

 下载:
下载:

-
Prion diseases are characterized by neurodegeneration and protein aggregation, which are caused by the accumulation of a misfolded and protease-resistant form of the cellular prion protein. Several studies suggest that the accumulation of a misfolded alternate cellular protein disturbs quality control mechanisms, leading to endoplasmic reticulum (ER) stress involved in prion diseases[1-3]. Prion infection activates the splicing of the unfolded protein response transcription factor XBP-1. Misfolded mutant PrP associated with inherited accumulation in the ER may also affect the activity of proteins controlling the influx/efflux of calcium and decrease ER calcium content[1-4]. These events may alter cellular protein folding and quality control mechanisms to generate ER stress. Under cellular stress, cap-dependent translation is inhibited. On the other hand, protein translation is regulated by an internal ribosome entry site (IRES)-mediated cap-independent mechanism. Thus, prion infection may induce IRES-mediated cap-independent initiation of translation. Up-regulation of 14-3-3 proteins, which regulate multiple pivotal points in the cellular life cycle, are found in cerebrospinal fluid prion diseases, as well as being abundant in the brain[5-15]. This implies that 14-3-3 cap-independent initiation translation may be induced by ER stress during prion infection.
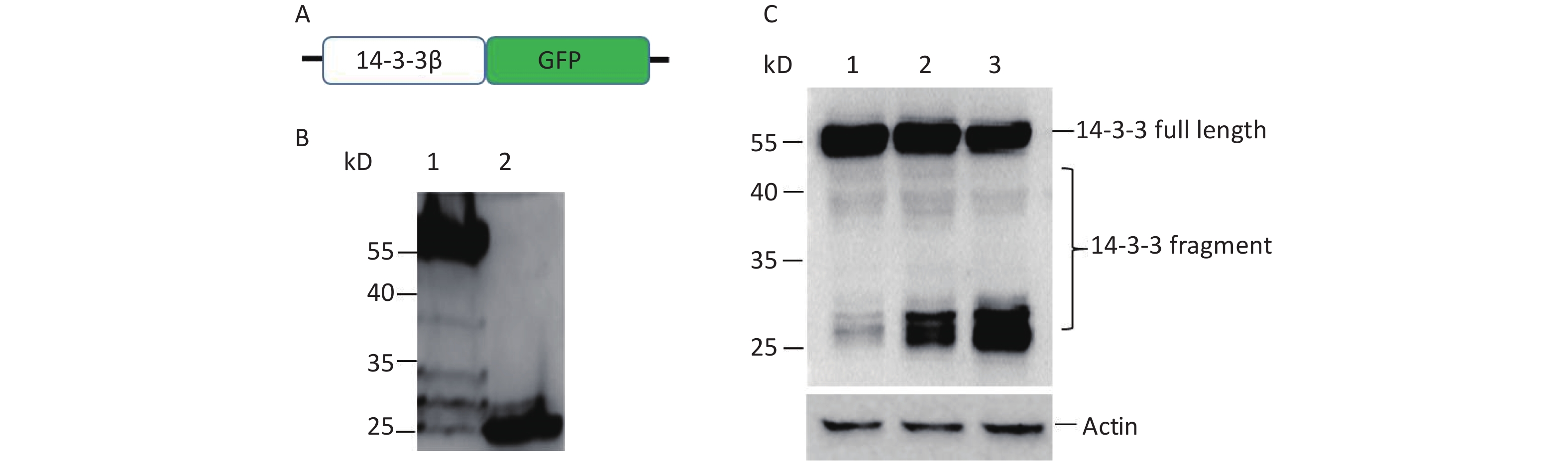
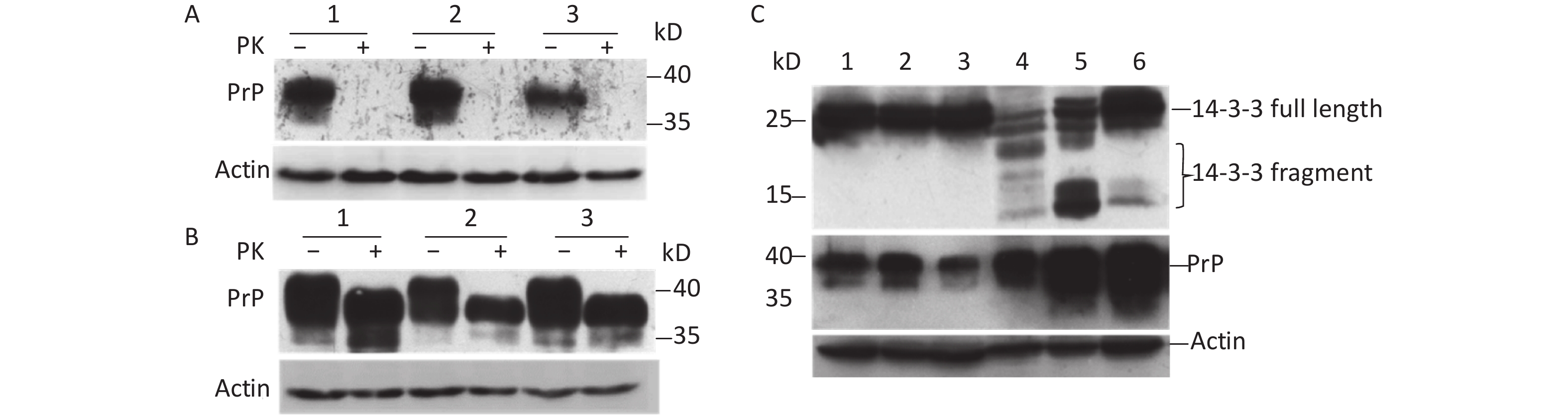
To understand whether prion infection induces 14-3-3 protein cap-independent translation initiation, scrapie 263K-infected hamster brain tissue was used in this study. Our results showed that normal prion protein (PrPC) could be completely digested by proteinase K (PK; Figure 1A). However, abnormal prion protein (PrPSc) from scrapie 263K-infected hamster brain had partial resistance to PK (Figure 1B). After exposure to 50 µg/mL PK for 30 min, the molecular weight of the protein became smaller (Figure 1B, lanes 4–6). The expression of PrP protein markedly increased compared with that of normal brain tissue (Figure 1C). Subsequently, 14-3-3β in scrapie 263K-infected hamster brain tissue was detected with a C-terminal antibody against 14-3-3β. The results showed that specific full-length 14-3-3 protein was detected in both the normal (Figure 1C, lanes 1–3) and infected brain tissues (Figure 1C, lanes 4–6). However, several lower molecular-weight bands appeared in the scrapie 263K-infected brain tissues (Figure 1C, lanes 4–6) but not in normal brain tissues (Figure 1C, lanes 1–3). These different molecular-weight 14-3-3β proteins in 263K-infected hamster brain tissue suggested that there were IRESs inside the open reading frame (ORF) of 14-3-3β genome to initiate cap-independent protein translation.
