAPDH (human) Forward: 5’-CTGACTTCAACAGCGACACC-3’ Reverse: 5’-TGCTGTAGCCAAATTCGTTGT-3’ ULK2 (human) Forward: 5’-ACAGCAAAGGAATCATCCACAG-3’ Reverse: 5’-TGATGCGAATACCACTGACAC-3’ p53 (human) Forward: 5’-AATCTACTGGGACGGAACAGC-3’ Reverse: 5’-CCAAGACTTAGTACCTGAAGGGTG-3’ U6 (human) Forward: 5’-GGAACGATACAGAGAAGATTAGC-3’ Reverse: 5’-TGGAACGCTTCACGAATTTGCG-3’ miR-224-5p (human) Forward: produced by Ribobio Reverse: Universal reverse sequence of the kit Table S2. The primer sequences used in this study
-
According to our previous study[12], Genewiz (China) constructed pmirGLO ULK2 CDS-MUT (mutant) and pmirGLO ULK2 CDS-WT (wild type) using pmirGLO vector (Promega, USA), which had been digested at XbaI and PmeI and then integrated with the seed sequence in the CDS of ULK2 as well as the matching wild seed nucleotides. Supplementary Table S3 (available in www.besjournal.com) lists the sequences that we put into pmirGLO to reconstruct plasmid.
Names of inserts Inserted sequences (5’→3’) ULK2 CDS-WT GTTTAAACCGTCCGCTGTGTGCCCCGGGGGCGCGGCCATGGAGGTGGTGG GTGACTT CGAGTACAGCAAGAGGGATCTCGTGGGACACGGGGCCTTCGTCTAGAULK2 CDS-MUT GTTTAAACCGTCCGCTGTGTGCCCCGGGGGCGCGGCCATGGAGGTGGTGG GCATGCG CGAGTACAGCAAGAGGGATCTCGTGGGACACGGGGCCTTCGTCTAGANote. Restriction sites are in underline; Wild-type or mutant seed regions are in red. Table S3. The sequences inserted in the pmirGLO to reconstruct ULK2 CDS-WT or ULK2 CDS-MUT plasmid
-
According to Lipofectamine 2000 instructions (Life Technologies, USA), CRC cells were transfected with 100 nmol/L miR-224-5p inhibitor/50 nmol/L miR-224-5p mimic, or their matching NC (RiboBio, Guangzhou). At 48 h after transfection, the protein or RNA level was determined by western blot or qRT-PCR, and the succeeding tests were finished within 72 h. HEK293T cells in 48-well plates were co-transfected with 5 ng pmirGLO ULK2 CDS-MUT or pmirGLO ULK2 CDS -WT and 20 nmol/L miR-224-5p mimic or NC, respectively.
-
Dual-Luciferase Reporter Gene Assay Kit (Yeasen, China) was applied to HEK293T cells after co-transfection for 48 h, and the results were assessed using GloMax® 20/20 Luminometer (Promega, USA). The activity of firefly luciferase was used to standardize the activity of renilla luciferase.
-
Harvested cells were washed two times with PBS at 1,000 rpm for 5 min. Then, 200 μL PBS was added to resuspend the cells, and the suspension is subsequently added drop by drop into a centrifuge tube containing 1 mL 70% cold ethanol to fix cells overnight at –20 °C. After being washed with PBS, the cells were resuspended in 200 μL PBS and then darkly stained for 15 min after adding 500 μL PI/RNase Staining Buffer. Finally, a flow cytometer was used to examine the red fluorescence at 488 nm excitation wavelength within 1 h.
-
Centrifugation was used to separate cells for 5 min at 1,000 rpm, followed by two PBS washes. After that, they were resuspended with 500 μL 1×Binding Buffer. For fluorescent labeling, 100 μL of cell suspension per well was mixed with 5 μL PI and 5 μL Annexin V-APC (BD Biosciences, USA). Upon gentle mixing, the cells were placed away from light for 15 min at room temperature. Finally, a flow cytometer was used to measure apoptosis within 1 h.
-
The total protein was lysed using RIPA lysis buffer (Thermo, USA), then quantified by the bicinchoninic acid protein assay (Thermo, USA). The following antibodies’ protein expressions were examined: ULK2 (1:2000, abs139410, Absin, China), p53 (1:2000, Sc-126, Santa Cruz Biotechnology), and GAPDH (1:2000, Sc-166574, Santa Cruz Biotechnology) as the endogenous reference. Image J was used to calculate western blot bands’ gray values.
-
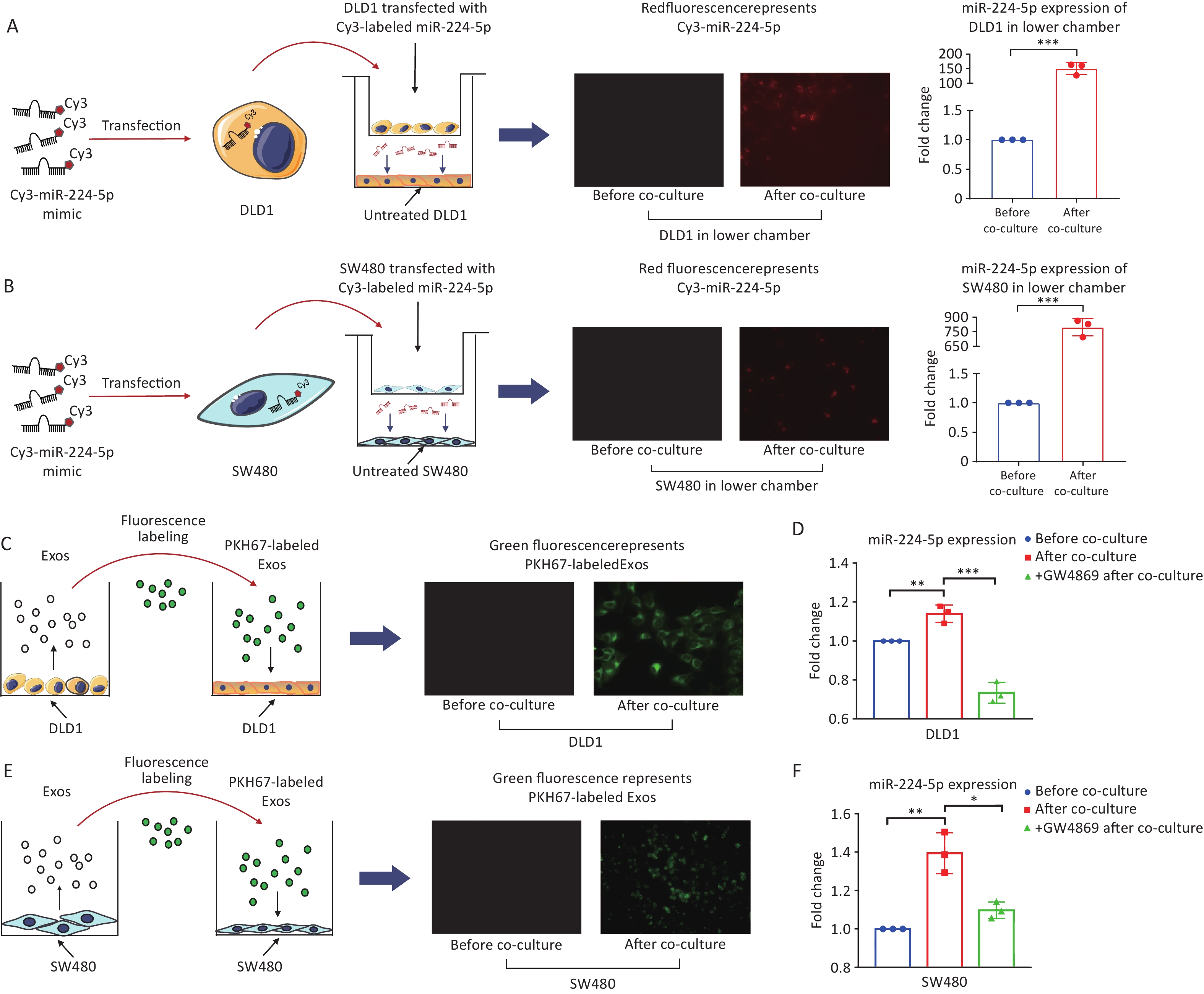
The CRC cells (donor cells) were inoculated into the upper chamber of a transwell plate (0.4 μm polycarbonate filter, Corning) at a density of 70% per well after being transfected with Cy3-miR-224-5p mimic for 48 h, while the conventionally cultured CRC cells (recipient cells) were inoculated into the lower chamber at a density of 50% per well 12–24 h in advance. After 24 h of incubation, recipient cells were washed twice with PBS, then observed under the ECLIPSE TE2000U (Nikon, Japan) for the presence of Cy3 red fluorescence. Donor cells were pretreated for 24 h with 10 μmol/L GW4869 (MedChemExpress, USA), a neutral sphingomyelinase inhibitor that can inhibit exosome secretion[21]. They were then co-cultured with recipient cells for a further 24 h in the culture medium added with GW4869. qRT-PCR was conducted to examine miR-224-5p expression in recipient cells.
-
To begin, we inoculated equivalent cells into 10 cm plates (Corning, USA). When the cell density reaches 80%, we switched the culture media to one without FBS after twice-repeated PBS washing. After 48 h, exosomes from the cell culture supernatant were extracted in accordance with the instructions of ExoQuick extraction solution (System Biosciences, USA). The collected exosomes were immediately applied in subsequent assays.
-
Exosomes were labeled using the PKH67 fluorescent cell linker kit (Sigma-Aldrich), then they were extracted using ExoQuick extraction solution after being washed with PBS. Subsequently, PKH67-labeled exosomes were added to a 12-well plate of recipient CRC cells and incubated for 4 h. Finally, three PBS washes of the cells were followed by fluorescence microscopy analysis (Nikon, Japan).
-
2 × 103 cells were inoculated into a 96-well plate with 100 μL medium per well, and the plate was incubated for the specified durations. Cell viability was separately evaluated by Cell Counting Kit-8 assay (CCK-8, Yeasen, China) at 12 h and 48 h after inoculation. Finally, the absorbance at 450 nm per well was detected by a Multiskan GO analyzer (Thermo, USA). All experiments were conducted in triplicate, with each group containing six multiple wells. Cell viability (%) = [(ODtreatment - ODblank)48 h - (ODtreatment - ODblank)12 h]/[(ODcontrol - ODblank)48 h-(ODcontrol - ODblank)12 h] × 100.
-
The target plasmids involved in this study were supplied by Shanghai Jikai Gene. The transfer vector and two packaging vectors including psPAX2 and pMD2.G were transfected into HEK293T cells for 48 h to generate lentiviral particles. Subsequently, the collected lentiviral particles were purified by 0.45 μm pore filters (Millipore, USA). For transduction, CRC cells at 70% density were inoculated in a 60mm dish and transduced with the lentivirus. The ULK2 protein level was detected by western blot assays.
-
The EdU assay was conducted with a BeyoClick EdU-488 Cell Proliferation Kit (Beyotime, Shanghai) in line with the manufacturer’s directions. Briefly, the cells transfected with miRNAs or fed with exosomes were inoculated on cell slides pre-placed in 12-well plates. After that, they sequentially underwent a 2-h incubation with 50 μmol/L EdU, a 30-min fixation with 4% paraformaldehyde, a 10-min permeabilization with 0.5% Triton X-100, and a 30-min incubation with the click reaction mixture. Finally, the slides were treated with Hoechst dye and imaged with AXIO Imager. A2 microscope (Zeiss, Germany). Following the random selection of three fields, EdU-positive cells (green-stained nuclei) were counted. The ratio of EdU positive cell numbers to Hoechst positive cell numbers is the EdU labeling rate. The experiment was carried out three times.
-
The miR-224-5p expression data were extracted from 627 samples of The Cancer Genome Atlas (TCGA) database, consisting of 11 normal samples and 616 CRC samples. Referring to our previous research[12], candidate targets of miR-224-5p were predicted via at least 4 out of 11 databases from miRecords (http://c1.accurascience.com/miRecords/).Mapping the candidate targets to cancer-associated pathways was completed by the Kyoto Encyclopedia of Genes and Genomes (KEGG) database (http://www.kegg.jp/kegg/download/kegtools.html). The GEPIA (http://gepia.cancer-pku.cn) was used to access the TCGA database and collect information about ULK2 transcripts in colon and rectum adenocarcinomas (COAD and READ). Every parameter was set to default.
-
After being fixed with 10% formalin and paraffin-embedded, primary tumor tissues were divided into slices with a thickness of about 4 μm. Then, tissue sections were incubated with anti-ULK2 antibody (1:100, Novus, USA) after blocking the activation of endogenous peroxidase, followed by DAB substrate staining and hematoxylin counterstaining. Finally, representative images were captured by a digitalized microscope camera. Statistical analysis of relative gray values of normal tissues and CRC tissues was performed by Image J, using ULK2 positive dots in the stroma as control and normalized to the normal tissues.
-
177 snap-frozen tissues were obtained by LCM and further used for microarray analysis. Following Arcturus's (Mountain View, CA) suggested procedure, the target colorectal epithelial cells (–2 × 105) were chosen and captured by ultraviolet laser cutting for RNA isolation. According to the manufacturer's protocol (Agilent Technologies, USA), 100 ng total RNA was then labeled and hybridized by microarray comprising probes for 723 human microRNAs from the Sanger database V.10.1. Please refer to our previous report for detailed methods of LCM and microarray analysis[22]. miR-224-5p chip data are shown in Supplemental File 2.
