Objective To in vestigate potential gender differences in the odds of overweight/obese, weight-related perceptions, and behaviors among Chinese school children.Methods Height, weight, and a survey of weight-related perceptions and behaviors were measured in a nationally representative survey of 12, 811 children in primary schools in China. Logistic regression analyses were used to assess gender differences, adjusting for confounders.Results Boys had higher odds of being overweight/obese compared to girls within both urban[adjusted odds ratio (OR) 2.30, 95% CI 2.00 to 2.65] and rural areas (OR=1.85, 95% CI 1.55 to 2.20). Girls reported healthier diets (e.g., daily vegetables OR=0.79, 95% CI 0.73 to 0.85) whereas boys consumed fried food (OR=1.21, 95% CI 1.06 to 1.38) and sugar-sweetened drinks more often (OR=1.49, 95% CI 1.34 to 1.65). Gender differences included higher odds of boys perceiving themselves as overweight if they had more highly educated mothers (OR=1.35, 95% CI 1.09 to 1.68), less educated fathers (OR=0.79, 95% CI 0.63 to 0.99), and if they frequently consumed carbonated drinks (OR=1.48, 95% CI 1.07 to 2.05).Conclusion Childhood obesity prevention in China should be gender-focused, particularly for boys who reported an unhealthier diet but were less likely to see they were fat, even though more boys were overweight or obese than girls.
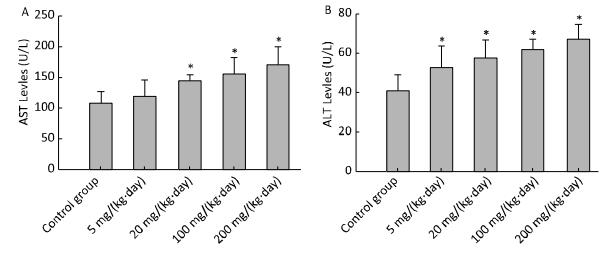
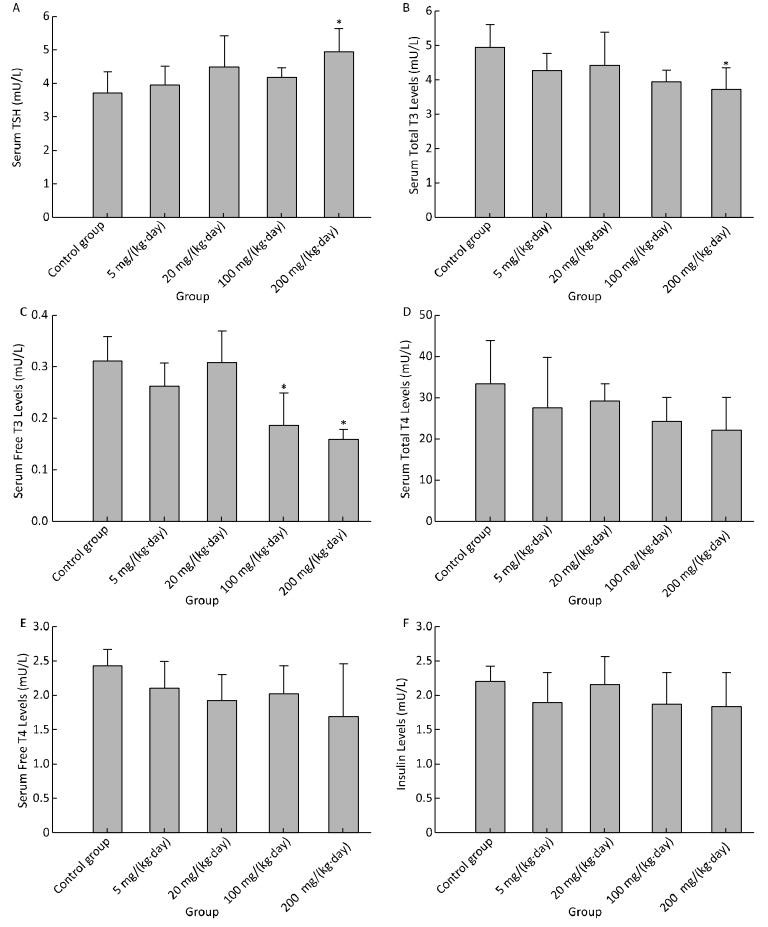
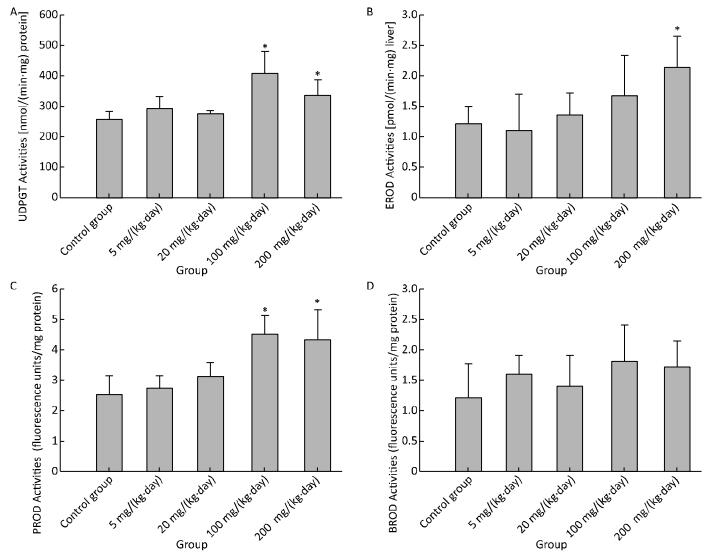
Objective This study aimed to evaluate the hepatotoxicity, metabolic disturbance activity and endocrine disrupting activity of mice treated by Decabromodiphenyl ethane (DBDPE).Methods In this study, Balb/C mice were treated orally by gavage with various doses of DBDPE. After 30 days of treatment, mice were sacrificed; blood, livers and thyroid glands were obtained, and