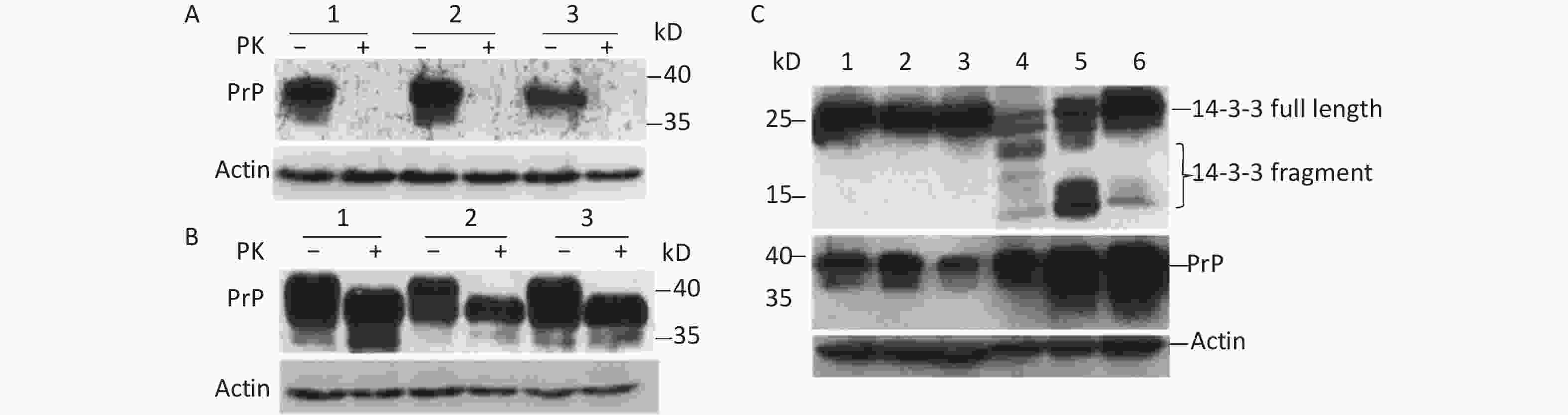
Figure 1. Cap-independent translation of 14-3-3β appeared in scrapie-infected hamster brain. (A) Prion protein was detected in three normal hamster brain tissues. (B) Prion protein was detected in three scrapie 263K-infected hamster brain tissues. (C) 14-3-3 protein and prion protein were detected in three normal hamster brain tissues (lanes 1−3) and in three scrapie 263K-infected hamster brain tissues (lanes 4−6). 14-3-3 protein and prion protein were detected by specific antibody’s Western blotting. The protein molecular-weight marker is indicated. ‘–’, no PK, ‘+’, PK.
To confirm the cap-independent translation inside ORF of 14-3-3β, full-length 14-3-3β mRNA was inserted into plasmid pEGFP-N1 with a green fluorescent protein (GFP) reporter gene to form p14-3-3 (Figure 2A). If there exists a single IRES in 14-3-3 mRNA, two fusing protein bands will be detected by GFP antibody’s Western blotting. The larger band is translated by cap-dependent initiation, the other is translated by IRES initiation. After transfection with p14-3-3β, 5 bands of about 55 kD, 38 kD, 32 kD, 30 kD, and 27 kD were detected by a GFP antibody’s Western blotting (Figure 2B, lane 1). Among them, the cap-dependent translational protein appears as a 55 kD fusion protein in the highest position (Figure 2B, lane 1). The other four bands were IRES-dependent translational proteins, and the expression of these fragments increased with the extension of transfection time from 12 h to 24 h (Figure 2C).