-
Cell/Tissue DNA Isolation Mini Kit (Vazyme, Nanjing) was used to extract genomic DNA from cells. The PCR amplification system for DNA was described below: DNA template 0.2 μg, 2×Phanta Max Master Mix (Vazyme, Nanjing) 25 μL, 10 μmol/L Forward primer (Sangon Biotech, Shanghai) 2 μL, 10 μmol/L Reverse primer (Sangon Biotech, Shanghai) 2 μL, RNase-Free ddH2O (Vazyme, Nanjing); the S1000 Thermal Cycle PCR instrument (Bio-Rad, USA) was used for the reaction under the conditions of 95 °C for 3 min, 95 °C for 15 s, 64 °C for 15 s, 72 °C for 30 s, 34 cycles; following 72 °C for 5 min. DNA agarose gel electrophoresis was used to validate amplification results. The target DNA fragment was recovered as manufacturer’s directions using DNA Gel Extraction Kit (Beyotime, China), and then the recovered DNA products were used for direct Sanger sequencing (Genewiz, China). DNA sequencing results were compared with target gene sequences using Nucleotide Blast Tool in NCBI.
Supplementary Table S2 shows the PCR amplification primer sequences. The results of DNA sequencing are shown inSupplementary Table S4 , available in www.besjournal.com. -
Unless otherwise noted, the data were given as mean ± standard deviation (SD). One-way analysis of variance (ANOVA) with Tukey’s post-hoc tests was used to investigate differences across numerous groups, while two-tailed Student’s t-tests were used to evaluate differences between two groups. Unpaired Student’s t-test with Benjaminie-Hochberg correction was used to assess the miR-224-5p expressions from TCGA. A statistical difference was considered to exist when P < 0.05. GraphPad Prism 7.0 (GraphPad Software, CA) was used to conduct each statistical analysis. Statistical details and P values of experiments can be found in the corresponding figure legends. FlowJo 10.0 was used to analyze the data of flow cytometry. At least three separate biological replicates were performed in every experiment.
-
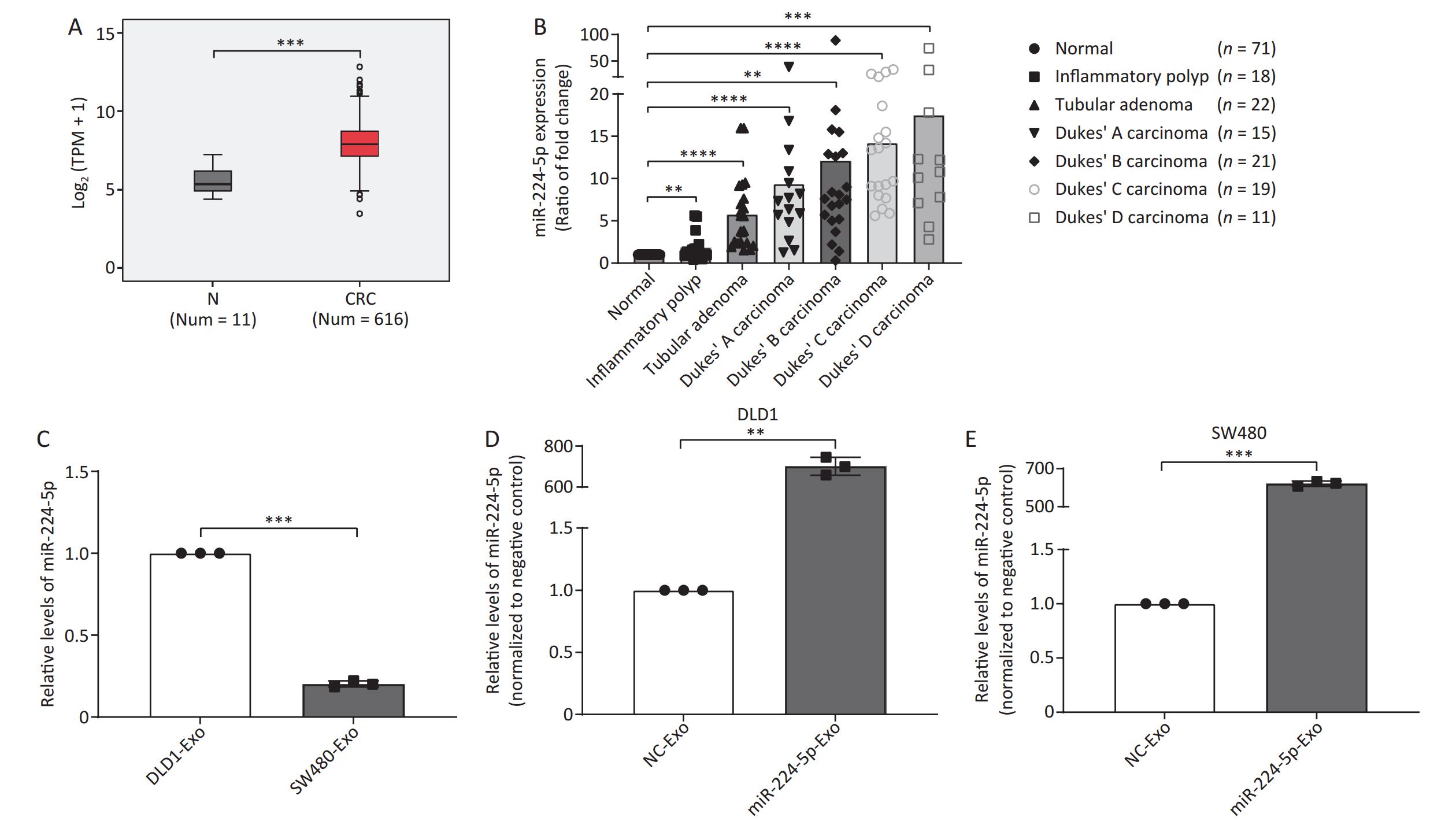
In this study, we analyzed miR-224-5p expression level through The Cancer Genome Atla (TCGA) database. The findings showed that miR-224-5p was noticeably upregulated in CRC tissues as compared to para-carcinoma tissues (Figure 1A). Meanwhile, we used LCM to obtain colorectal epithelial cells from human normal and colorectal tumor tissues and then extracted total RNA for miRNA microarray analysis. The results suggested that the miR-224-5p expression level in tubular adenoma tissues was remarkably higher than that in normal tissues at the surgical margin (P < 0.0001, Figure 1B). Additionally, we found during the evolution from adenoma tissues to colorectal cancer tissues, miR-224-5p expression progressively elevated compared to normal tissues (Dukes’ A: P < 0.0001, Dukes’ B: P = 0.0022, Dukes’ C: P < 0.0001, Dukes’ D: P = 0.0001, Figure 1B). Whereas, the miR-224-5p expression level in these patient tissues was not correlated with sex, age, tumor position, tumor size, and lymphatic invasion (Supplementary Figure S1, available in www.besjournal.com).
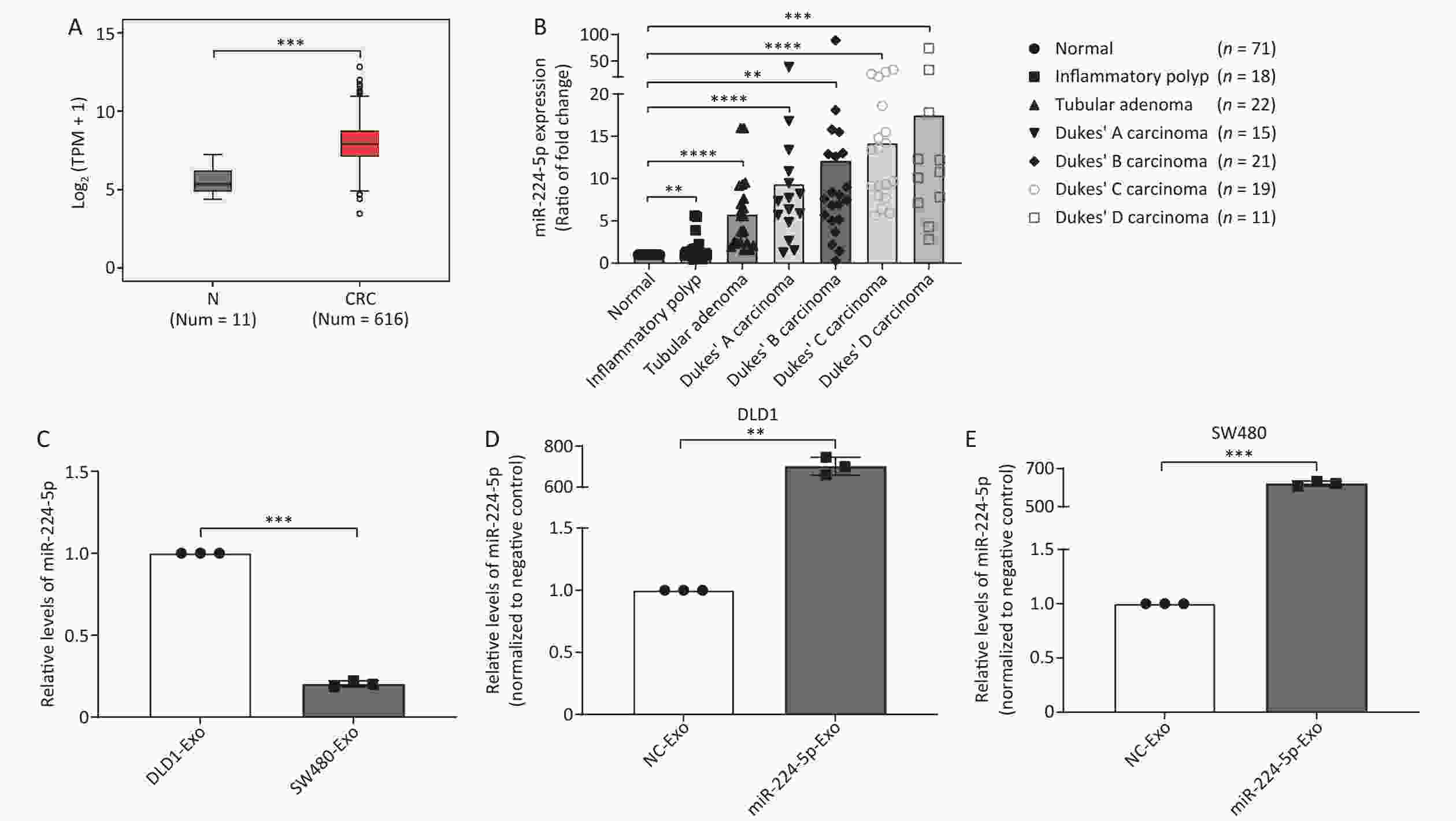
Figure 1. miR-224-5p is upregulated in CRC tissues and CRC cell-derived exosomes. (A) Expression of miR-224-5p in normal and CRC tissues. N, Normal; TPM, transcripts per million; CRC, colorectalcancer. (B) The ratio (mean) of miR-224-5p signals in different colorectal lesions normalized with normal tissues. (C) qRT-PCR evaluation of miR-224-5p expression in DLD1 and SW480 cell-derived exosomes. (D, E) qRT-PCR evaluation of miR-224-5p expression in exosomes derived from DLD1 (D) and SW480 (E) cells transfected with miR-224-5p mimic or miR-NC. Data are mean ± SD (n ≥ 3). **P < 0.01, ***P < 0.001, ****P < 0.0001.
Currently, studies show that exosomes from human CRC cells have been successfully isolated and identified by a precipitation method using Exoquick precipitation solution[23]. To determine whether upregulated-miR-224-5p contributes to the development of CRC via exosomes, we further extracted exosomes from DLD1 and SW480 cell media according to the widely-used method abovementioned. As shown in Figure 1C, miR-224-5p expression in SW480 cell-secreted exosomes was considerably lower than that in exosomes secreted by DLD1 cells (P = 0.0002), suggesting that the expression difference of exosomal miR-224-5p between CRC cell lines was consistent with cellular miR-224-5p expression from our previous experimental results[12]. Exosomal miR-224-5p expression levels in CRC cells that had been transfected with miR-224-5p mimic were significantly higher than those in the control group (P = 0.0013, P = 0.0002, Figure 1D and 1E). Collectively, these findings suggested that CRC cells may be capable of secreting extracellular miR-224-5p via exosomes, and overexpressed exosomal miR-224-5p may exert an impact on CRC phenotypes.
-
We first determined whether CRC cells can secret miR-224-5p and be transported to target cells. After 24h of co-culture, Cy3-labeled miR-224-5p mimic produced by CRC cells in the upper chamber was able to be detected in the untreated CRC cells in the lower chamber of transwell plate (Figure 2A and 2B). At the same time, the miR-224-5p expression level was markedly increased in the lower chamber CRC cells (P = 0.0002, P = 0.0001, Figure 2A and 2B). As a result, CRC cells can secrete extracellular miR-224-5p and target neighboring tumor cells.