hepatic microsomes were isolated. Biochemical parameters including 8 clinical chemistry parameters, blood glucose and hormone levels including insulin and thyroid hormone were assayed. The effects of DBDPE on hepatic cytochrome P450 (CYP) levels and activities and uridinediphosphate-glucuronosyltransferase (UDPGT) activities were investigated. Liver and thyroid glands were observed.Results There were no obvious signs of toxicity and no significant treatment effect on body weight, or liver-to-body weight ratios between treatment groups. The levels of ALT and AST of higher dose treatment groups were markedly increased. Blood glucose levels of treatment groups were higher than those of control group. There was also an induction in TSH, T3, and fT3. UDPGT, PROD, and EROD activities were found to have been increased significantly in the high dose group. Histopathologic liver changes were characterized by hepatocyte hypertrophy and cytoplasmic vacuolization. Our findings suggest that DBDPE can cause a certain degree of mouse liver damage and insufficiency.Conclusion DBDPE has the activity of endocrine disruptors in Bal/C mice, which may induce drug-metabolizing enzymes including CYPs and UDPGT, and interfere with thyroid hormone levels mediated by AhR and CAR signaling pathways. Endocrine disrupting activity of DBDPE could also affect the glucose metabolism homeostasis.
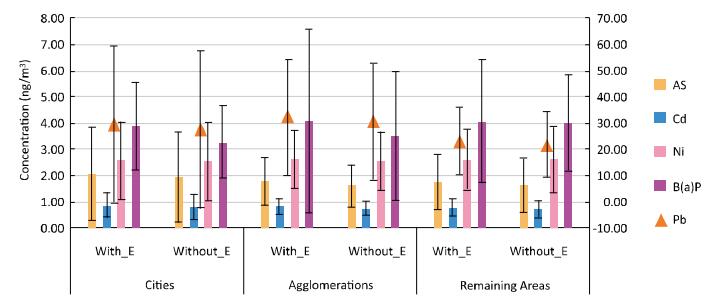
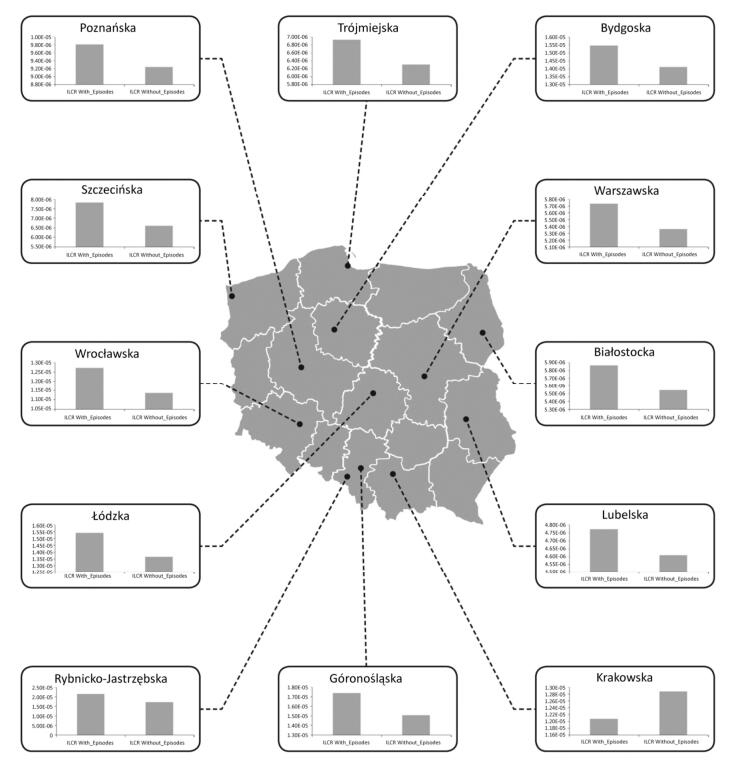
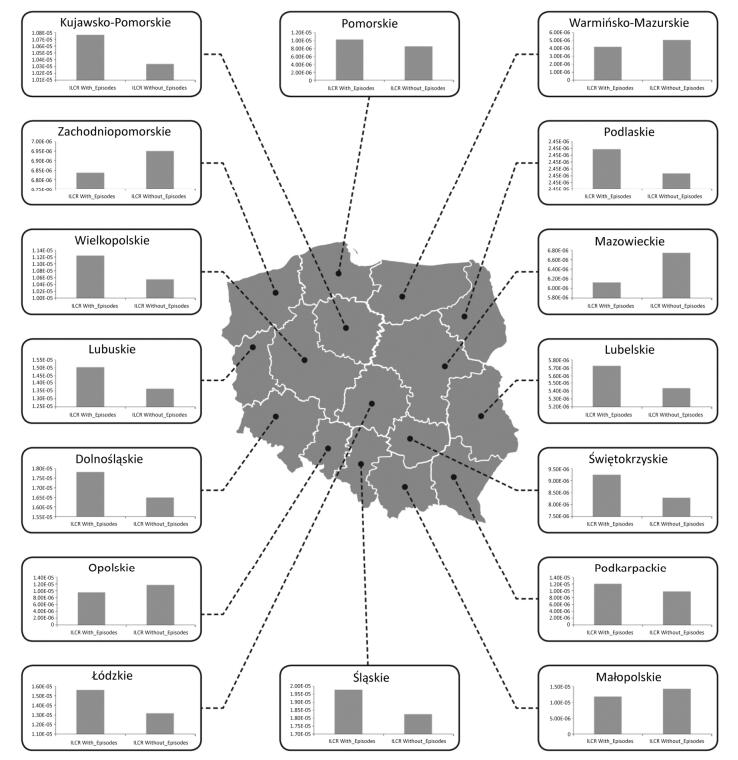
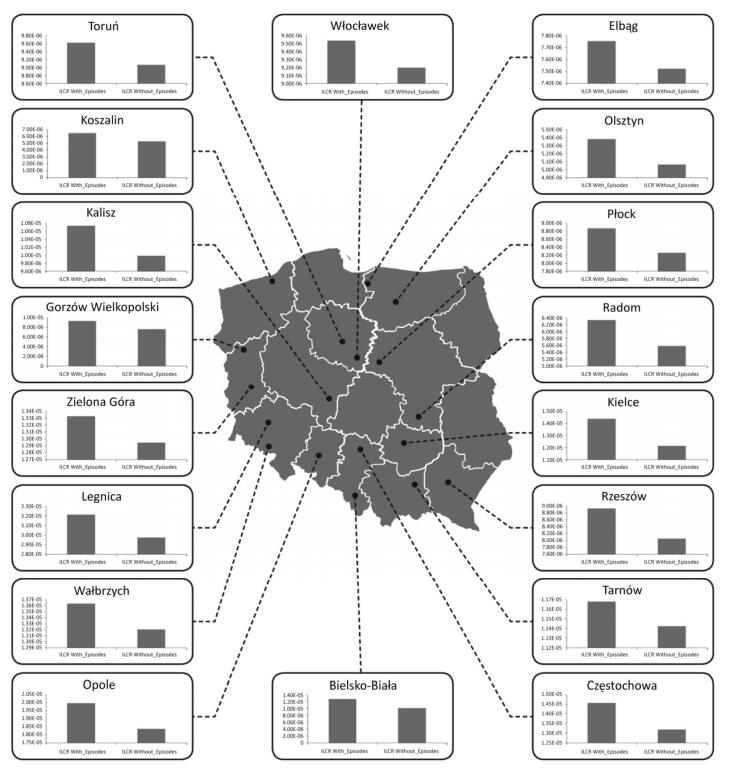
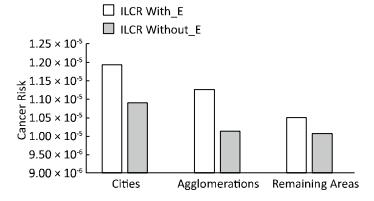