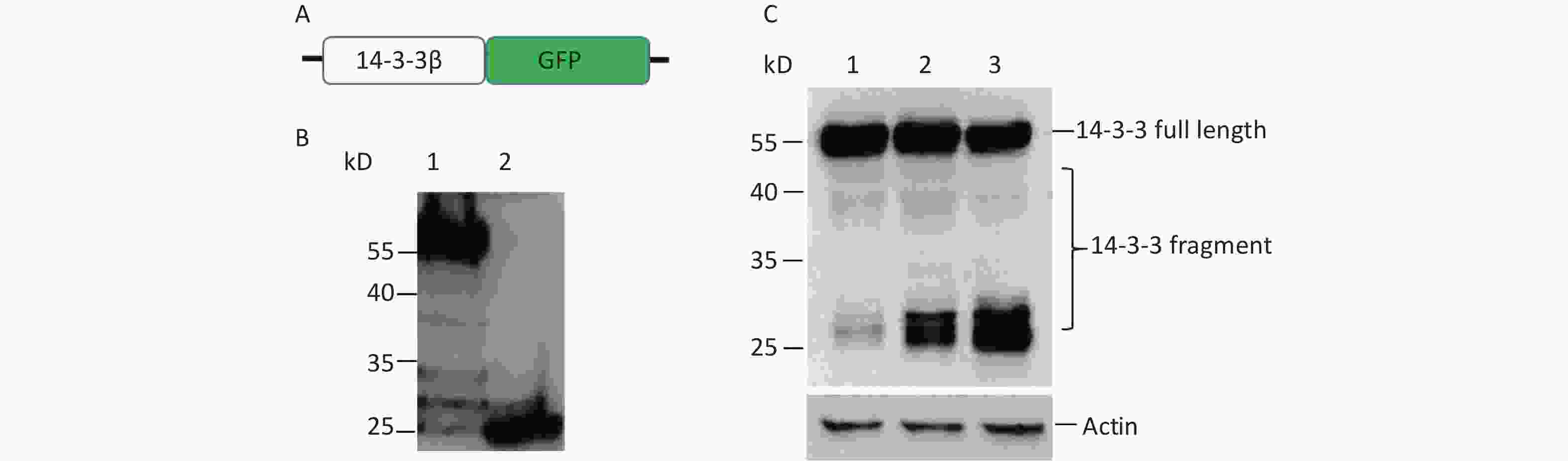
Figure 2. Cap-independent translation of 14-3-3β appeared in BHK cells. (A) Schematic diagram of IRES screen vectors with GFP fusion protein of 14-3-3β. Sequences from each ATG to 738 nt of 14-3-3β mRNA were serially cloned into pEGFP-N1. (B) Full-length protein and fragments of 14-3-3β were detected by a GFP antibody’ Western blotting. After BHK cells were transfected with GFP reporter plasmid p14-3-3(1-738), multiple 14-3-3β bands were detected around 55, 38, 32, 30, and 27 kD (lane 1). The plasmid pEGFP-N1 was used as a control (lane 2). (C) IRES-dependent translation amount increased with transfection time from 12 h (lane 1), 18 h (lane 2), to 24 h (lane 3). The protein molecular-weight marker is indicated. All the experiments were repeated no fewer than three times.
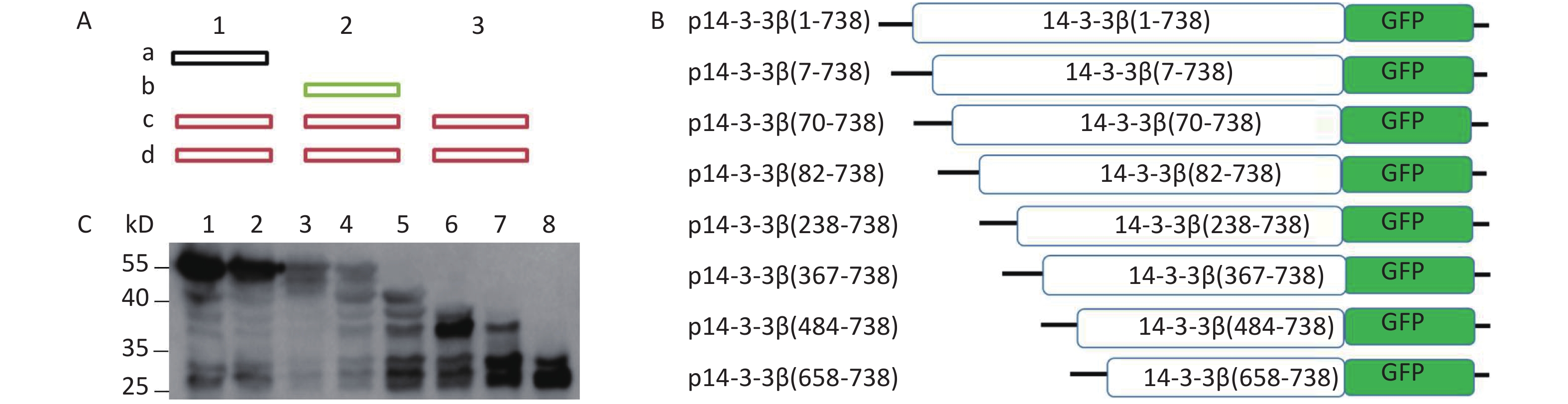
To determine the precise position of the IRES, eight sequences [7 start code (AUG)] from each start code to 3’-terminal of 14-3-3β were respectively introduced into pEGFP-N1 to form eight plasmids: p14-3-3β(1-738), p14-3-3β(7-738), p14-3-3β(70-738), p14-3-3β(82-738), p14-3-3β(238-738), p14-3-3β(367-738), p14-3-3β(484-738), and p14-3-3β(658-738) (Figure 3). The number of protein bands apparent by western blotting depends on the number of IRESs present within 14-3-3β mRNA. Following transfection, expressed 14-3-3 protein from eight plasmids was separated by SDS-PAGE and analyzed by Western blotting (Figure 3). We judged unambiguously that there were six candidate IRESs within 14-3-3β mRNA (Figure 3). The results further confirmed that multiple IRESs exist within 14-3-3 mRNA.