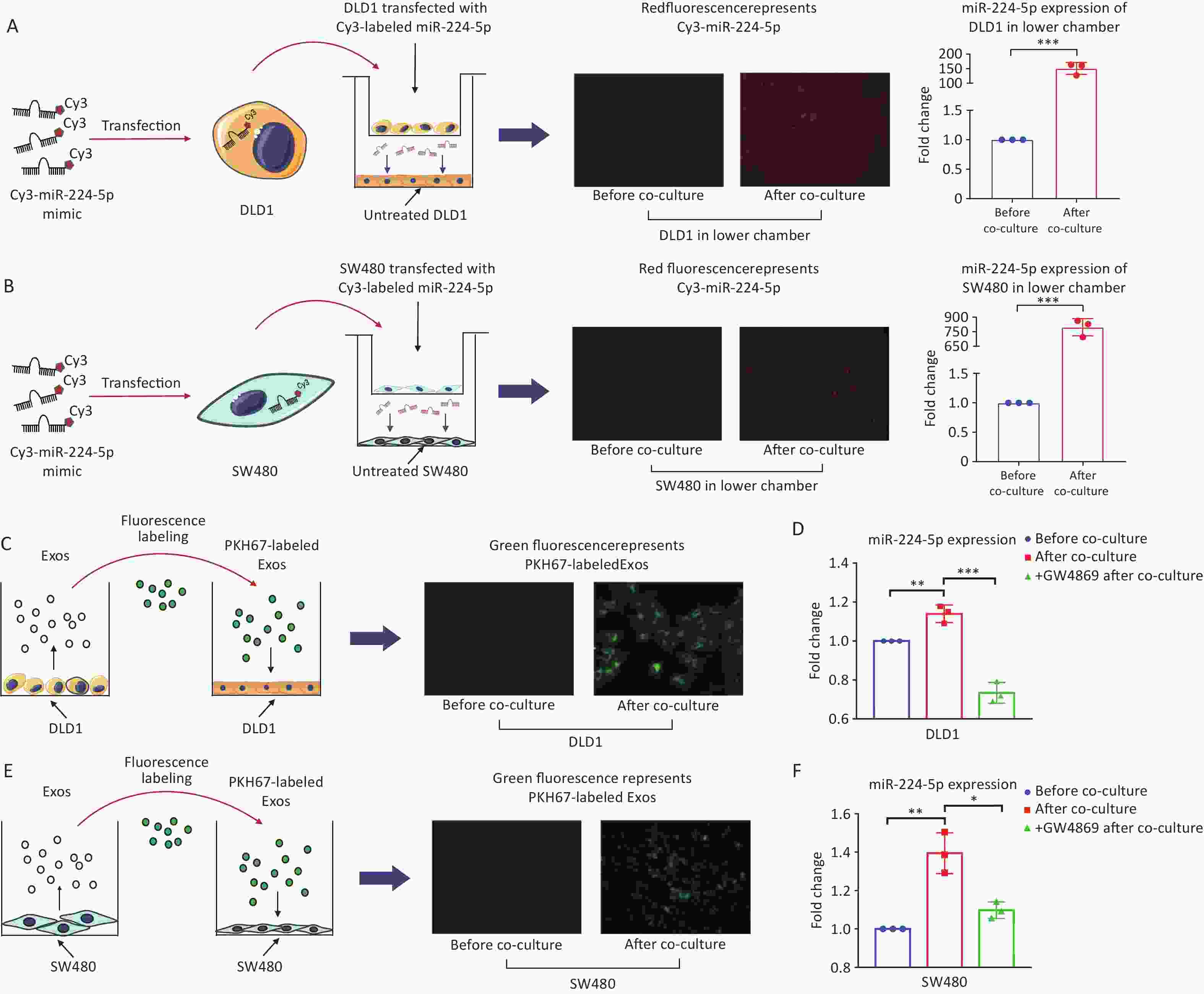
Figure 2. CRC cells secrete exosomal miR-224-5p and target adjacent tumor cells. (A, B) Cy3-labeled miR-224-5p mimic-transfected CRC cells were co-cultured with untreated DLD1 cells (A) and SW480 cells (B) (20×). (C) PKH67-labeled Exosomes from DLD1 cells were added to DLD1 cell cultures (40×). (D, F) Effects of GW4689 on exosome-dependent miR-224-5p transfer from DLD1 (D) and SW480 cells (F). (E) PKH67-labeled Exosomes from SW480 cells were added to SW480 cell cultures (20×). Data are mean ± SD (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001.
Secondly, we investigated whether exosomal miR-224-5p (exo-miR-224-5p in short) secreted by CRC cells can also target neighboring tumor cells. After adding CRC cell-derived exosomes labeled with PKH67 dye, green fluorescence was observed in recipient CRC cells (Figure 2C and 2E). In addition, after co-culture, lower chamber CRC cells showed a substantially fold increase in miR-224-5p expression (Figure 2D and 2F). More significantly, prior addition of the exosome secretion inhibitor GW4869 into the upper chamber obstructed exosome production and miR-224-5p transfer (Figure 2D and 2F). Briefly, these results show that exo-miR-224-5p can be secreted by donor CRC cells, then efficiently target recipient cells.
-
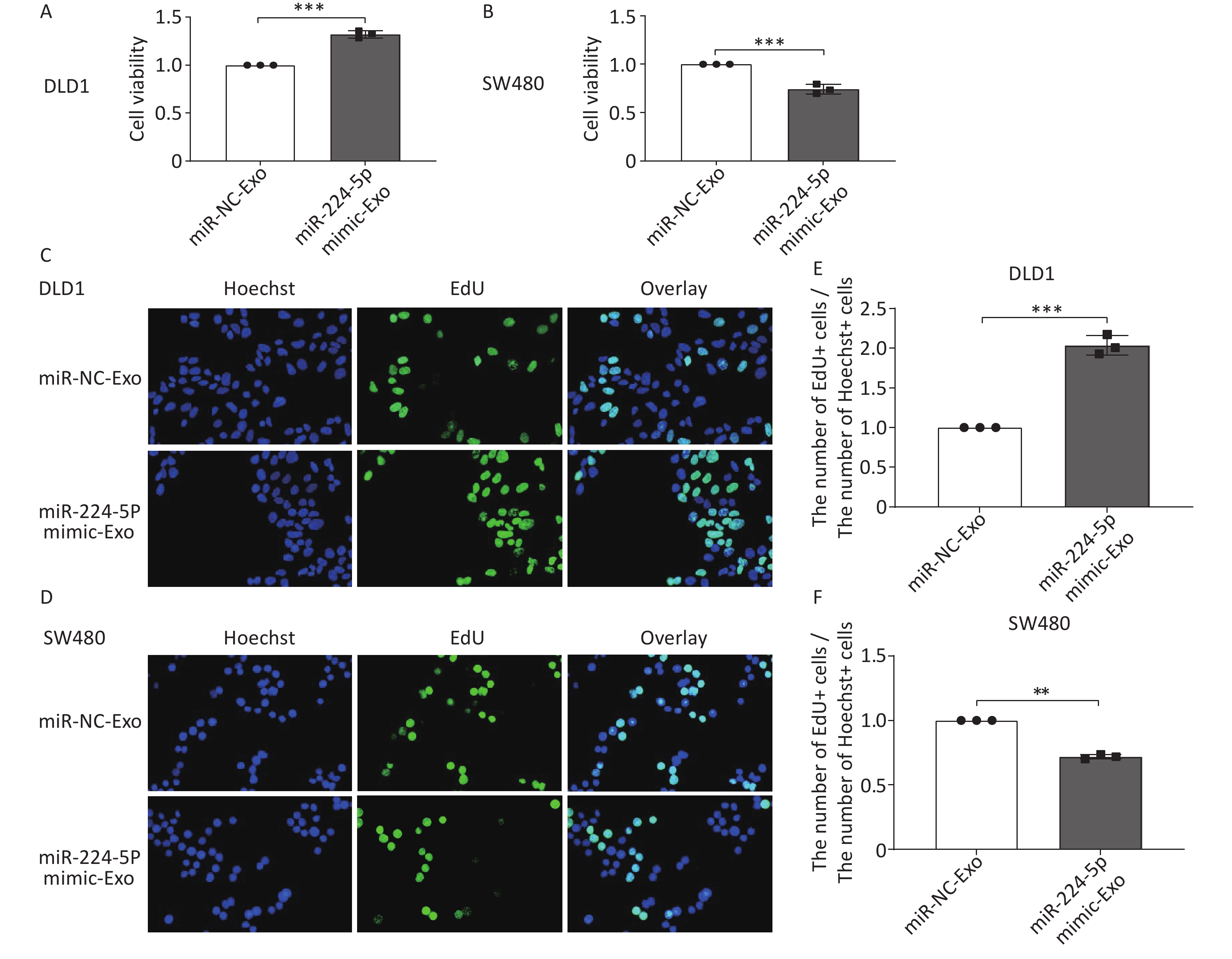
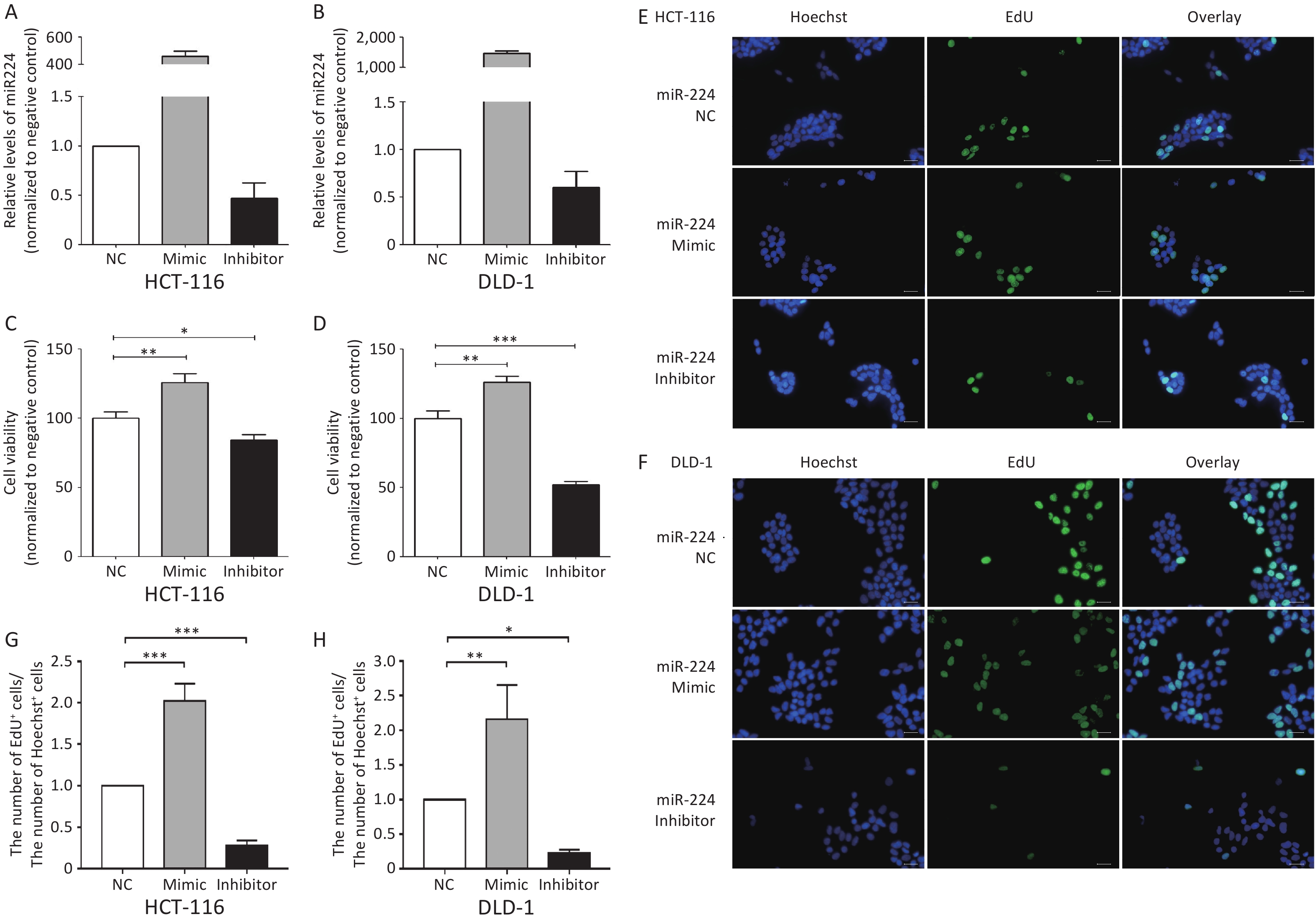
We first verified that miR-224-5p regulated CRC cell proliferation. The results showed that the proliferation of CRC cells was significantly increased after transfection with miR-224-5p mimic, while it was dramatically decreased after transfection with miR-224-5p inhibitor (Supplementary Figure S2, available in www.besjournal.com). We then further explored the possibility that this effect might be mediated by exosomes. Exosomes were extracted from the culture media of DLD1 and SW480 cells transiently transfected with miR-NC or miR-224-5p mimic. Subsequently, exosome feeding assays revealed that elevated exo-miR-224-5p had different regulatory effects on the proliferative ability of different CRC cells. Compared with their control groups, exo-miR-224-5p overexpression promoted cell viability of DLD1 (Figure 3A), while inhibiting cell viability of SW480 (Figure 3B). To further verify this differential regulation, we conducted an EdU incorporation assay and discovered the same regulatory trends (Figure 3C–F), which may be regulated by a certain underlying mechanism.