Objective To check whether health risk impacts of exposure to airborne metals and Benzo(a) Pyrene during episodes of high PM10 concentrations lead to an increased number of lung cancer cases in Poland.Methods In this work, we gathered data from 2002 to 2014 concerning the ambient concentrations of PM10 and PM10-bound carcinogenic Benzo(a)pyrene[B(a)P] and As, Cd, Pb, and Ni. With the use of the criterion of the exceedance in the daily PM10 mass concentration on at least 50% of all the analyzed stations, the PM10 maxima's were selected. Lung cancer occurrences in periods with and without the episodes were further compared.Results During a 12-year period, 348 large-scale smog episodes occurred in Poland. A total of 307 of these episodes occurred in the winter season, which is characterized by increased emissions from residential heating. The occurrence of episodes significantly (P < 0.05) increased the concentrations of PM10-bound carcinogenic As, Cd, Pb, Ni, and B(a)P. During these events, a significant increase in the overall health risk from those PM10-related compounds was also observed. The highest probability of lung cancer occurrences was found in cities, and the smallest probability was found in the remaining areas outside the cities and agglomerations.Conclusion The link between PM pollution and cancer risk in Poland is a serious public health threat that needs further investigation.
Objective We aimed to elucidate the rates of repeat HIV testing and incident HIV diagnosis, and baseline CD4+ T cell count among individuals attending HIV voluntary counseling and testing (VCT) clinics in Wuxi, China.Methods A repeat HIV testing within 12 months was recorded if individuals had their first test with negative results, during 2013-2014 and retested within 12 months. An incident HIV diagnosis was recorded if individuals had their first test with negative results, during 2013-2015 and had a subsequent positive result at any point by the end of 2015. Data on HIV testing and diagnosis among individuals attending 32 VCT clinics from 2013 to 2015 and HIV diagnosis from other clinical services