Figure 3. Investigation of IRES-dependent translation of 14-3-3β through Western blotting. (A) Schematic diagram of Western blotting ladder of IRES identification by GFP-fusing protein. Sequences from first ATG to 3’-terminal of gene will be serially fused into 3’-terminal of tag protein (The ideal tag protein should be greater than 15 kD). After transfection, brands dependent on IRESs will be identified by tag protein antibody’s Western blotting. IRES independent brand only appears in one lane (lane 2, b), not in the lane 1. There were IRES sites in the sequence of vectors, the same brand (c) will appear in the all lanes (c, d). (B) Schematic diagram of pEGFP-N1 inserted with sequences from each ATG to the end of 14-3-3. (C) IRESs of 14-3-3β mRNA were identified using a GFP antibody’s Western blotting. BHK cells were transfected with plasmids p14-3-3β(1-738) (lane 1), p14-3-3β(7-738) (lane 2), p14-3-3β(70-738) (lane 3), p14-3-3β(82-738) (lane 4), p14-3-3β(238-738) (lane 5), p14-3-3β(367-738) (lane 6), p14-3-3β(484-738) (lane 7), and p14-3-3β(658-738) (lane 8). The protein molecular-weight marker is indicated on the left. All experiments were repeated no fewer than three times.
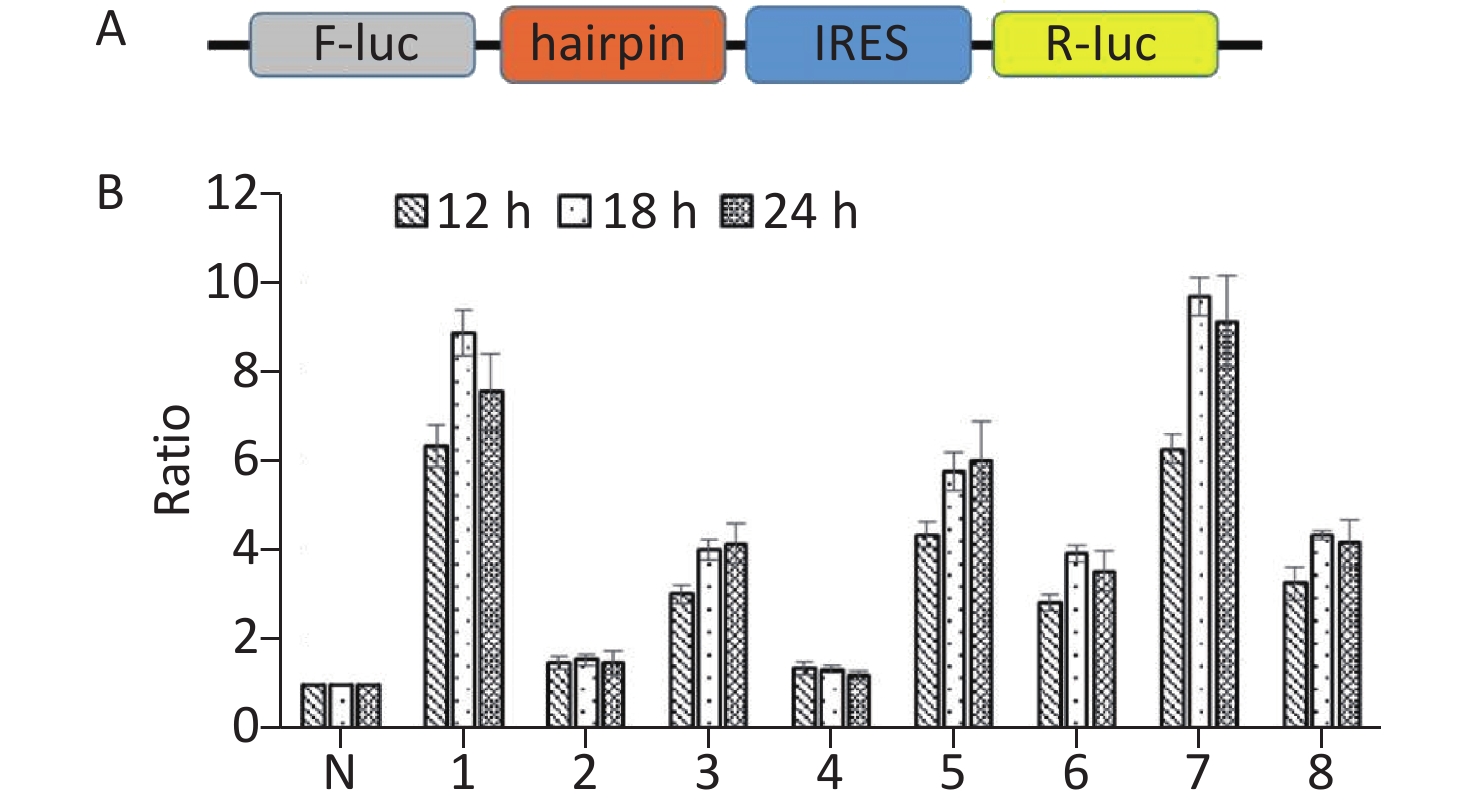
To further confirm the above results, we constructed bicistronic constructs with a hairpin between the Renilla luciferase (R-luc) and Firefly luciferase (F-luc) genes. The potential IRES sequences were inserted between the hairpin and the F-luc gene of the bicistronic constructs. The hairpin can prevent ribosome read‐through the first cistron to the second cistron. The upstream cistron R-luc is translated in a cap‐dependent manner, whereas the downstream F-luc will be translated and detected if the intercistronic sequence contains an IRES (Figure 