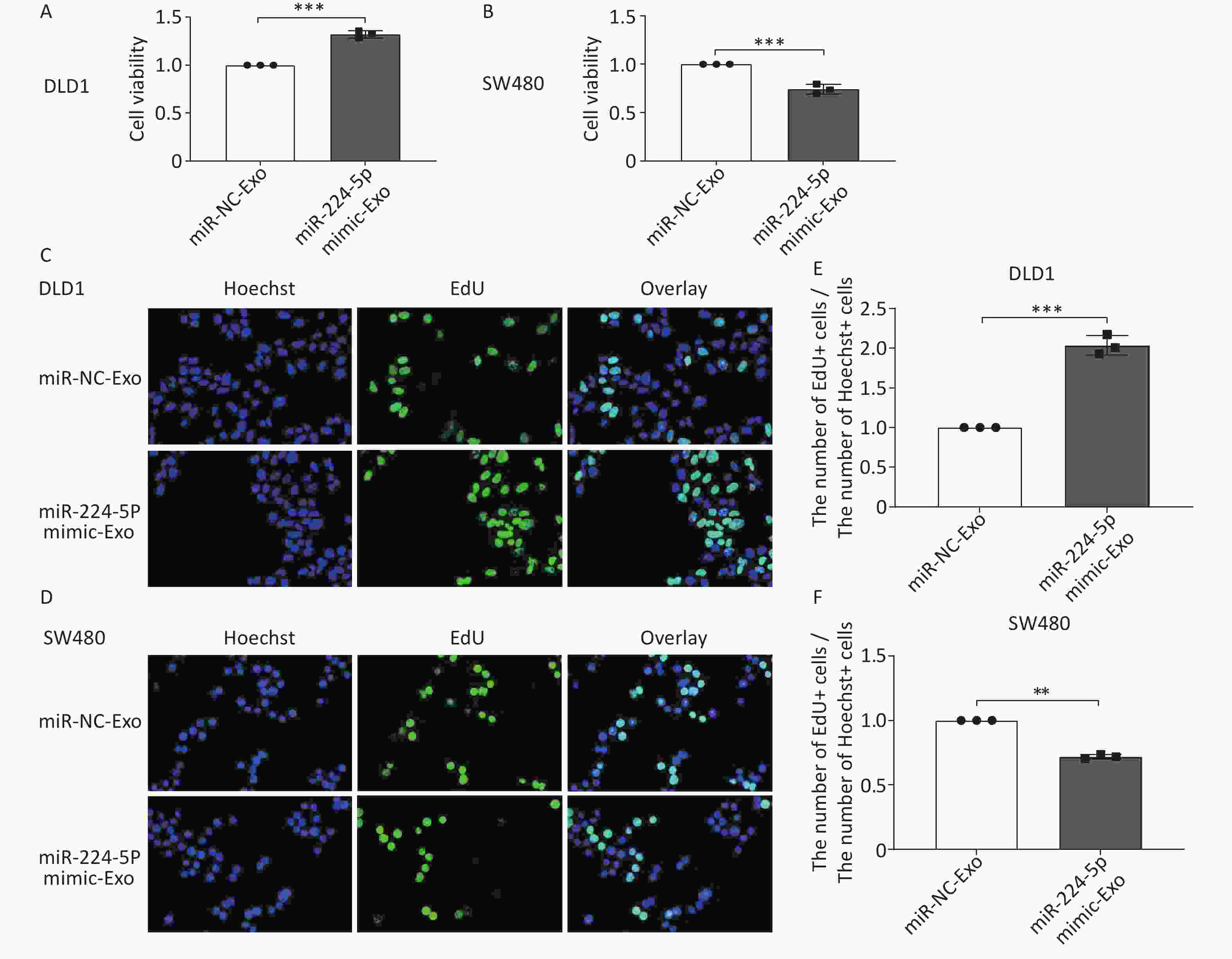
Figure 3. Exosomal miR-224-5p regulates CRC cell proliferation. (A, B) CCK8 assays were conducted to measure cell viability in DLD1 (A) and SW480 (B) cells after 72 h of exosome-feeding. (C, D) Representative images of Hoechest+ cells, EdU+ cells, and overlay in DLD1 (C) and SW480 (D) cells after 72 h of exosome-feeding (20×). (E, F) The ratio of EdU+ cell numbers to Hoechst+ cell numbers in DLD1 (E) and SW480 (F) cells after 72 h of exosome-feeding. Data are mean ± SD (n = 3). **P < 0.01, ***P < 0.001.
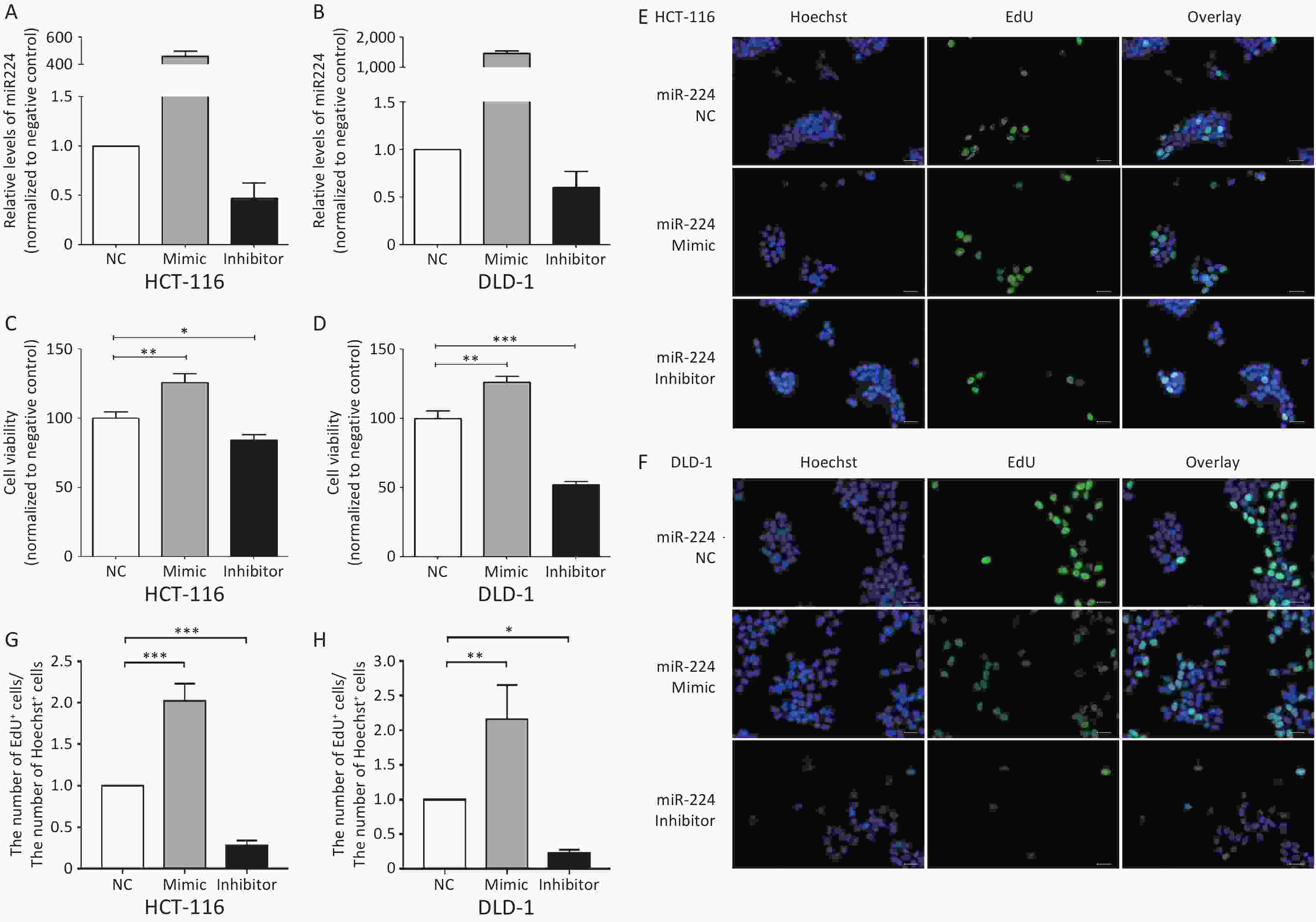
Figure S2. miR-224-5p promotes CRC cell proliferation. A and B Expression of miR-224-5p in HCT116 (A) and DLD1 (B) cells 48h after transfection with miR-224-5p mimic/inhibitor and miR-NC by qRT-PCR. Data are shown as mean ± SD (n = 3). C and D CCK-8 assays were conducted to detect cell viability 48 h after transfection with miR-224-5p mimic/inhibitor and miR-NC in HCT116 (C) and DLD1 (D) cells. Data are shown as mean ± SD (n = 3). *P < 0.05, **P < 0.01 & ***P < 0.001 vs. NC (Student’s t-test). E and F Representative images of Hoechest+ cells, EdU+ cells, and overlay 48 h after transfection with miR-224-5p mimic/inhibitor and miR-NC in HCT116 (E) and DLD1 (F) cells, using 40× objective lens. G and H The ratio of the number of EdU+ cells to the number of Hoechst+ cells 48 h after transfection with miR-224-5p mimic/inhibitor and miR-NC in HCT116 (G) and DLD1 (H) cells. Data are shown as mean ± SD (n = 3). *P < 0.05, **P < 0.01 & ***P < 0.001 vs. NC (Student’s t-test).
-
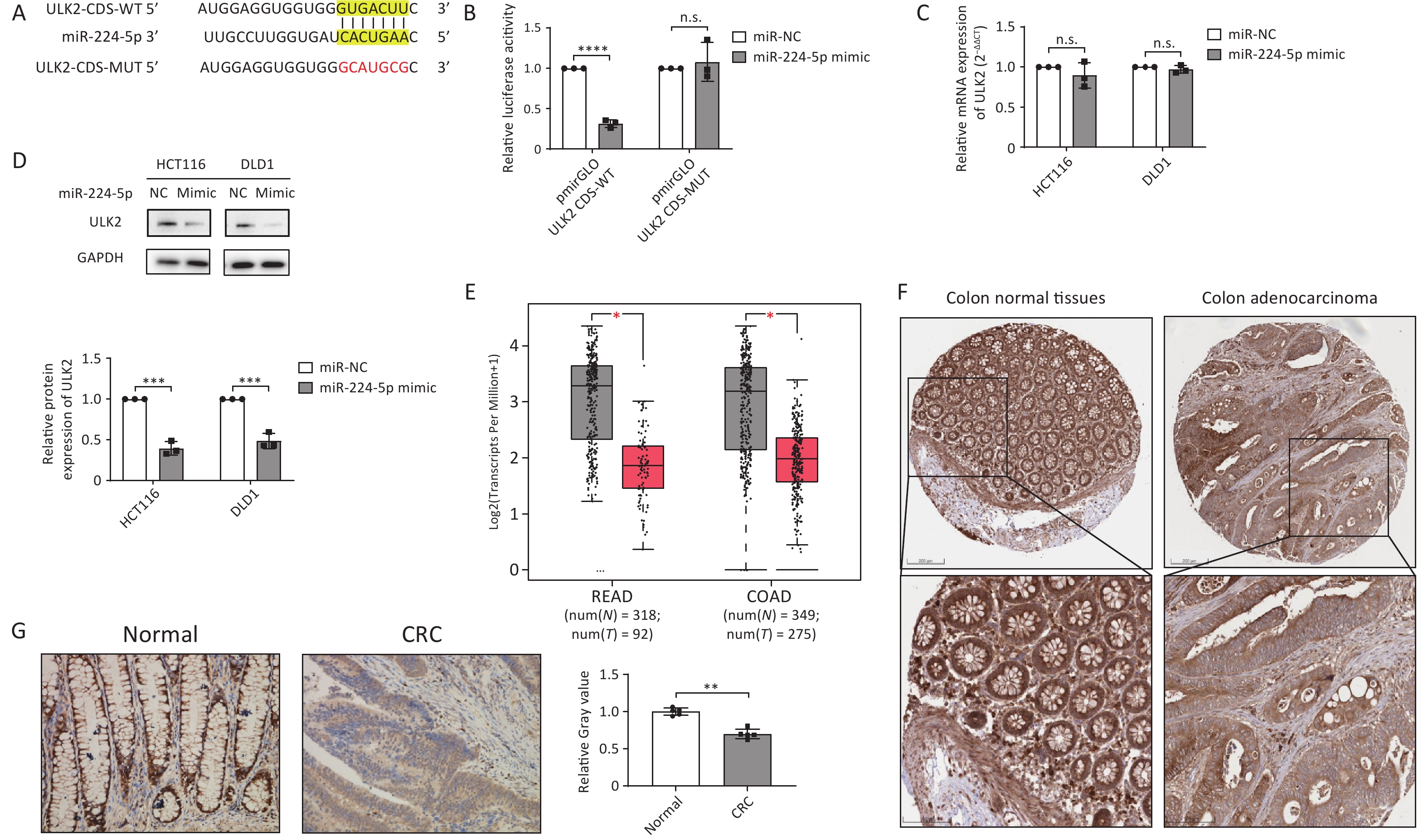
To investigate the molecular mechanism by which miR-224-5p contributes to CRC cell proliferation, we predicted ULK2 as a possible target for miR-224-5p with a putative binding site at 3’UTR by miRecords database. However, we found that the predicted binding site was actually located in the CDS of ULK2 mRNA through NCBI sequence alignment. Figure 4A and Supplementary Table S3 displayed the miR-224-5p binding site with ULK2 CDS. Then, this prediction was verified using the dual-luciferase reporter gene test. The results suggested that the relative luciferase activity in the cells co-transfected with miR-224-5p mimic and pmirGLO-ULK2-CDS-WT was significantly declined in comparison to the control group, indicating specific inhibition of the ULK2 CDS binding site by miR-224-5p mimic (Figure 4B). However, this decline could be eliminated when the ULK2 CDS was mutated (Figure 4B). Additionally, our data revealed that the ULK2 mRNA level in miR-224-5p-overexpressed CRC cells did not differ statistically from those in the control group (Figure 4C), whereas the ULK2 protein level was strikingly declined in these cells (Figure 4D).