in Wuxi, China, were retrieved. A multivariate logistic regression model was used to analyze factors associated with repeat HIV testing. Cox regression was used to evaluate factors associated with incident HIV diagnosis.Results From 2013 to 2014, 11, 504 individuals tested HIV negative at their first recorded test, with 655 (5.7%) retesting within 12 months. Higher repeat HIV testing within 12 months was associated with male gender[adjusted odds ratio (aOR)=1.7, 95% confidence interval (CI):1.4-2.2], risk behaviors[commercial heterosexual behaviors (aOR=1.4, CI:1.1-1.6), male-male sexual behaviors (aOR=3.7, CI:2.7-4.9)], injection drug use (aOR=9.9, CI:6.5-15.1), and having taken HIV tests previously (aOR=2.0, CI:1.6-2.4). From 2013 to 2015, 1, 088 individuals tested negative on HIV test at their visit and at ≥ 2 subsequent tests; of them 30 had incident HIV diagnosis. The overall rate of incident HIV diagnosis among all VCT individuals was 1.6 (95% CI:1.1-2.1) per 100 person-years. Incident HIV diagnosis was associated with male gender[adjusted hazard ratio (aHR)=8.5, 95% CI:1.9-38.1], attending hospital-based VCT clinics (aHR=7.8, 95% CI:1.1-58.3), and male-male sexual behavior (aHR=8.4, 95% CI:1.5-46.7). Individuals diagnosed at VCT clinics had higher CD4+ T cell count compared with those diagnosed at other clinical services (median 407 vs. 326 copies/mm3, P=0.003).Conclusion VCT individuals in Wuxi, China, had a low repeat HIV testing rate and high HIV incidence. VCT-clinic-based interventions aimed at increasing repeat HIV testing are needed to detect more cases at an
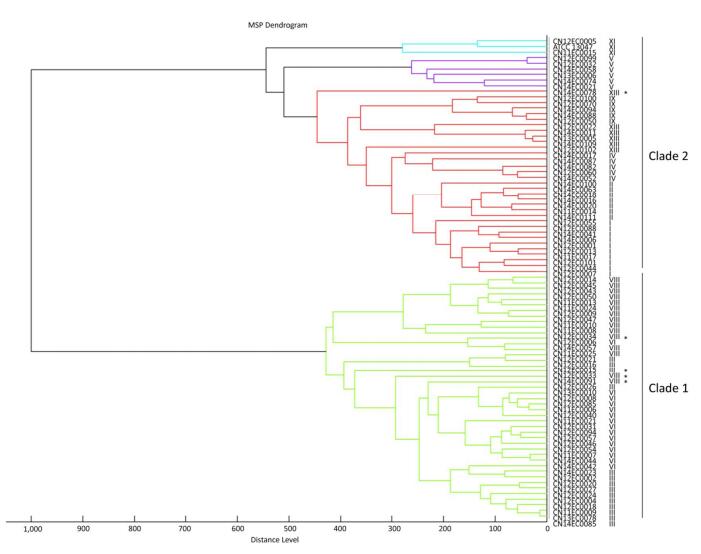
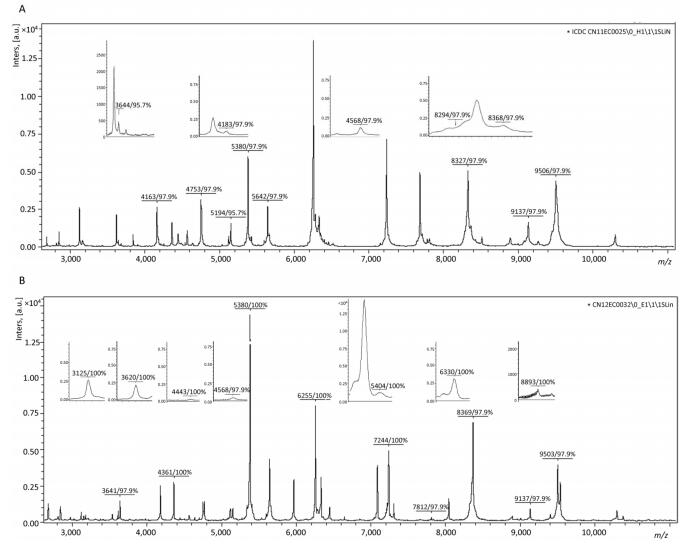