4A). Subsequently, seven IRES candidates were respectively inserted in the bicistronic constructs to form plasmids p14-3-3-IRES-(1-6), p14-3-3-IRES-(7-69), p14-3-3-IRES-(70-81), p14-3-3-IRES-(82-237), p14-3-3-IRES-(238-366), p14-3-3-IRES-(367-483), p14-3-3-IRES-(484-657), and p14-3-3-IRES-(657-735). After transfection for 12 h, 18 h, and 24 h, p14-33-IRES-(1-6), p14-33-IRES-(70-81), p14-3-3-IRES-(238-366), p14-3-3-IRES-(367-483), p14-3-3-IRES-(484-657), and p14-3-3-IRES-(657-735). All plasmids had expressed higher F‐luc activity than that of control plasmid (Figure 4B). These results further confirmed the presence of IRES elements within the ORF of 14-3-3β. Combined with the results of Western blotting, we conclude that the IRES regions were located between 1st nt -6st nt, 70st nt -81st nt, 238st nt -366st nt, 367st nt -483st nt, 484st nt -657st nt, and 657st nt -735st nt of the 14-3-3 mRNA.
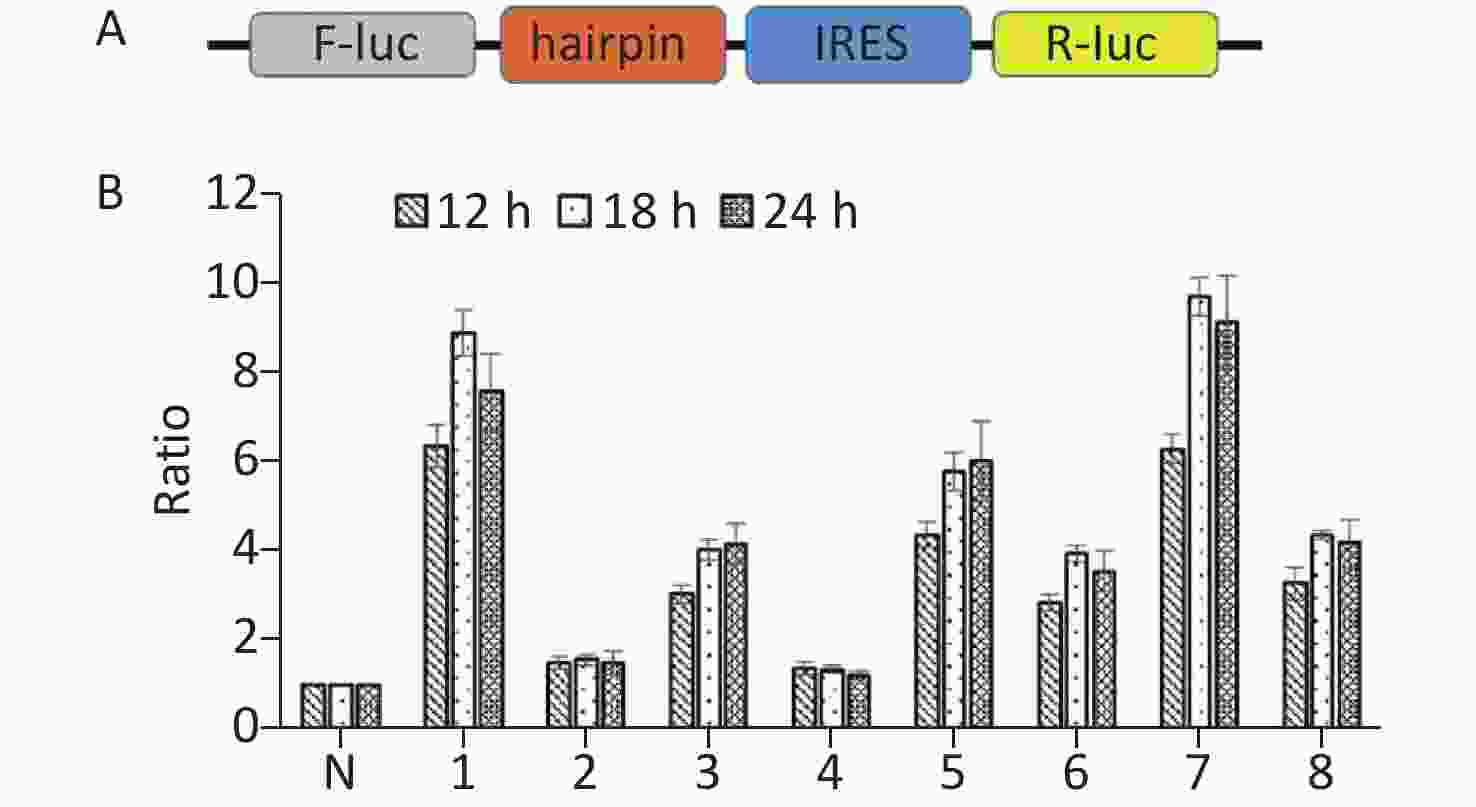
Figure 4. Identification of IRES sequences within 14-3-3β through bicistronic vectors. (A) Schematic diagram of bicistronic vectors for IRES identification. A bicistronic vector was constructed with a hairpin structure. The hairpin structure between R-luc and the IRES sequence prohibits ribosome read-through from R-luc to the F-luc cistron. (B) IRESs within 14-3-3 mRNA were identified using a bicistronic vector. Following transfection of BHK cells with p14-3-3-IRES-(1-6), p14-3-3-IRES-(7-69), p14-3-3-IRES-(70-81), p14-3-3-IRES-(82-237), p14-3-3-IRES-(238-366), p14-3-3-IRES-(367-483), p14-3-3-IRES-(484-657), p14-3-3-IRES-(657-735) for 12 h, 18 h, and 24 h, the activity ratio between F-luc and R-luc was calculated from each plasmid and compared with that of control plasmid. All experiments were repeated no fewer than three times.