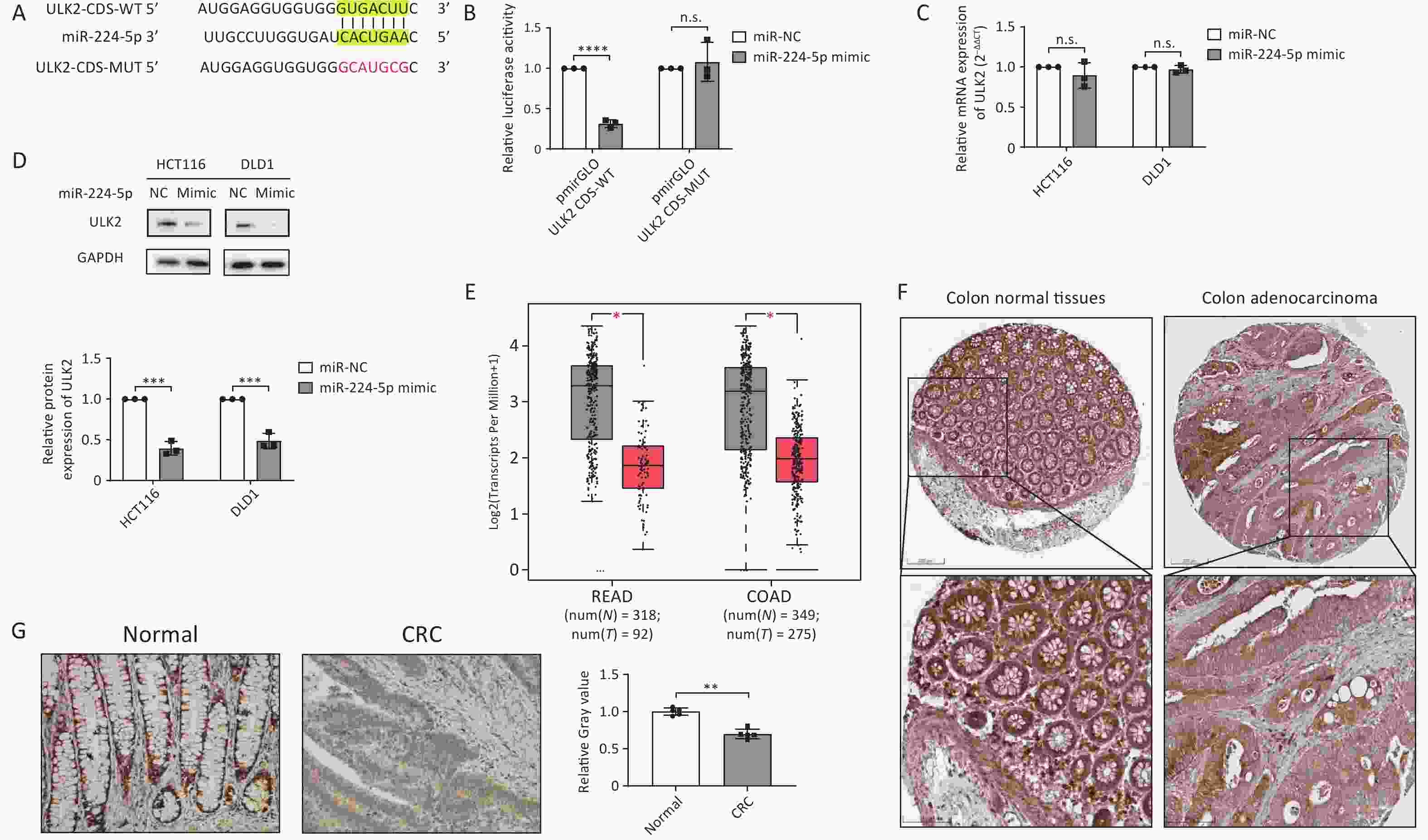
Figure 4. ULK2 is a direct target of miR-224-5p and is downregulated in CRC tissues. (A) The predicted MUT and corresponding WT binding site of ULK2 CDS with miR-224-5p. Red sequences represent mutant nucleotides while vertical line sequences show their binding site. (B) The dual-luciferase activity of HEK293T cells co-transfected of miR-NC or miR-224-5p mimic with pmirGLO-ULK2-CDS-MUT/WT. (C) qRT-PCR analysis of ULK2 mRNA expression. (D) Western blot analysis of ULK2 protein expression. (E) ULK2 transcript expression in READ and COAD tumor tissues (T) and normal tissues (N). (F) Immunohistochemistry pictures of ULK2 from HPA database. Scale bar, 200 nm (upper); Scale bar, 100 nm (lower). (G) Immunohistochemistry analysis of ULK2. Scale bar, 100 μm. Data are mean ± SD (n ≥ 3). n.s. means no significance, **P < 0.01, ***P < 0.001, ****P < 0.0001.
We further investigated the ULK2 expression in TCGA database, and the data showed transcripts of ULK2 in COAD and READ tumor tissues were both lower than in normal tissues (Figure 4E). Subsequently, we preliminarily observed that the ULK2 protein expression in CRC tissues was slightly lower than it was in normal colorectal epithelial tissues by referring to the IHC data from the Human Protein Altas (HPA) database (Figure 4F). Furthermore, paraffin sections of tumor tissues from 5 early CRC patients were selected for IHC staining. The results demonstrated that the positive rate of ULK2 in total cancer cells was somewhat lower than that in normal intestinal gland cells (Figure 4G), which was in accordance with the expression of ULK2 at the mRNA level.
Given the above evidence, miR-224-5p specifically binds to the CDS of ULK2 mRNA to inhibit ULK2 protein expression. ULK2 is downregulated in CRC tissues, which is negatively correlated with the expression of miR-224-5p in CRC.
-
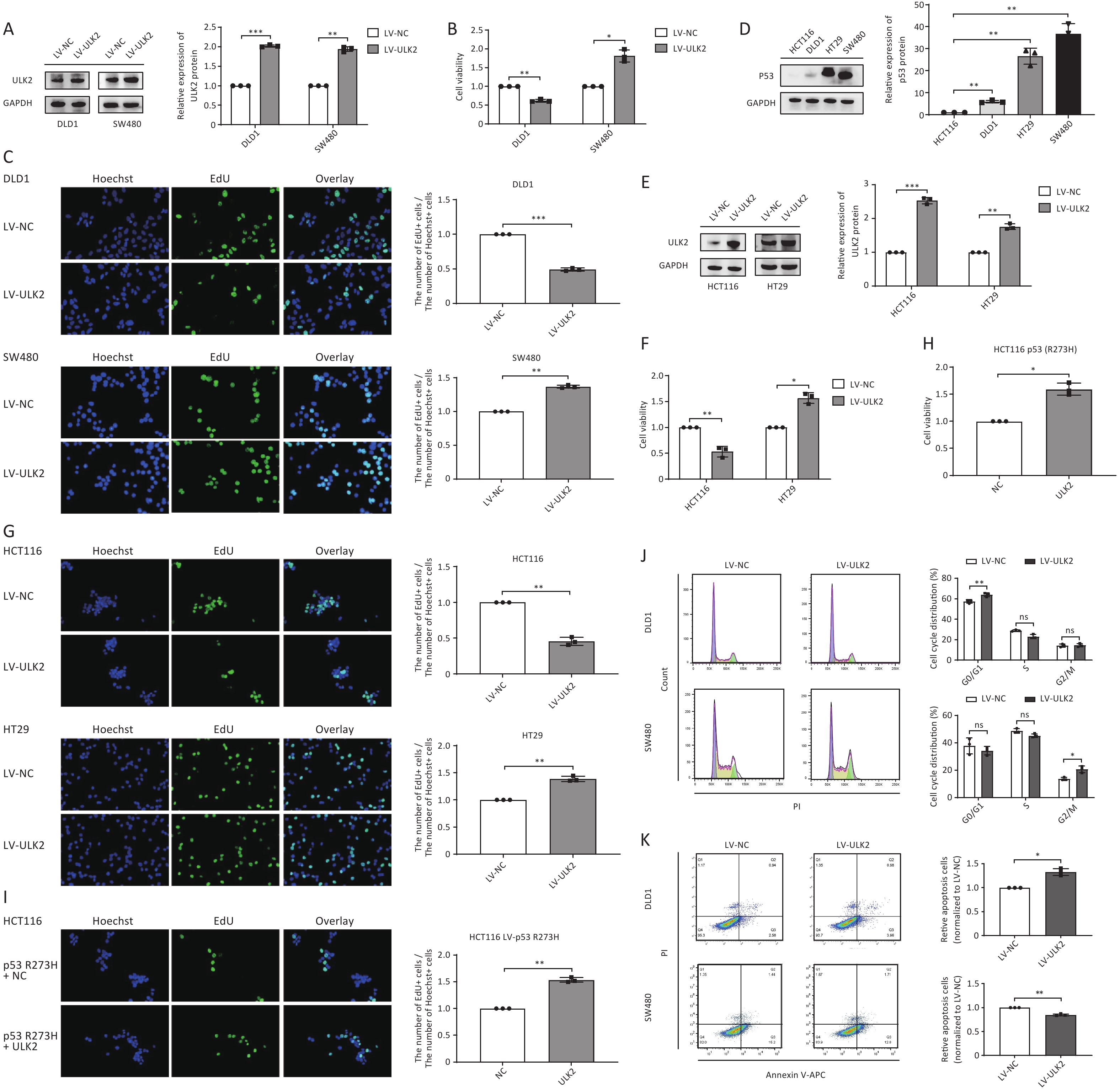
To investigate the biological role of ULK2 in CRC cell growth, we first constructed stably ULK2-overexpressed DLD1 and SW480 cells by lentivirus (LV) infection (Figure 5A). Compared with their respective control groups, we found that overexpressed ULK2 significantly inhibited cell viability and proliferative ability of DLD1 cells but promoted those of SW480 cells (Figure 5B and 5C). This differential regulation echoes the regulation of exosomal miR-224-5p on CRC cell proliferation, as we mentioned earlier.
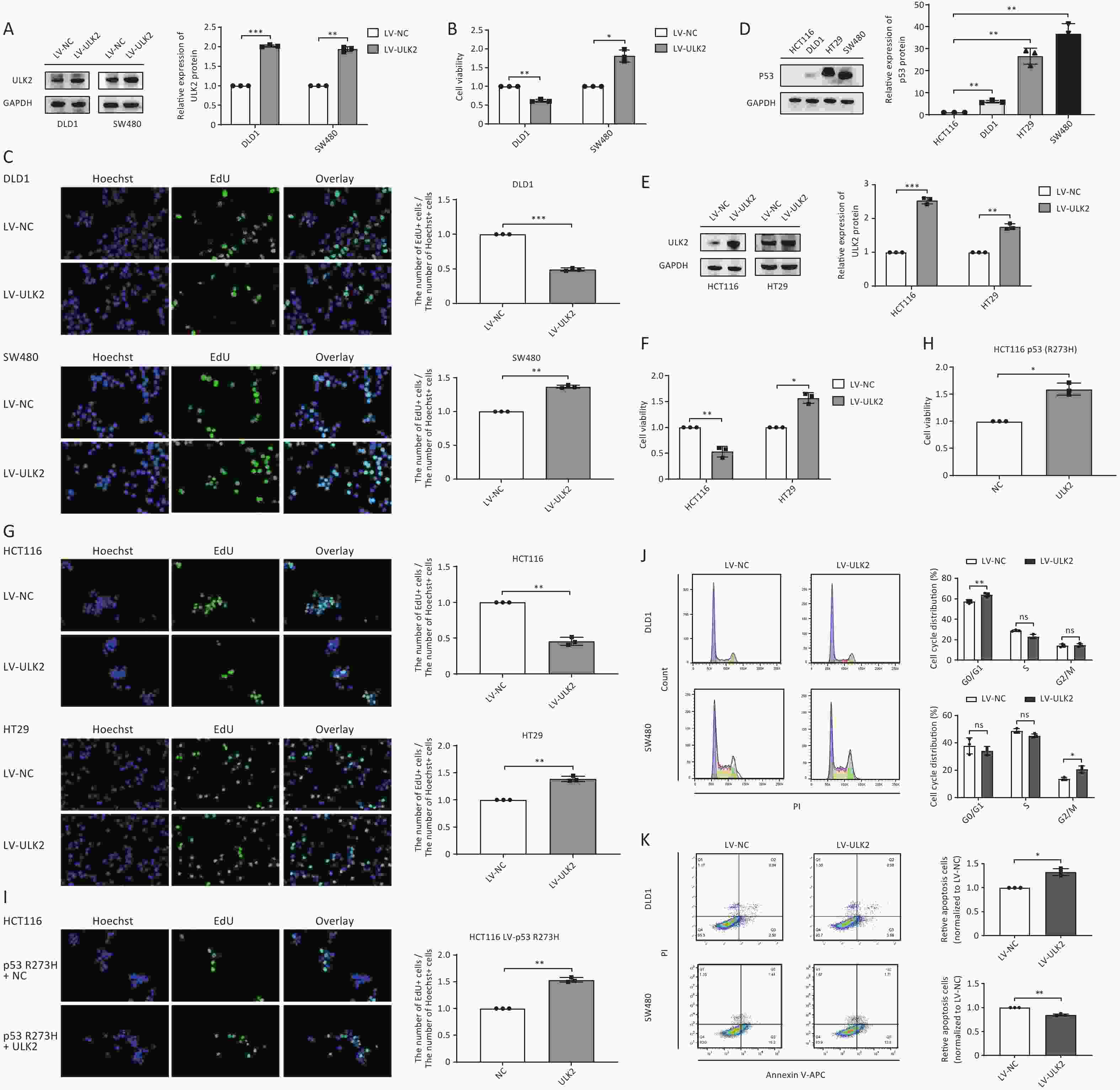
Figure 5. ULK2 regulates CRC cell proliferation in a p53-dependent manner. (A–C) Western blot analysis (A), CCK8 analysis (B), and EdU assay (C) (20×) in ULK2-overexpressed DLD1 and SW480 cells. (D) Western blot analysis of p53 protein expression in CRC cells. (E–G) Western blot analysis (E), CCK8 analysis (F), and EdU assay (G) (20×) in ULK2-overexpressed HCT116 and HT29 cells. (H, I) CCK8 analysis and EdU assay (20×) in p53 R273H-overexpressed HCT116 after transfection with NC or ULK2. (J, K) Flow cytometry analysis of ULK2 on the cell cycle (J) and cell apoptosis (K) in DLD1 and SW480 cells. Data are mean ± SD (n = 3). n.s. means no significance, *P < 0.05, **P < 0.01.