Objective To establish a domestic database of Enterobacteria cloacae (E. cloacae), and improve the identification efficiency using peptide mass fingerprinting.Methods Peptide mass fingerprinting was used for the identification and subtyping of E. cloacae. Eighty-seven strains, identified based on hsp60 genotyping, were used to construct and evaluate a new reference database.Results Compared with the original reference database, the identification efficiency and accuracy of the new reference database was greatly improved at the species level. The first super reference database for E. cloacae identification was also constructed and evaluated. Based on the super reference database and the main spectra projection dendrogram, E. cloacae strains were divided into two clades.Conclusion Peptide mass fingerprinting is a powerful method to identify and subtype E. cloacae, and the use of this method will allow us to obtain more information to understand the heterogeneous organism E. cloacae.
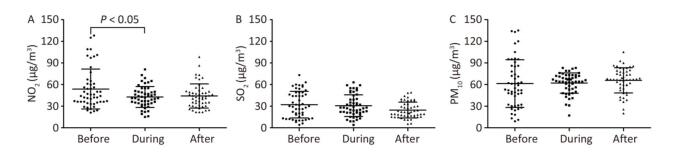
This three-year study, based on the Guangzhou Institute of Respiratory Disease (GRID), chronic obstructive pulmonary disease (COPD) Biobank, was conducted in 36 COPD patients to estimate whether changes in levels of leukocytes, erythrocytes, hemoglobin, and platelets were related to changes in air pollutant concentration. Daily NO2 levels exhibited significant differences between baseline years and the 2010 Asian Game period. We observed significant reductions in leukocyte and neutrophils counts levels, by 15.51% and 23.01%, from pre-Asian Games to during-Asian Games, respectively. In the post-Asian Game period, most pollutants approximated pre-Asian Game period levels, and similar effects were demonstrated in leukocyte and neutrophil counts. For both items, we identified significant increases resulting from elevated NO2 at lag days 0-2/5-6. We concluded that reductions in pollutants during the intervention period were associated with inactivation of hematological events in COPD.