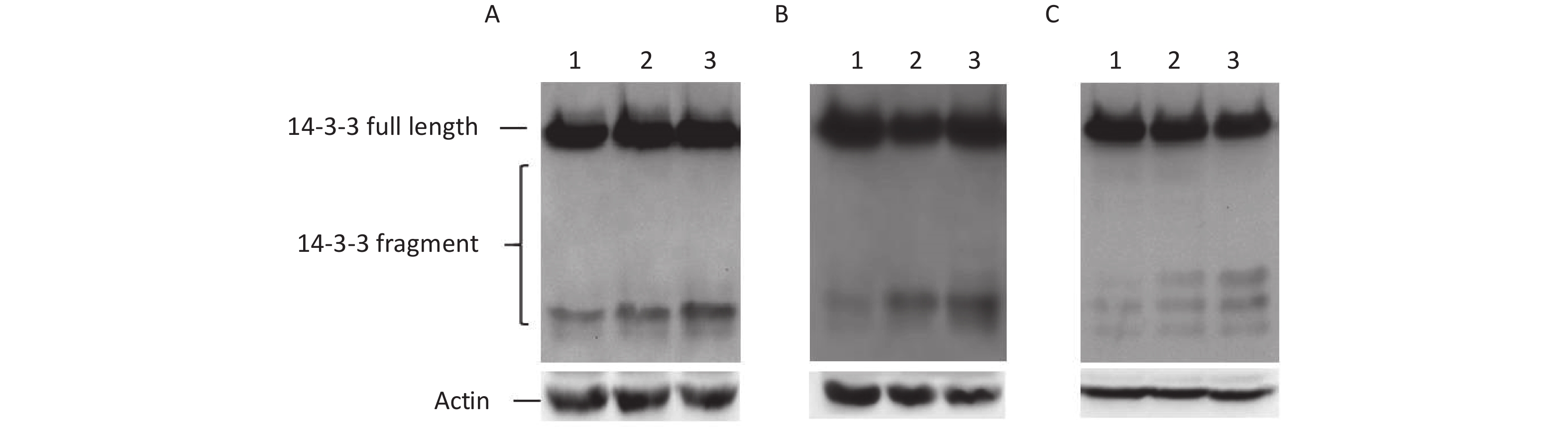
To understand the effect of ER stress on IRES-mediated initiation of 14-3-3β translation, three ER stress inducers, tunicamycin, thapsigargin or baflin A, were used in this study. Three drugs induced ER stress using their own way. tunicamycin interferes with protein folding in the ER by blocking the glycosylation of Asn residues of newly synthesized proteins[16]. Brefeldin A inhibits ER-Golgi vesicular transport leading to the accumulation of misfolded and unfolded proteins[17]. Thapsigargin induces ER stress by depleting ER Ca2+ stores[18]. Subsequently, BHK cells transfected with p14-3-3β(1-738) were treated with three inducers. We found that these inducers, brefeldin A, tunicamycin, and thapsigargin, clearly encouraged some IRES-mediated 14-3-3β protein expression, such as that of p14-3-3β(367-738), p14-3-3β(484-738), and p14-3-3β(658-738) (Figure 5). The results further demonstrated that cellular ER stress induces IRES-mediated 14-3-3β translation.