Given the findings above, we hypothesized that the differential regulation of exosomal miR-224-5p and ULK2 on CRC cell proliferation might be caused by the different molecular genetic backgrounds of the CRC cell line itself. We first searched the American Type Culture Collection (ATCC) database and found CRC cells had different p53 mutation statuses. Then, we discovered that p53 protein expression abundance in SW480 and HT29 cells with p53 R273H mutation was remarkedly higher than that in HCT116 and DLD1 cells (Figure 5D). Then, DNA sequencing results verified that SW480 and HT29 cells did exist p53 R273H mutation, while HCT116 and DLD1 cells did not (
Supplementary Figure S3 , available in www.besjournal.com).To clarify whether the regulation of ULK2 on CRC cell proliferation is related to P53 mutation sites and status, we also constructed stably ULK2-overexpressed HCT116 and HT29 cells (Figure 5E). Subsequently, CCK8 and EdU assays confirmed our suspicion that overexpressed ULK2 resulted in a suppression on HCT116 cell proliferation consistently with DLD1, whereas resulted in a facilitation on HT29 cell proliferation consistently with SW480 (Figure 5F and 5G). Then, we found that the proliferation ability of p53 R273H-overexpressed HCT116 cells after transfection with ULK2 was dramatically enhanced in contrast to the control group (Figure 5H and 5I), reflecting the cancer-promoting effect of p53 with R273H mutation.
Additionally, we further explored how ULK2 affects CRC cell cycle progression and apoptosis. The results showed that overexpressed ULK2 facilitated the G0/G1 phase arrest and reduced the apoptosis rate in DLD1 cells, but accelerated the cell cycle progression and elevated the proportion of cells in the G2/M phase (Figure 5J and 5K). According to the above results, this differential regulation may also be regulated by p53 to some extent.
-
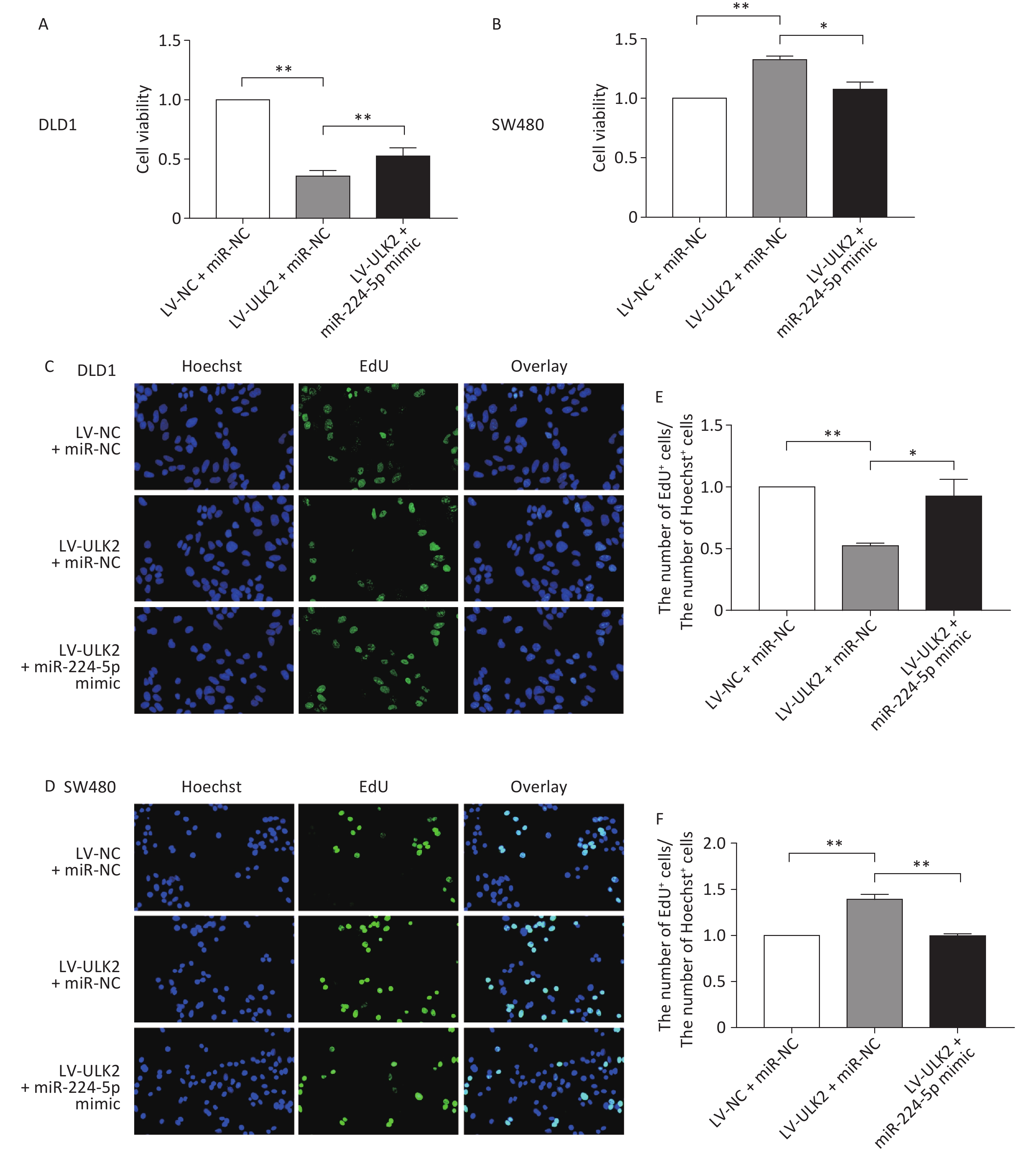
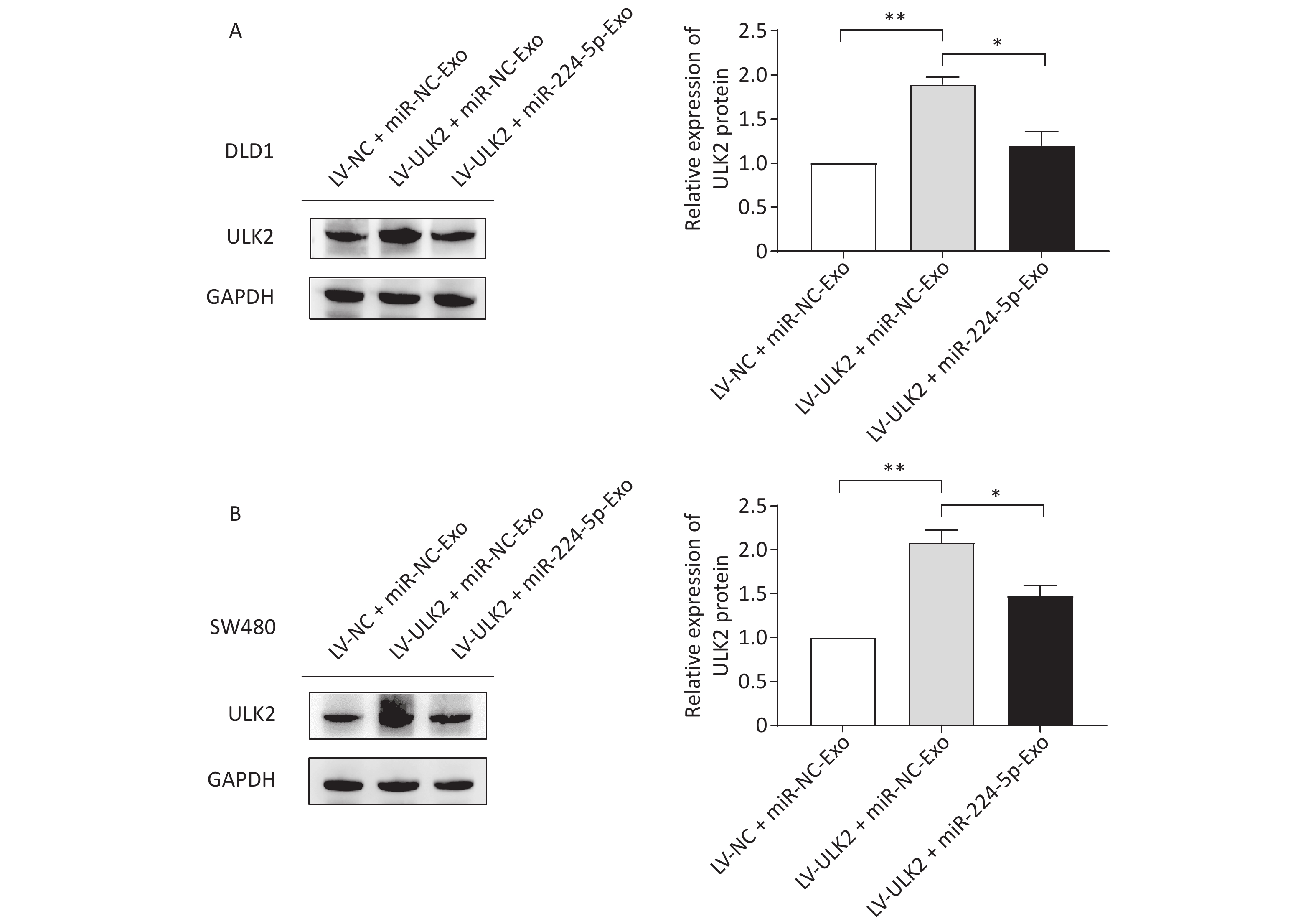
To further confirm the impact of the miR-224-5p-ULK2 axis on CRC cell proliferation, we first proved that exogenous miR-224-5p can partially reverse the proliferation regulation of ULK2 on CRC cells (Supplementary Figure S4, www.besjournal.com). In addition, miR-224-5p mimic-exo feeding significantly neutralized the promotion effect of LV-ULK2 on ULK2 expression in DLD1 and SW480 cells (Supplementary Figure S5, www.besjournal.com).