To examine accuracy of body composition predicted by the Multi-Frequency Bioelectric Impedance Analysis (MF-BIA) compared with the Dual-energy X-ray Absorptiometry (DXA) in adults with obese. We measured body composition of 749 adults with obese both by the MF-BIA and DXA. The Lin's concordance correlation and the Bland-Altman plots were used to examine the consistency. The concordance correlation coefficient of %BF between the MF-BIA and DXA in men and women was 0.560, and 0.669, respectively. Compared with the DXA, the MF-BIA significantly underestimated %BF by 4.33% in men (P < 0.001), however overestimated %BF by 0.50% in women (P < 0.001). After corrected by the correction equations established in this study, the differences were significantly decreased. Therefore, the MF-BIA (TANITA MC-180) may need to be corrected in estimating body composition for adults with obese.
The molecular pathogenesis of T-2 toxin-induced cartilage destruction has not been fully unraveled yet. The aim of this study was to detect changes in serum metabolites in a rat anomaly model with articular cartilage destruction. Thirty healthy male Wistar rats were fed a diet containing T-2 toxin (300 ng/kg chow) for 3 months. Histopathological changes in femorotibial cartilage were characterized in terms of chondrocyte degeneration/necrosis and superficial cartilage defect, and the endogenous metabolite profile of serum was determined by UPLC/Q-TOF MS. Treated rats showed extensive areas of chondrocyte necrosis and superficial cartilage defect in the articular cartilage. In addition, 8 metabolites were found to change significantly in these rats compared to the control group, including lysoPE (18:0/0:0), lysoPC(14:0), lysoPC[18:4 (6Z, 9Z, 12Z, 15Z)], lysoPC[(16:1(9Z)], lysoPC(16:0), L-valine, hippuric acid, and asparaginyl-glycine. These 8 metabolites associated with cartilage injury are mainly involved in phospholipid and amino acid metabolic pathways.
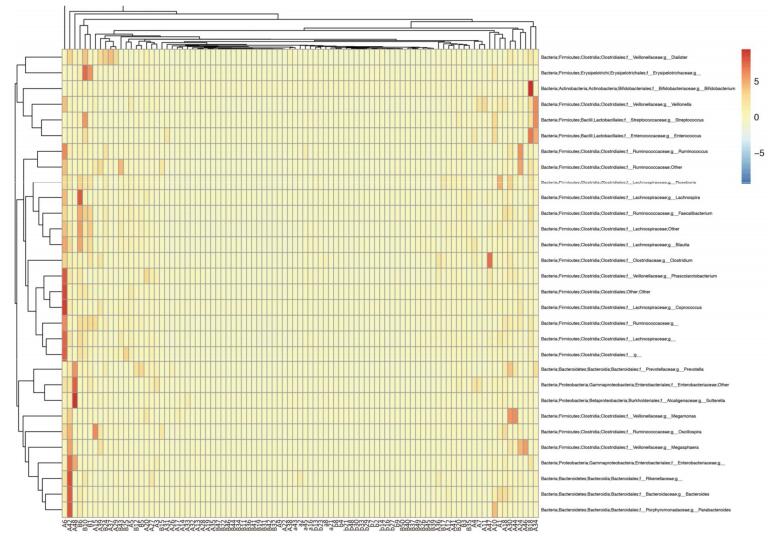
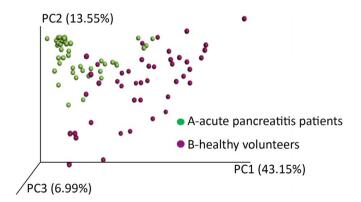
A case control study including 45 acute pancreatitis and 44 healthy volunteers was performed to investigate the association between intestinal microbial community and acute pancreatitis. High-throughput 16S rRNA gene amplicon sequencing was used to profile the microbiological composition of the samples. In total, 27 microbial phyla were detected and the samples of pancreatitis patients contained fewer phyla. Samples from acute pancreatitis patients contained more Bacteroidetes and Proteobacteria and fewer Firmicutes and Actinobacteria than those from healthy volunteers. PCoA analyses distinguished the fecal microbial communities of acute pancreatitis patients from those of healthy volunteers. The intestinal microbes of acute pancreatitis patients are different from those of healthy volunteers. Modulation of the intestinal microbiome may serve as an alternative strategy for treating acute pancreatitis.































 Quick Links
Quick Links