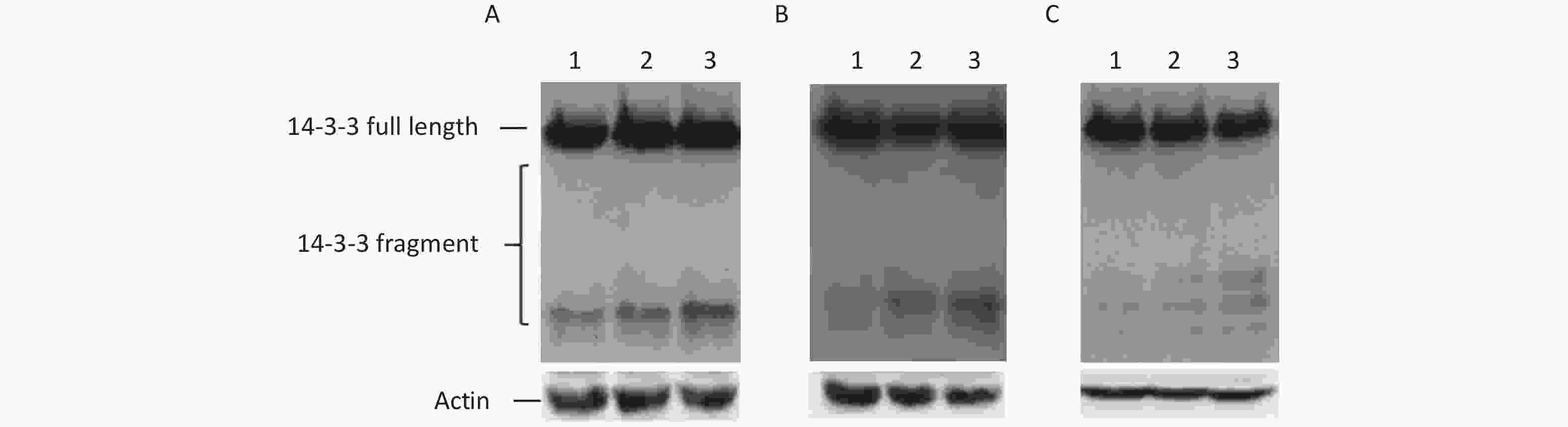
Figure 5. IRES-dependent 14-3-3 protein translation appeared during ER Stress. IRESs within 14-3-3 mRNA were regulated by tunicamycin, thapsigargin, or baflin A. After transfection with the plasmid p14-3-3β (1-735), BHK cells were exposed to tunicamycin. (A) (0 μg/mL lane 1, 10 μg/mL lane 2, 100 μg/mL, lane 3), baflin A. (B) (0 μmol/L lane 1, 10 μmol/L lane 2, 100 μmol/L, lane 3), or thapsigargin. (C) (0 μmol/L, lane 1, 0.1 μm lane 2, 0.2 μm, lane 3) for 24 h respectively, IRES-dependent 14-3-3 protein translation was identified by a GFP antibody following western blotting. All experiments were repeated no fewer than three times.
In conclusion, our results show that prion infection induces IRES-dependent 14-3-3 protein translation in the 263K-infected hamster brain. Our results also found there were six IRES regions inside the open reading frame (ORF) of 14-3-3β. These results nevertheless confirmed that 14-3-3β, as a stress response gene, undergoes multiple types of protein translation during ER stress.
doi: 10.3967/bes2020.037
Identification of Internal Ribosomal Entry Site inside Open Reading Frame of 14-3-3β Gene
-
-
Figure 1. Cap-independent translation of 14-3-3β appeared in scrapie-infected hamster brain. (A) Prion protein was detected in three normal hamster brain tissues. (B) Prion protein was detected in three scrapie 263K-infected hamster brain tissues. (C) 14-3-3 protein and prion protein were detected in three normal hamster brain tissues (lanes 1−3) and in three scrapie 263K-infected hamster brain tissues (lanes 4−6). 14-3-3 protein and prion protein were detected by specific antibody’s Western blotting. The protein molecular-weight marker is indicated. ‘–’, no PK, ‘+’, PK.
Figure 2. Cap-independent translation of 14-3-3β appeared in BHK cells. (A) Schematic diagram of IRES screen vectors with GFP fusion protein of 14-3-3β. Sequences from each ATG to 738 nt of 14-3-3β mRNA were serially cloned into pEGFP-N1. (B) Full-length protein and fragments of 14-3-3β were detected by a GFP antibody’ Western blotting. After BHK cells were transfected with GFP reporter plasmid p14-3-3(1-738), multiple 14-3-3β bands were detected around 55, 38, 32, 30, and 27 kD (lane 1). The plasmid pEGFP-N1 was used as a control (lane 2). (C) IRES-dependent translation amount increased with transfection time from 12 h (lane 1), 18 h (lane 2), to 24 h (lane 3). The protein molecular-weight marker is indicated. All the experiments were repeated no fewer than three times.
Figure 3. Investigation of IRES-dependent translation of 14-3-3β through Western blotting. (A) Schematic diagram of Western blotting ladder of IRES identification by GFP-fusing protein. Sequences from first ATG to 3’-terminal of gene will be serially fused into 3’-terminal of tag protein (The ideal tag protein should be greater than 15 kD). After transfection, brands dependent on IRESs will be identified by tag protein antibody’s Western blotting. IRES independent brand only appears in one lane (lane 2, b), not in the lane 1. There were IRES sites in the sequence of vectors, the same brand (c) will appear in the all lanes (c, d). (B) Schematic diagram of pEGFP-N1 inserted with sequences from each ATG to the end of 14-3-3. (C) IRESs of 14-3-3β mRNA were identified using a GFP antibody’s Western blotting. BHK cells were transfected with plasmids p14-3-3β(1-738) (lane 1), p14-3-3β(7-738) (lane 2), p14-3-3β(70-738) (lane 3), p14-3-3β(82-738) (lane 4), p14-3-3β(238-738) (lane 5), p14-3-3β(367-738) (lane 6), p14-3-3β(484-738) (lane 7), and p14-3-3β(658-738) (lane 8). The protein molecular-weight marker is indicated on the left. All experiments were repeated no fewer than three times.