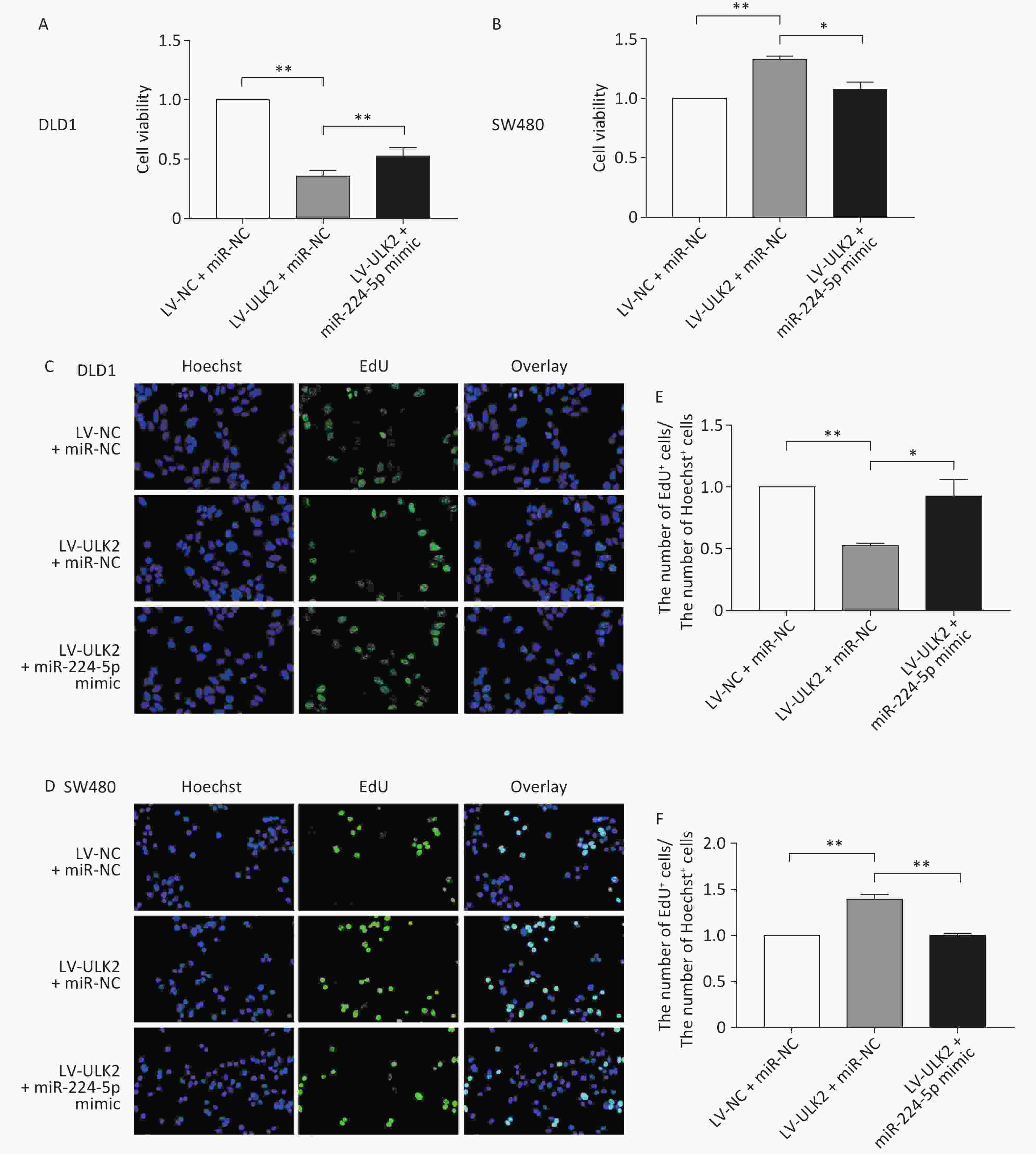
Figure S4. Exogenous miR-224-5p partially reverses the regulation effect of ULK2 on CRC cell proliferation. A and B CCK-8 assays were conducted to detect cell viability after transfection with miR-224-5p mimic/miR-NC in ULK2-overexpressed DLD1 (A) and SW480 (B) cells. Data are shown as mean ± SD (n = 3). **P < 0.01 vs. LV-NC+miR-NC; *P < 0.05 & **P < 0.01 vs. LV-ULK2 + miR-NC (Student’s t-test). C and D Representative images of Hoechest+ cells, EdU+ cells, and overlay after transfection with miR-224-5p mimic/miR-NC in ULK2-overexpressed DLD1 (C) and SW480 (D) cells, using 20×objective lens. E and F The ratio of the number of EdU+ cells to the number of Hoechst+ cells after transfection with miR-224-5p mimic/miR-NC in ULK2-overexpressed DLD1 (E) and SW480 (F) cells. Data are shown as mean ± SD (n = 3). **P < 0.01 vs. LV-NC+miR-NC; *P < 0.05 & **P < 0.01 vs. LV-ULK2+miR-NC (Student’s t-test).
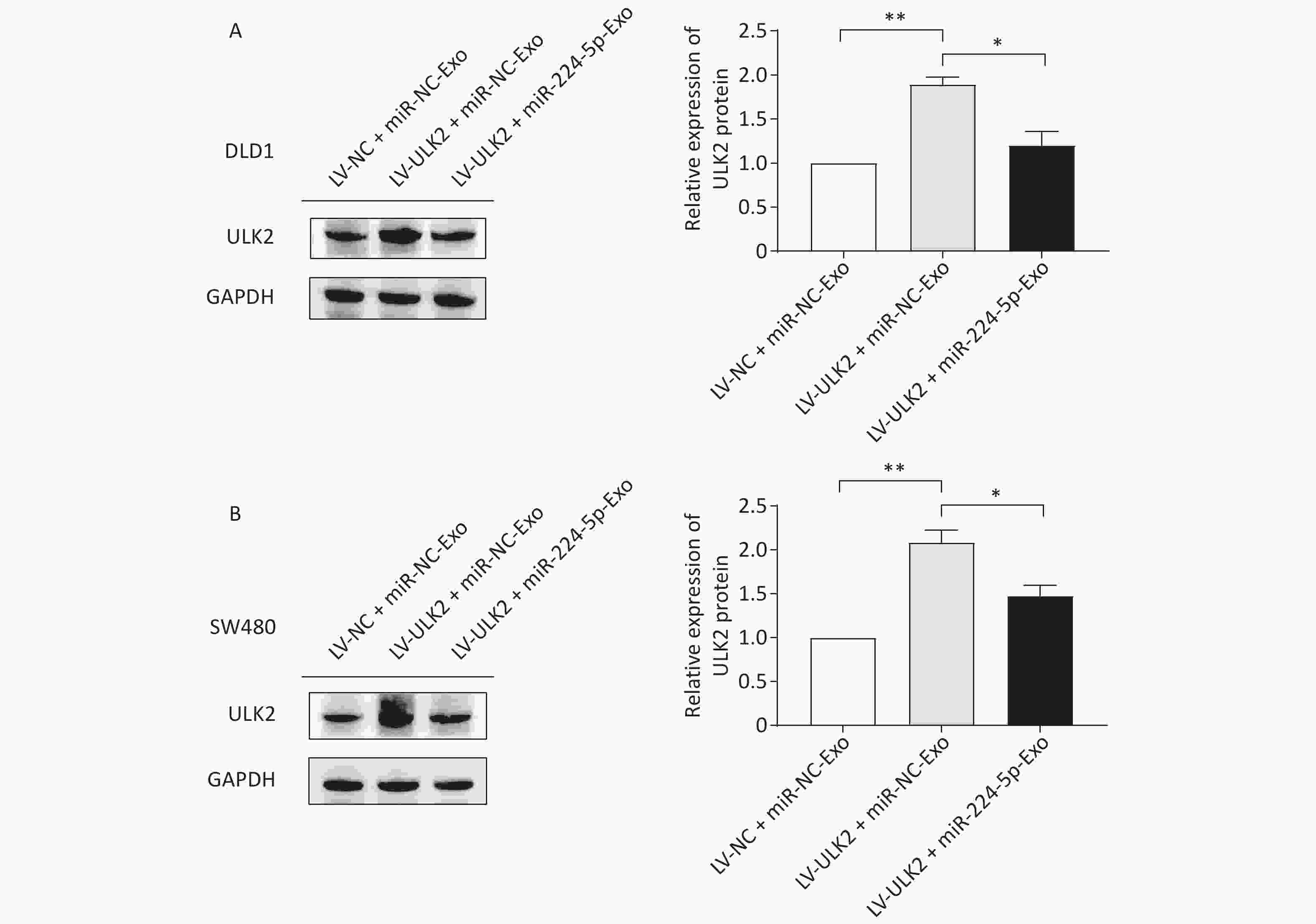
Figure S5. Effects of exosomal miR-224-5p on ULK2 expression in CRC cells. (A) and (B) Western blot assays of ULK2 expression in LV-NC/LV-ULK2-transfected DLD1 (A) and SW480 (B) cells after being treated with miR-NC-Exo/ miR-224-5p-Exo. Data are shown as mean ± SD (n = 3). **P < 0.01 vs. LV-NC + miR-NC-Exo; *P < 0.05 vs. LV-ULK2+miR-NC-Exo (Student’s t-test).
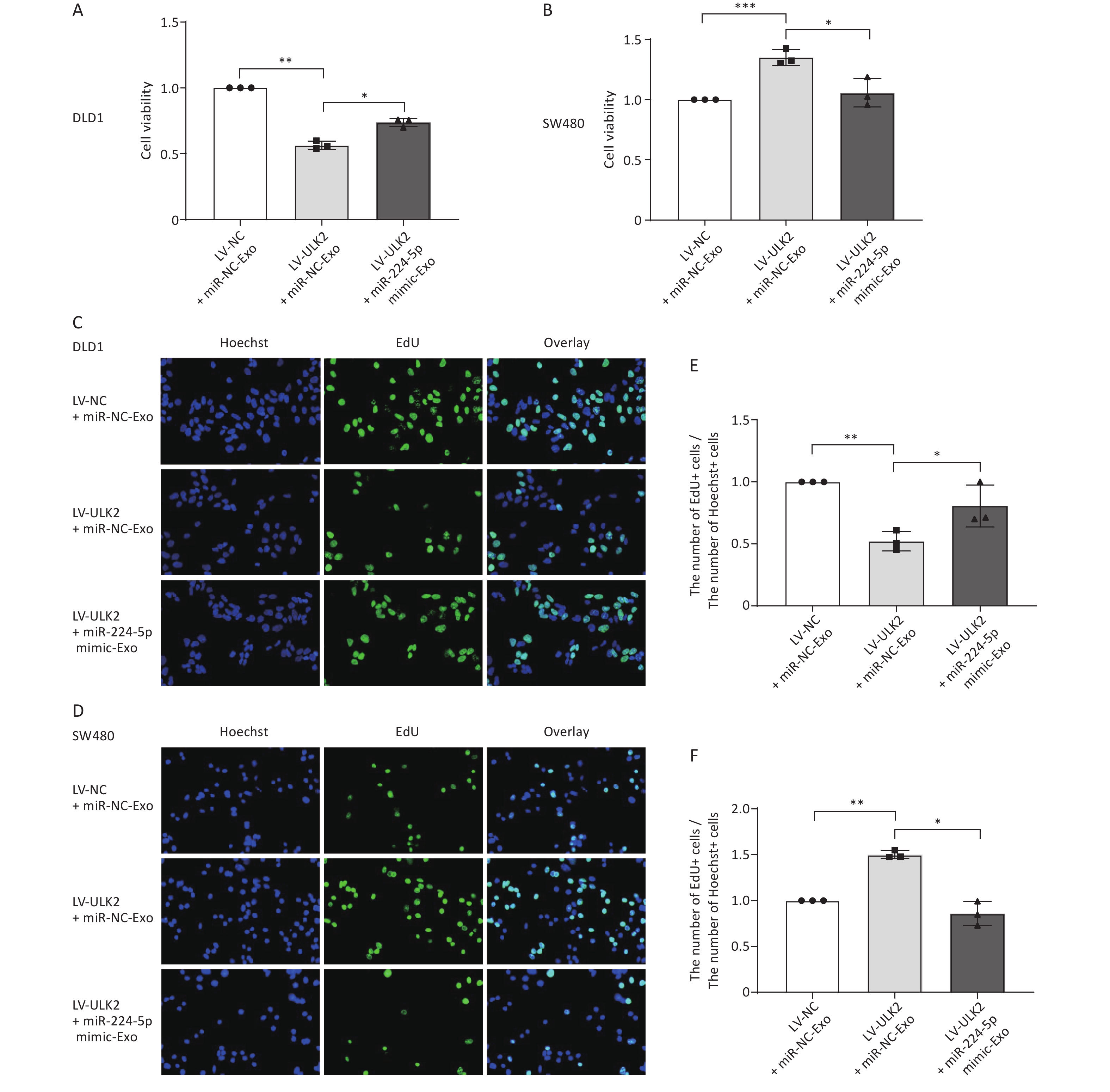
Next, we investigated whether exo-miR-224-5p is involved in this reversal regulation. After transiently transfected miR-NC or miR-224-5p mimic, exosomes were extracted from the cell culture medium and then added to ULK2-overexpressed DLD1 and SW480 cells. Notably, ULK2-regulated proliferation of DLD1 and SW480 cells was partially reversed with miR-224-5p mimic-exo feeding (Figure 6). Taken together, these findings indicate that exo-miR-224-5p might promote the proliferation of recipient CRC cells with p53 273 site wild-type but inhibit the proliferation of recipient CRC cells with p53 R273H mutation by targeting ULK2.