Figure 4. Identification of IRES sequences within 14-3-3β through bicistronic vectors. (A) Schematic diagram of bicistronic vectors for IRES identification. A bicistronic vector was constructed with a hairpin structure. The hairpin structure between R-luc and the IRES sequence prohibits ribosome read-through from R-luc to the F-luc cistron. (B) IRESs within 14-3-3 mRNA were identified using a bicistronic vector. Following transfection of BHK cells with p14-3-3-IRES-(1-6), p14-3-3-IRES-(7-69), p14-3-3-IRES-(70-81), p14-3-3-IRES-(82-237), p14-3-3-IRES-(238-366), p14-3-3-IRES-(367-483), p14-3-3-IRES-(484-657), p14-3-3-IRES-(657-735) for 12 h, 18 h, and 24 h, the activity ratio between F-luc and R-luc was calculated from each plasmid and compared with that of control plasmid. All experiments were repeated no fewer than three times.
Figure 5. IRES-dependent 14-3-3 protein translation appeared during ER Stress. IRESs within 14-3-3 mRNA were regulated by tunicamycin, thapsigargin, or baflin A. After transfection with the plasmid p14-3-3β (1-735), BHK cells were exposed to tunicamycin. (A) (0 μg/mL lane 1, 10 μg/mL lane 2, 100 μg/mL, lane 3), baflin A. (B) (0 μmol/L lane 1, 10 μmol/L lane 2, 100 μmol/L, lane 3), or thapsigargin. (C) (0 μmol/L, lane 1, 0.1 μm lane 2, 0.2 μm, lane 3) for 24 h respectively, IRES-dependent 14-3-3 protein translation was identified by a GFP antibody following western blotting. All experiments were repeated no fewer than three times.
-
[1] Ron D, Walter P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat Rev Mol Cell Biol, 2007; 8, 519−29. doi: 10.1038/nrm2199 [2] Hetz C, Lee AH, Gonzalez-Romero D, et al. Unfolded protein response transcription factor XBP-1 does not influence prion replication or pathogenesis. Proc Natl Acad Sci U S A, 2008; 105, 757−62. doi: 10.1073/pnas.0711094105 [3] Steele AD, Hetz C, Yi CH, et al. Prion pathogenesis is independent of caspase-12. Prion, 2007; 1, 243−7. doi: 10.4161/pri.1.4.5551 [4] Torres M, Encina G, Soto C, et al. Abnormal calcium homeostasis and protein folding stress at the ER: a common factor in familial and infectious prion disorders. Commun Integr Biol, 2011; 4, 258−61. [5] Aitken A. 14-3-3 proteins: a historic overview. Semin Cancer Bio, 2006; 16, 162−72. doi: 10.1016/j.semcancer.2006.03.005 [6] Aitken A, Baxter H, Dubois T, et al. Specificity of 14-3-3 isoform dimer interactions and phosphorylation. Biochem Soc Trans, 2002; 30, 351−60. doi: 10.1042/bst0300351 [7] Berg D, Holzmann C, Riess O. 14-3-3 proteins in the nervous system. Nat Rev Neurosci, 2003; 4, 752−62. doi: 10.1038/nrg1159-c1 [8] Giacometti S, Camoni L, Albumi C, et al. Tyrosine phosphorylation inhibits the interaction of 14-3-3 proteins with the plant plasma membrane H+-ATPase. Plant Biol (Stuttg), 2004; 6, 422−31. doi: 10.1055/s-2004-820933 [9] Obsil T, Obsilova V. Structural basis of 14-3-3 protein functions. Semin Cell Dev Biol, 2011; 22, 663−72. [10] Steinacker P, Aitken A, Otto M. 14-3-3 proteins in neurodegeneration. Semin Cell Dev Biol, 2011; 22, 696−704. [11] Takahashi Y. The 14-3-3 proteins: gene, gene expression, and function. Neurochem Res, 2003; 28, 1265−73. doi: 10.1023/A:1024296932670 [12] van Heusden GP. 14-3-3 proteins: regulators of numerous eukaryotic proteins. IUBMB Life, 2005; 57, 623−9. doi: 10.1080/15216540500252666 [13] Wang W, Shakes DC. Molecular evolution of the 14-3-3 protein family. J Mol Evol, 1996; 43, 384−98. doi: 10.1007/BF02339012 [14] Yaffe MB. How do 14-3-3 proteins work?-- Gatekeeper phosphorylation and the molecular anvil hypothesis. FEBS Lett, 2002; 513, 53−7. doi: 10.1016/S0014-5793(01)03288-4 [15] VanGuilder HD, Farley JA, Yan H, et al. Hippocampal dysregulation of synaptic plasticity-associated proteins with age-related cognitive decline. Neurobiol Dis, 2011; 43, 201−12. doi: 10.1016/j.nbd.2011.03.012 [16] Brewer JW, Diehl JA. PERK mediates cell-cycle exit during the mammalian unfolded protein response. Proc Natl Acad Sci U S A, 2000; 97, 12625−30. doi: 10.1073/pnas.220247197 [17] Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Gene Develop, 1999; 13, 1211−33. [18] Harding HP, Novoa I, Zhang Y, et al. Regulated translation initiation controls stress-induced gene expression in mammalian cells. Molecular cell, 2000; 6, 1099−108. doi: 10.1016/S1097-2765(00)00108-8 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1256
- HTML全文浏览量: 362
- PDF下载量: 65
- 被引次数: 0

 Quick Links
Quick Links