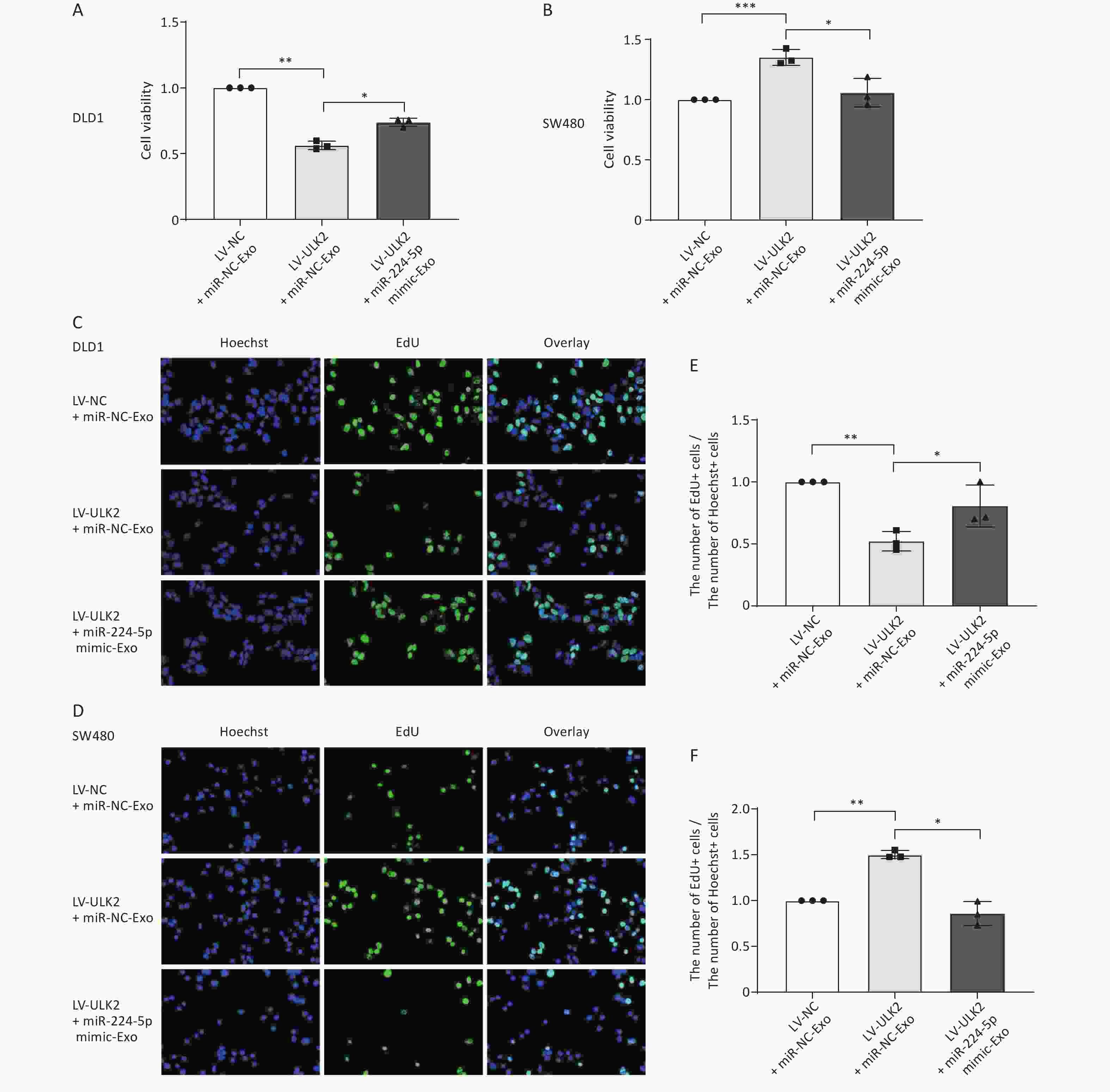
Figure 6. Exosome-transmitted miR-224-5p partially reverses the regulation effect of ULK2 on CRC cell proliferation. (A, B) CCK8 analysis of cell viability in ULK2-overexpressed DLD1 (A) and SW480 (B) cells after transfection with miR-NC-Exo or miR-224-5p mimic-Exo. (C–F) EdU assay in ULK2-overexpressed DLD1 (C, E) and SW480 (D, F) cells after transfection with miR-NC-Exo or miR-224-5p mimic-Exo (20×). Data are mean ± SD (n = 3). *P < 0.05, **P < 0.01, ***P < 0.001.
-
This study demonstrated that miR-224-5p was upregulated in CRC pure epithelium cells of resected tumor tissue. ULK2 was characterized as a direct target of miR-224-5p that selectively binds to the CDS of ULK2 mRNA to inhibit ULK2. Furthermore, miR-224-5p could be enriched in CRC-derived exosomes, secreted into the tumor microenvironment, and then transported to adjacent receptor cells efficiently in the form of exo-miR-224-5p. Moreover, we found that the proliferation effect of ULK2 on different CRC cell lines was different due to the mutation status of p53 R273H. Furthermore, our results also showed that exo-miR-224-5p could regulate CRC cell proliferation by directly inhibiting ULK2 in vitro (Figure 7).
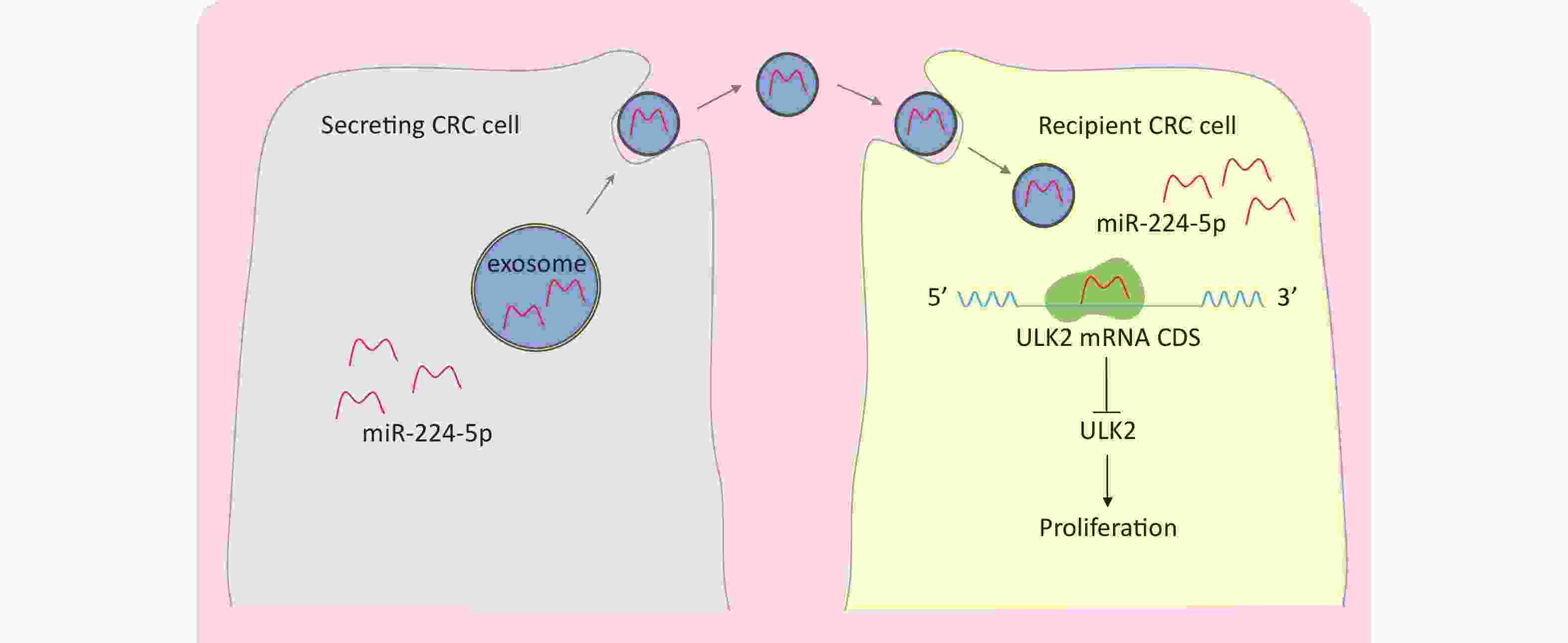
Figure 7. A schematic model of the role of exo-miR-224-5p in regulating CRC cell with no p53 R273H mutation.
The expression of miR-224-5p increased progressively with tumor stage in CRC tissues. It is suggested that miR-224-5p has great potential as an independent risk factor for CRC. It is the first time that we report the comprehensive expression of miR-224-5p in gut diseases by using pure tumor cells as test samples captured by LCM. At present, most of the experimental samples used in studies on miRNA expression are whole tumor tissues, from which obtained biological information is not only from tumor cells but also from stromal cells and many infiltrating lymphocytes. Such results are not quite reliable and can lead to many specific signals being obscured by other confounding signals. Only the miRNA expression profile detected from pure tumor cells can accurately reflect the real expression situation in vivo and may become a reliable tumor marker. Therefore, our data may be more subdivided and accurate than TCGA data.
In addition, our prior study revealed that miR-224-5p expression was not detected in peripheral blood among CRC patients or healthy individuals[24]. Also, our vitro experiments proved that overexpressed miR-224-5p could be secreted into the supernatant of CRC cells. These results are in line with our hypothesis that miR-224-5p derived from colorectal cancer cells is not released into the tumor microenvironment through endocrine but through the manner of autocrine or paracrine.
MiR-224-5p is secreted into the extracellular environment via exosomes and then taken up by adjacent receptor CRC cells. Although microRNA therapies have demonstrated considerable promise for secure and effective cancer treatment, the main obstacle to miRNA delivery is their instability in the microenvironment[25,26]. The appearance of exosomes brings hope to the application of this therapy. Due to many beneficial properties of exosomes, including circulatory stability, low toxicity, low immunogenicity, and biocompatibility, targeted delivery of functional miRNAs mediated by exosomes is a new strategy for precision cancer therapy[27]. Liang et al. successfully reversed 5-fluorouracil resistance in colon cancer by applying engineered exosomes to deliver chemotherapeutics and miR-21 inhibitor to drug-resistant HCT116 cells[28], suggesting that modifying or inhibiting exosomal miRNAs could be an effective therapeutic approach for cancer. Deeper exploration into the precise molecular mechanism of exosome-mediated tumorigenesis will raise the possibility that exosome-derived miR-224-5p can serve as a diagnostic and therapeutic target towards precision medicine.
Furthermore, exo-miR-224-5p regulates CRC cell proliferation by targeted downregulation of ULK2. This suggests that miR-224-5p may be an ideal candidate for targeted therapeutic intervention in CRC expressing low levels of ULK2. Despite the paucity of studies on the function and regulatory mechanism of ULK2 in disease pathogenesis, it has been demonstrated that changes in ULK2 expression have a significant impact on tumor occurrence and development[29-32], providing perspective into the implications for CRC therapies that target ULK2. Currently, chemotherapeutic drugs are used as an auxiliary means for CRC treatment, while autophagy inhibitors have been shown to improve the effectiveness of chemotherapy in cancer[33]. As a direct target of miR-224-5p, ULK2 is a crucial autophagy-associated gene that activates the autophagy pathway[34]. Hence, the application of miR-224-5p/ULK2 inhibitors may be a promising strategy for CRC adjuvant chemotherapy.
Last but not least, we found that overexpression of ULK2 suppressed cell proliferation in CRC cells with no p53 R273H mutation while promoting it in CRC cells with p53 R273H mutation. This indicated that the p53 mutation status of cells would significantly affect the regulation of gene expression. Similar findings were reported by Scarola et al.[35], who found that overexpressed miR-335 reduced Rb1 protein and activated the p53 tumor inhibitor pathway to prevent cell proliferation in tumor cells expressing the p53 wild-type allele, but boosted overproliferation when the p53 tumor inhibitor pathway was absent. In our study, exo-miR-224-5p suppressed the proliferation of CRC cells with p53 R273H mutation. Therefore, targeted delivery of miR-224-5p mimic using exosomes as drug carriers for CRC patients with p53 R273H mutation may be a valuable therapeutic strategy. Likewise, targeted delivery of miR-224-5p inhibitor mediated by exosomes for CRC patients with p53 wild-type is also a novel idea for anticancer therapy. To realize that, having a good command of the relationship and potential mechanism between the miR-224-5p-ULK2 axis and p53 is necessary.
The present study had some limitations. First, although we found an interaction between exosomes and receptor CRC cells, a question that remains to be answered is whether their interaction is achieved through adhesion or internalization. Second, the role of miR-224-5p-deficient exosomes on receptor CRC cells needs to be further explored. Finally, there are some challenges in excluding the effects of microvesicles despite our evidence revealing the role and mechanism of exo-miR-224-5p on CRC cell proliferation.
In summary, we found a new way that miR-224-5p regulates the proliferation of colorectal cancer cells through the paracrine or autocrine pathway, which is realized by tumor-secreting exosomes. Furthermore, miR-224-5p-enriched exosomes enhance cell proliferation of CRC with no p53 R273H mutation via directly targeting ULK2, perhaps providing prospective targets for CRC prevention and therapy.
HTML
Clinical Tissue Samples
Cell Culture
RNA Extraction and qRT-PCR Analysis
Reconstruction of the CDS of ULK2
miRNAs Transfection and Reconstructed pmirGLO Co-transfection
Dual-luciferase Reporter Gene Assay
Cell Cycle Assay
Cell Apoptosis Analysis
Western Blot Analysis
Co-culture Assay
Isolation of Exosomes
Internalization of Exosomes
Cell Viability Analysis
Lentiviral Vector Construction and Transduction
5-Ethynyl-2′-deoxyuridine. (EdU) Incorporation Assay
Bioinformatic Analysis
Immunohistochemistry (IHC) Analysis
Laser Capture Microdissection (LCM) and miRNA Microarray
Genomic DNA Extraction, PCR Amplification, and Sequencing Analysis
Statistical Analysis
miR-224-5p is Upregulated in CRC Tissues and CRC Cell-derived Exosomes
CRC Cells Secrete Exosomal miR-224-5p and Target Adjacent Tumor Cells
miR-224-5p-enriched Exosomes Regulate CRC Cell Proliferation in vitro
ULK2 is a Direct Target of miR-224-5p
ULK2 Inhibits CRC Cell Proliferation in a p53-dependent Manner
Exosome-transmitted miR-224-5p Partially Reverses the Regulation Effect of ULK2 on CRC Cell Proliferation
&These authors contributed equally to this work.
 23053+Supplementary Materials.pdf
23053+Supplementary Materials.pdf
|

|

 Quick Links
Quick Links
 DownLoad:
DownLoad: