

 下载:
下载:



-
Vibrio cholerae naturally lives in aquatic environments and causes deadly cholera in humans[1]. V. cholerae biofilms play significant roles in virulence, persistence, transmission, and survival in adverse environments[2-5], and their formation requires specific structures, including flagella, exopolysaccharide, and pili[5]. Numerous factors, such as VpsR[6,7], VpsT[6,7], VxrB[8], H-NS[9], CRP[10], CarR[11], PhoB[12], Fur[13], quorum sensing (QS)[14,15], and cyclic diguanylate (c-di-GMP)[16], are required for biofilm regulation in V. cholerae.
Cyclic di-GMP (c-di-GMP) is diffusely applied by bacteria to modulate multiple behaviors, including motility, biofilm formation, virulence determinant production, DNA repair, and cell shape[16-18]. Increased c-di-GMP levels in bacterial cells induce biofilm formation, and decreased c-di-GMP concentrations induce a motile lifestyle[16,19]. c-di-GMP is catalyzed by diguanylate cyclase (DGC) harboring the GGDEF domain and is degraded by phosphodiesterase harboring the EAL or HD-GYP domain[16]. More than 50 genes encode GGDEF- and/or EAL-type proteins in V. cholerae[20], and about 50% of these genes contribute to c-di-GMP metabolism[10,19,21-33]. However, the c-di-GMP metabolic pool in V. cholerae warrants further investigation.
The VCA0560 gene encodes a GGDEF-type protein in V. cholerae[34]. VCA0560 overexpression reduces motility[35] but only slightly alters biofilm formation[36]. Whether the VCA0560 protein affects the intracellular c-di-GMP pool needs to be further investigated. Here, the data showed that VCA0560 gene deletion does not affect the c-di-GMP synthesis, biofilm formation, and motility. Meanwhile, VCA0560 overexpression enhances c-di-GMP production and biofilm formation but reduces swimming motility. In addition, the transcription of the VCA0560 gene is directly repressed by the master quorum sensing (QS) regulator HapR[37] and the ferric uptake regulator (Fur)[38]. In summary, overexpressed VCA0560 acts as an active DGC, and its transcription is repressed by Fur and HapR.
-
V. cholerae El Tor C7258 was applied as the wild-type (WT) strain. Nonpolar hapR and fur single-gene mutants (ΔhapR and Δfur) and the complementary mutants ΔhapR/pBAD24-hapR (C-ΔhapR) and Δfur/pBAD24-fur (C-Δfur) were constructed as previously described[39]. The VCA0560 gene mutant (ΔVCA0560) was constructed from WT using the suicide plasmid pWM91 by the allelic exchange as previously described[40]. The coding region of the VCA0560 gene was cloned into the pBAD24 vector containing an arabinose-inducing pBAD promoter to construct the overexpression plasmid pBAD24-VCA0560[41]. The recombinant pBAD24-VCA0560 was transferred into WT to construct the VCA0560 overexpression strain WT/pBAD24-VCA0560, which produced about 3.5 times as many VCA0560 mRNA levels as that of the WT/pBAD24 strain (data not shown). The pBAD24 vector was also introduced into WT, ΔhapR, and Δfur to prepare controls. All primers used are listed in Table 1.
Table 1. Oligonucleotide primers used in this study
Target Primers (forward/reverse, 5'–3’) Construction of mutant VCA0560 GCGGGATCCGAACAGCGATCAGAGCATG/CCTCCTTTGCCAATTTTGAATTCCTGAGGTTCTACATTG CAATGTAGAACCTCAGGAATTCAAAATTGGCAAAGGAGG/GGACTAGTGCGAGTTGATGTGGGTAG GCGGGATCCGAACAGCGATCAGAGCATG/GGACTAGTGCGAGTTGATGTGGGTAG Construction of overexpressed plasmid VCA0560 GATTCTAGAATGTCTTCTAGCTTTGTTAC/GCGAAGCTTTTAGCTAGCGACTTTGACAC Luminescence assay VCA0560 GCGGAGCTCGAACAGCGATCAGAGCAT/GCGGGATCCCAGCAGCAATAGAGGAAAA EMSA VCA0560 GAACAGCGATCAGAGCAT/CAGCAGCAATAGAGGAAAA recA AAGATTGGTGTGATGTTTGGTA/CACTTCTTCGCCTTCTTTGA DNase I footprinting VCA0560 GTAAAACGACGGCCAGTCAAGCTGCAACGGCAAGTAG/CAGGAAACAGCTATGACCAGCAGCAATAG
AGGAAAAM13 FAM-GTAAAACGACGGCCAGT/CAGGAAACAGCTATGAC-HEX -
Luria-Bertani (LB) broth (1% tryptone, 0.5% yeast extract, and 1% NaCl) was applied for V. cholerae cultivation[39]. Overnight bacterial cultures were diluted at 1:50 into 5 mL of fresh LB broth and grown at 30 °C with shaking at 200 rpm to an OD600 value of 1.0. The resultant cultures were diluted 1:100 into 5 mL of fresh LB broth for the third-round growth. The bacterial cells were harvested at an OD600 value of about 0.6. Whenever required, antibiotics were used in the following concentrations: 100 mg/mL ampicillin, 5 mg/mL chloramphenicol, and 0.1% arabinose.
-
Colony morphology was characterized as previously described[42]. In brief, 2 μL of overnight bacterial culture was spotted onto the LB agar and then statically incubated at 30 °C for 48 h.
-
CV staining was performed as previously described[42]. The third round of cultures was diluted 50-fold into 2 mL of fresh LB broth in glass tubes and then allowed to grow at 30 °C with shaking at 100 rpm for 48 h. The surface-attached cells were stained with 0.1% CV. The bound dye was dissolved with dimethylsulfoxide (DMSO), and the OD570 values were measured for each strain as the index of CV staining.
-
Intracellular c-di-GMP levels were detected as previously described[13,43]. Bacterial cells were harvested at OD600 = 0.6, washed twice with ice-cold phosphate buffered solution (PBS), resuspended in 2 mL of ice-cold PBS, incubated at 100 °C for 5 min, and sonicated for 15 min (power 100%, frequency 37 kHz) in an ice-water bath. The supernatant was collected, and the pellet was resuspended in 2 mL of ice-cold PBS and re-extracted twice. The intracellular c-di-GMP levels were determined with a c-di-GMP ELISA kit (Mskbio, Beijing, China). Total protein concentration in the supernatant was also determined using a Pierce BCA Protein Assay kit (ThermoFisher Scientific, USA) in accordance with the manufacturer’s instructions. c-di-GMP concentration was expressed as pmol/mg protein.
-
Swimming motility was assayed as previously described[44]. In brief, 2 μL of the third round of cell cultures were inoculated into semisolid swim plates (1% tryptone, 0.5% yeast extract, 1% NaCl, and 0.35% bacterial agar), and the diameter of the swimming areas was measured after incubation at 30 °C for about 10 h.
-
A luminescence assay was performed as previously described[45]. The regulatory DNA region of VCA0560 was cloned into the pBBRlux vector harboring a promoterless luxCDABE reporter gene. The recombinant plasmid was transferred into WT and regulatory gene mutants (ΔhapR and Δfur) to measure lux activity in each strain. Luminescence was determined using an Infinite® 200 Pro NanoQuant (Tecan, Switzerland), and lux activity was calculated as light units/OD600.
-
The coding regions of hapR and fur were separately cloned into the pET28a (Novagen, USA) to express His-HapR and His-Fur using Escherichia coli BL21λDE3 cells[46]. His-HapR and His-Fur were prepared as previously described [47,48]. The dialyzed proteins were concentrated to approximately 0.5 mg/mL, and their purity was analyzed by SDS-10% PAGE.
-
EMSA was performed as previously described[48]. The 5ʹ-ends of the regulatory DNA region of the VCA0560 gene were labeled with [γ-32P] ATP and T4 polynucleotide kinase. DNA binding was performed in a 10 µL reaction volume containing binding buffer (1 mmol/L MgCl2, 100 μmol/L MnCl2 or 0.5 mmol/L EDTA, 0.5 mmol/L DTT, 50 mmol/L NaCl, 10 mmol/L Tris-HCl/pH 7.5, and 10 mg/mL salmon sperm DNA), labeled DNA probe, and increasing amounts of purified His-Fur or His-HapR. After incubation at room temperature for 20 min, the products were loaded onto a native 4% (w/v) polyacrylamide gel and electrophoresed in 1.0× TB or 0.5× TBE buffer for approximately 60 min at 150 V. Radioactive species were detected by autoradiography after exposure to Fuji Medical X-ray film at −20 °C. Three controls were included in each EMSA experiment: 1) a cold probe as a specific DNA competitor (the same promoter DNA region unlabeled), 2) a negative probe as a nonspecific DNA competitor (the unlabeled coding region of the 16S rRNA gene), and 3) a nonspecific protein competitor (rabbit anti-F1-protein polyclonal antibodies).
-
DNase I footprinting was performed as previously described[49-51]. The results were analyzed using an ABI 3500XL DNA Genetic analyzer with GeneMarker software 2.2.
-
Colony morphological characterization, CV staining, EMSA, and DNase I footprinting assays were performed at least twice with similar results. Luminescence assay and c-di-GMP quantification were performed three times, and the values were expressed as mean ± standard deviation (SD). Paired Student’s t-tests were applied to calculate statistical significance. P values less than 0.01 were considered significant.
-
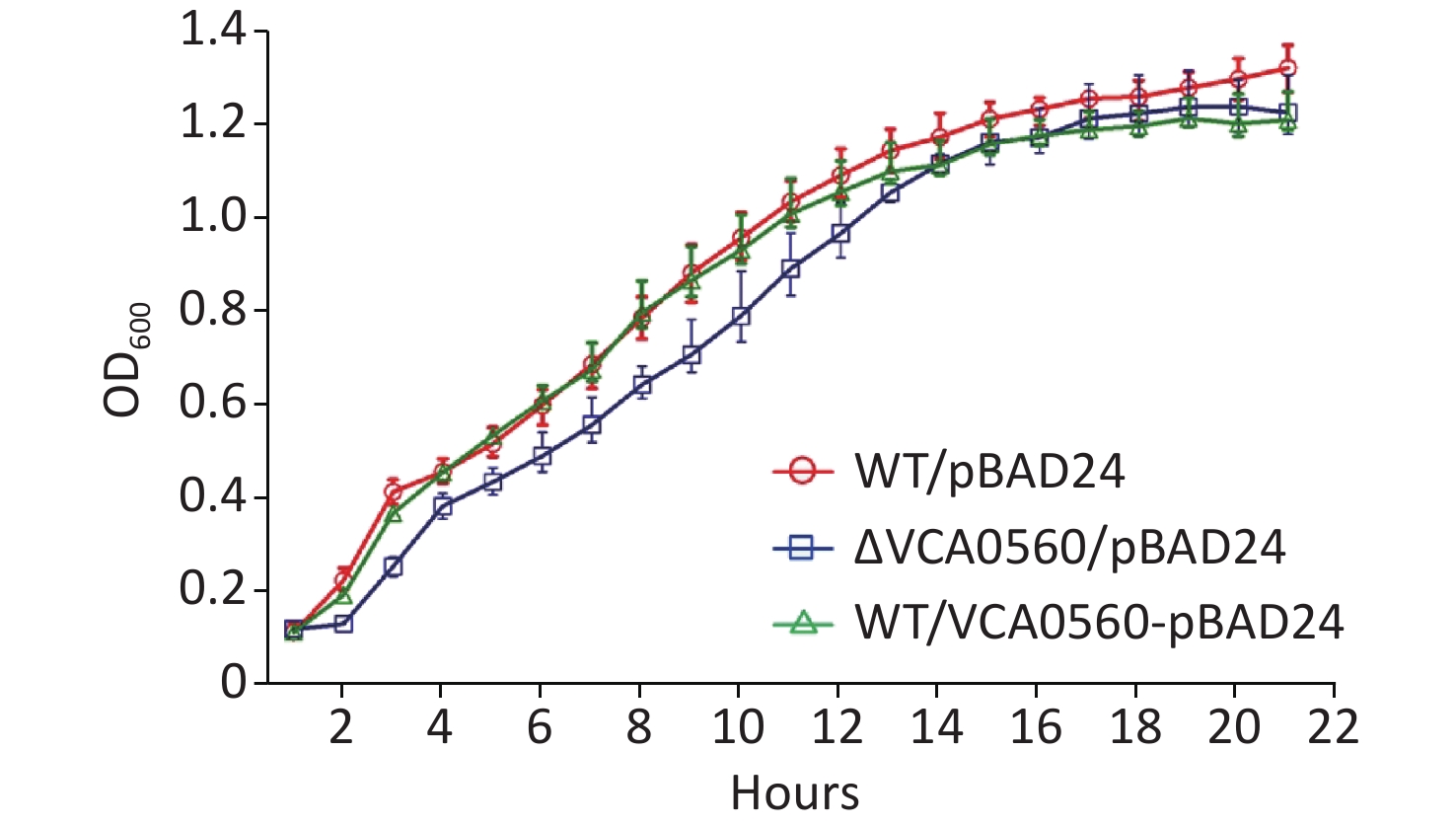
The growth curves of WT/pBAD24, ΔVCA0560/pBAD24, and WT/pBAD24-VCA0560 grown at 30 °C in LB broth were recorded with 1 h intervals to determine the effect of VCA0560 on the growth of V. cholerae. As shown in Figure 1, indistinguishable growth rates were observed for WT/pBAD24 and WT/pBAD24-VCA0560, indicating that VCA0560 overexpression did not affect the growth of V. cholerae. However, the ΔVCA0560/pBAD24 strain showed a slight growth defect relative to the WT/pBAD24 and WT/pBAD24-VCA0560 strains, suggesting that VCA0560 might be required for the growth of V. cholerae.
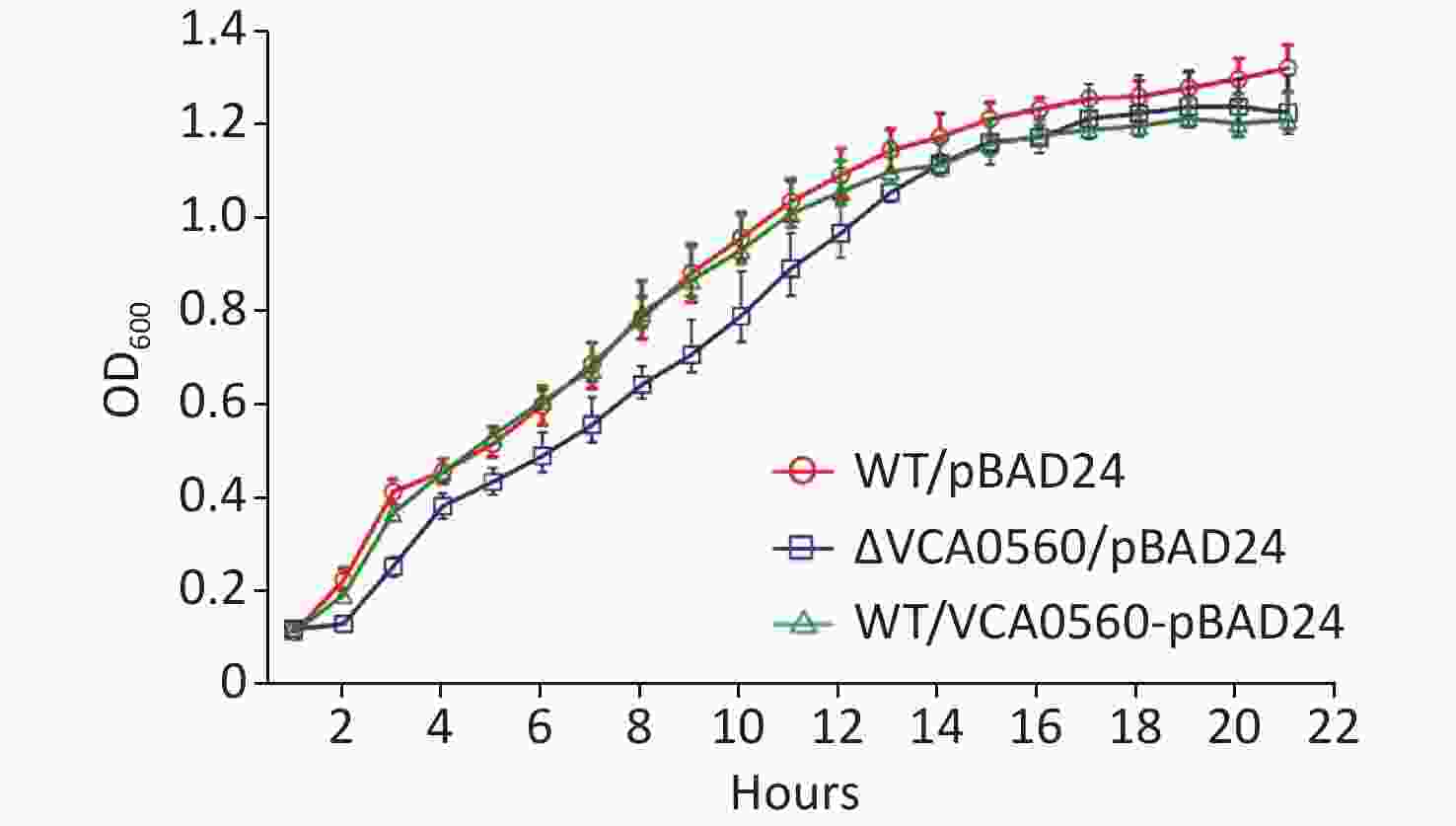
Figure 1. Growth curves of V. cholerae strains. The overnight growth cultures of WT/pBAD24, ΔVCA0560/pBAD24, and WT/pBAD24-VCA0560 were diluted at 1:100 into 20 mL of LB broth supplemented with 100 mg/mL ampicillin and 0.1% arabinose, mixed well, and then divided into a microwell plate with 300 μL in each well. V. cholerae strains were grown at 30 °C with shaking at 200 rpm, and the OD600 values were monitored at 1 h intervals. The experiment was conducted twice, with five replicates per trial for each strain.
-
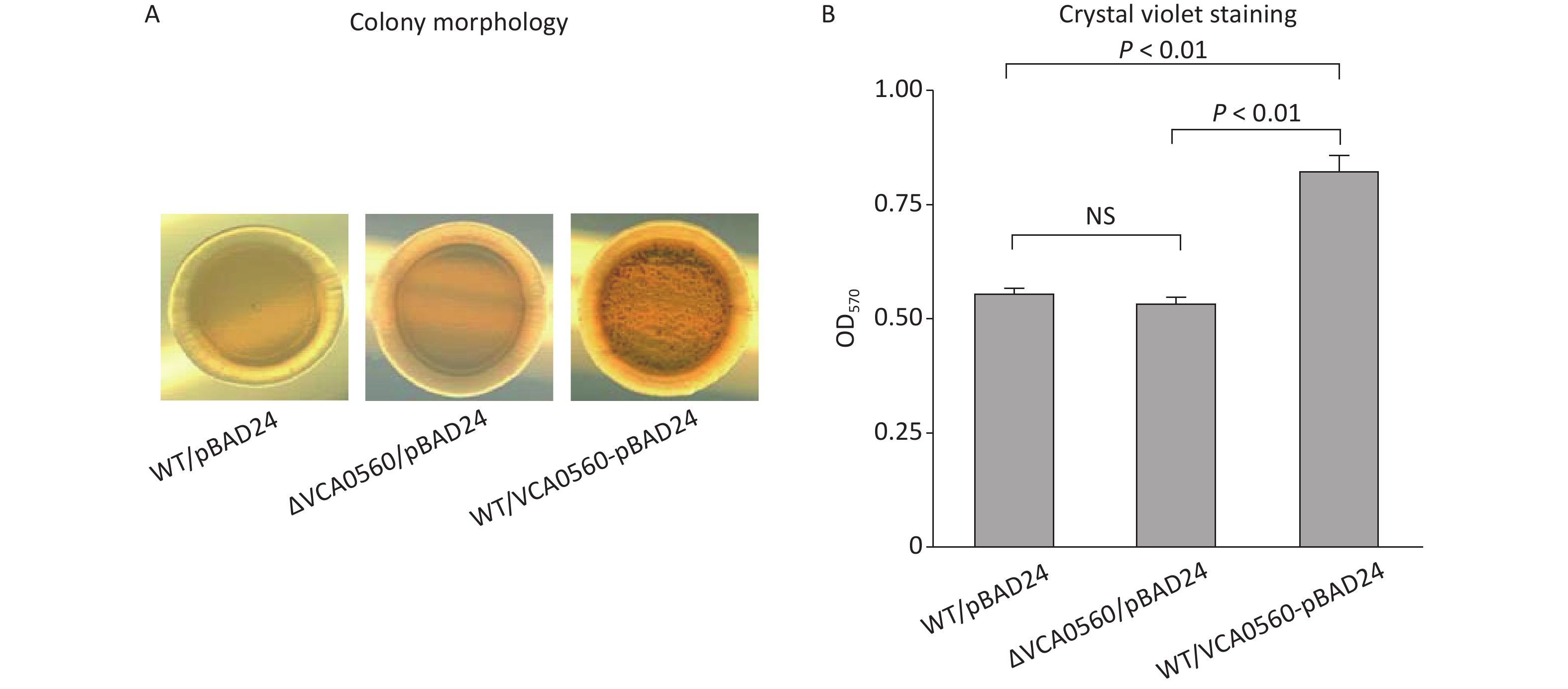
Colony morphological characterization and CV staining were performed to compare colony morphology and biofilm quantity among WT/pBAD24, ΔVCA0560/pBAD24, and WT/pBAD24-VCA0560. As shown in Figure 2A, WT/pBAD24 and ΔVCA0560/pBAD24 produced smooth colonies on the LB plates, and WT/pBAD24-VCA0560 formed wrinkled colonies. CV staining assay showed that WT/pBAD24-VCA0560 produced significantly more normalized CV staining than WT/pBAD24 and ΔVCA0560/pBAD24, and ΔVCA0560/pBAD24 showed similar CV staining result with WT/pBAD24 (Figure 2B). These results indicated that VCA0560 gene deletion had no regulatory activity on biofilm formation, but overexpression of the protein did induce biofilm formation by V. cholerae.
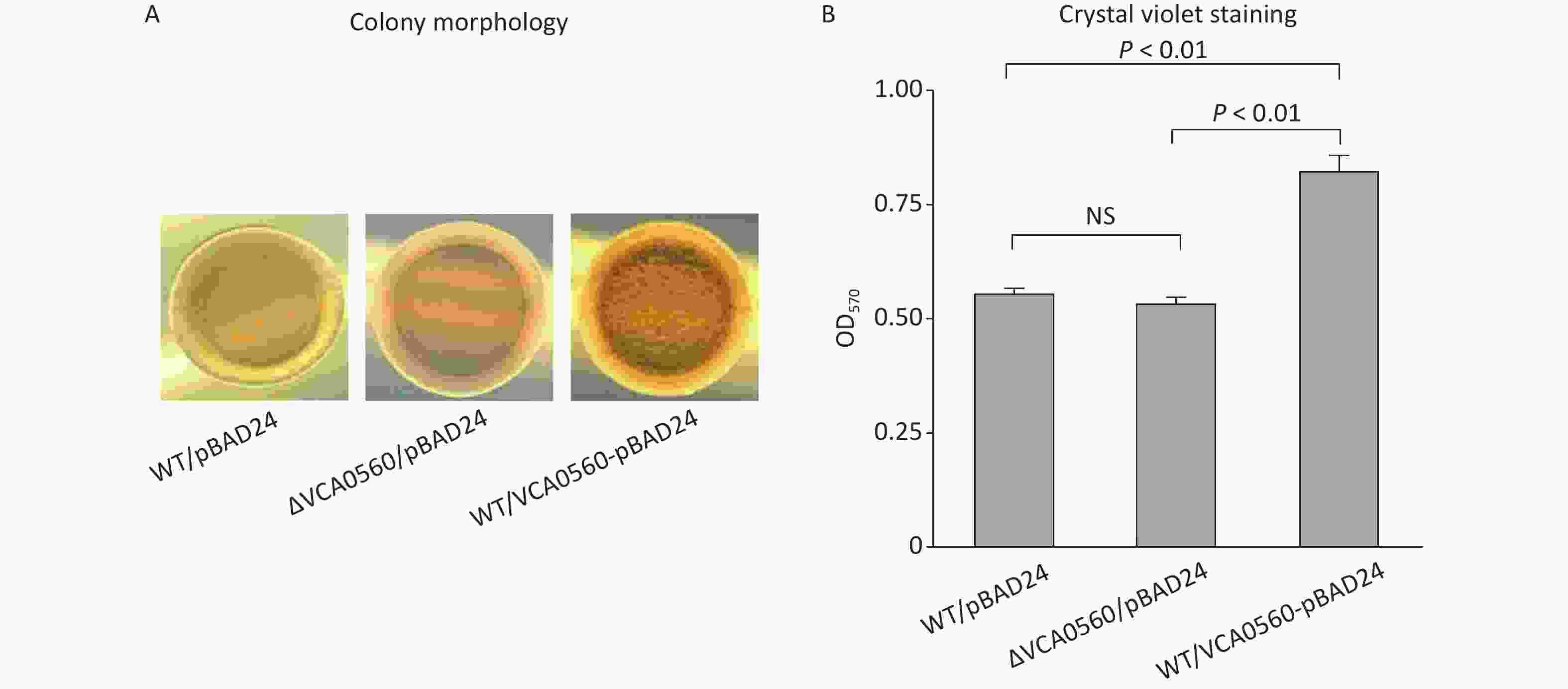
Figure 2. VCA0560 overexpression induced biofilm formation by V. cholerae. (A) Colony morphological characterization. In brief, 2 μL of overnight bacterial culture was spotted on the LB agar supplemented with 100 mg/mL ampicillin and 0.1% arabinose and then statically incubated at 30 °C for 48 h. (B) Crystal violet (CV) staining. Bacterial cells were inoculated into 2 mL of fresh LB broth supplemented with 100 mg/mL ampicillin and 0.1% arabinose in glass tubes and allowed to grow at 30 °C with shaking at 100 rpm for 48 h. The surface-attached cells were stained with 0.1% CV. The bound CV was dissolved with dimethylsulfoxide (DMSO), and the OD570 values were measured as the index of CV staining. NS represents no significant difference.
-
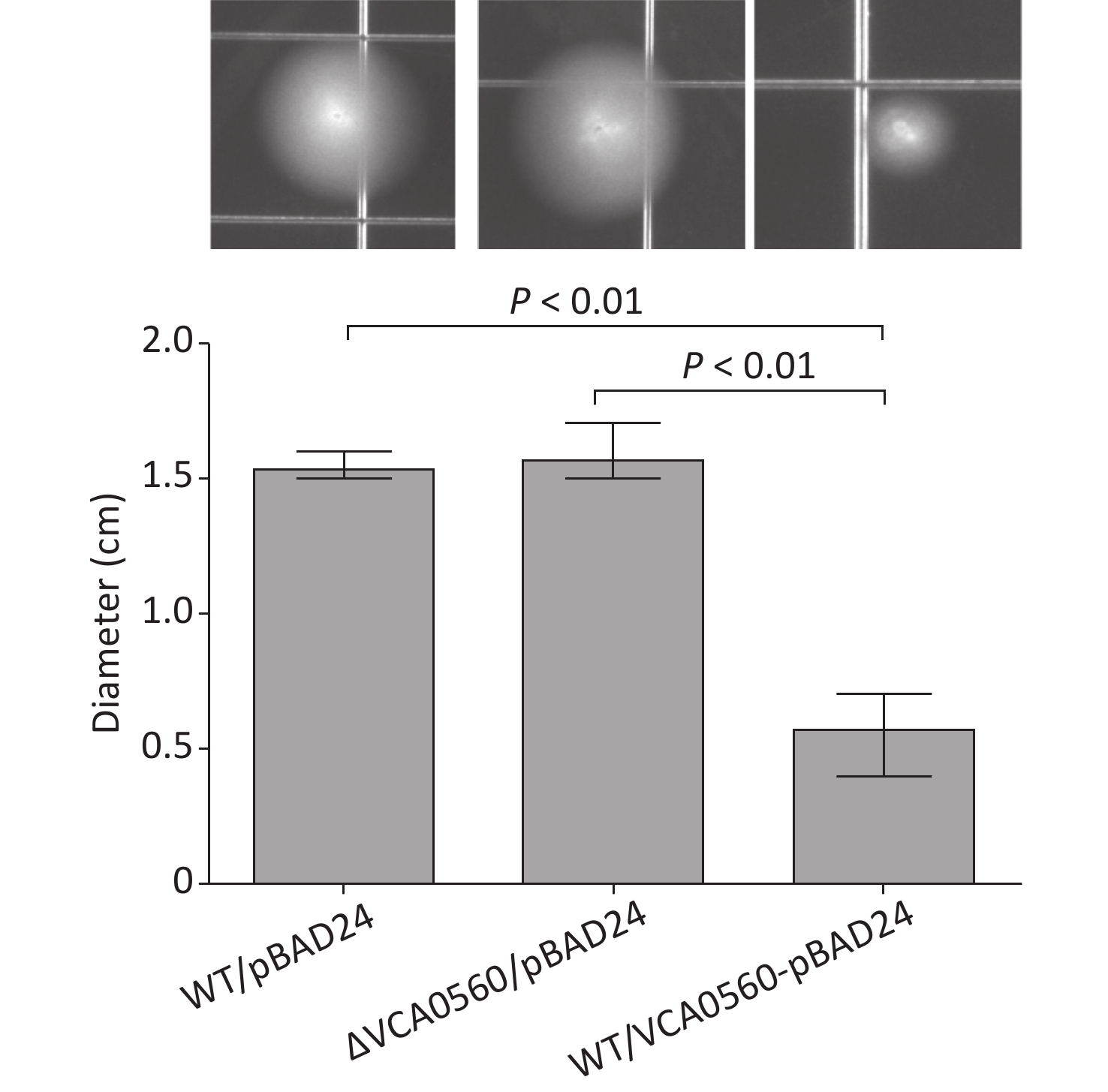
The swimming motility assay showed that the swimming capacity of WT/pBAD24-VCA0560 was significantly inhibited relative to those of WT/pBAD24 and ΔVCA0560/pBAD24. ΔVCA0560/pBAD24 and WT/pBAD24 manifested a similar capacity for swimming motility (Figure 3). These results suggested that VCA0560 overexpression inhibited V. cholerae swimming motility.
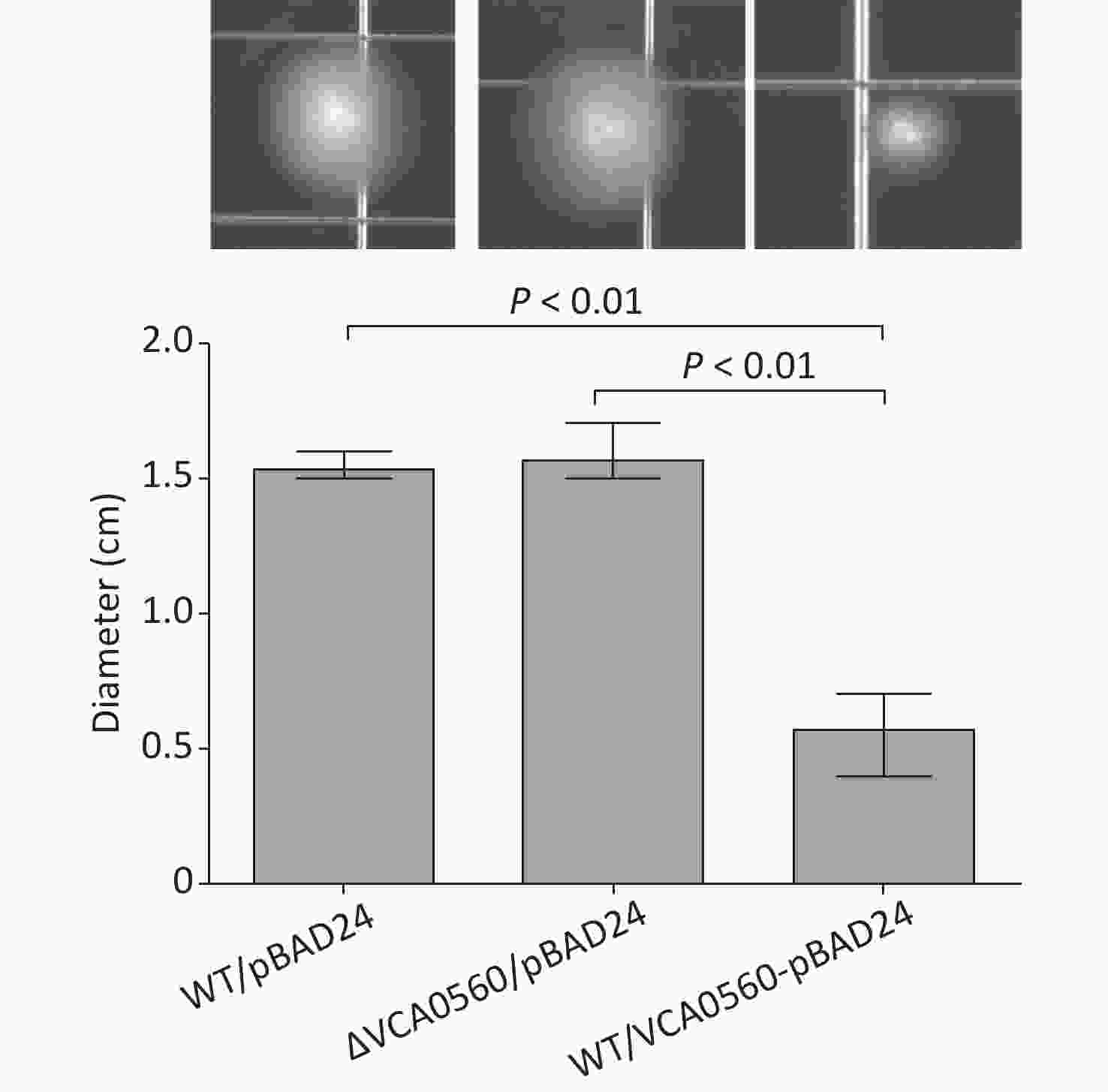
Figure 3. Regulation of swimming motility by VCA0560 protein in V. cholerae. In brief, 2 μL of bacterial cells were inoculated into semisolid swim plates (1% tryptone, 0.5% yeast extract, 1% NaCl, and 0.35% bacterial agar) supplemented with 100 mg/mL ampicillin and 0.1% arabinose. The diameter of the swimming areas was measured after incubation at 30 °C for 10 h.
-
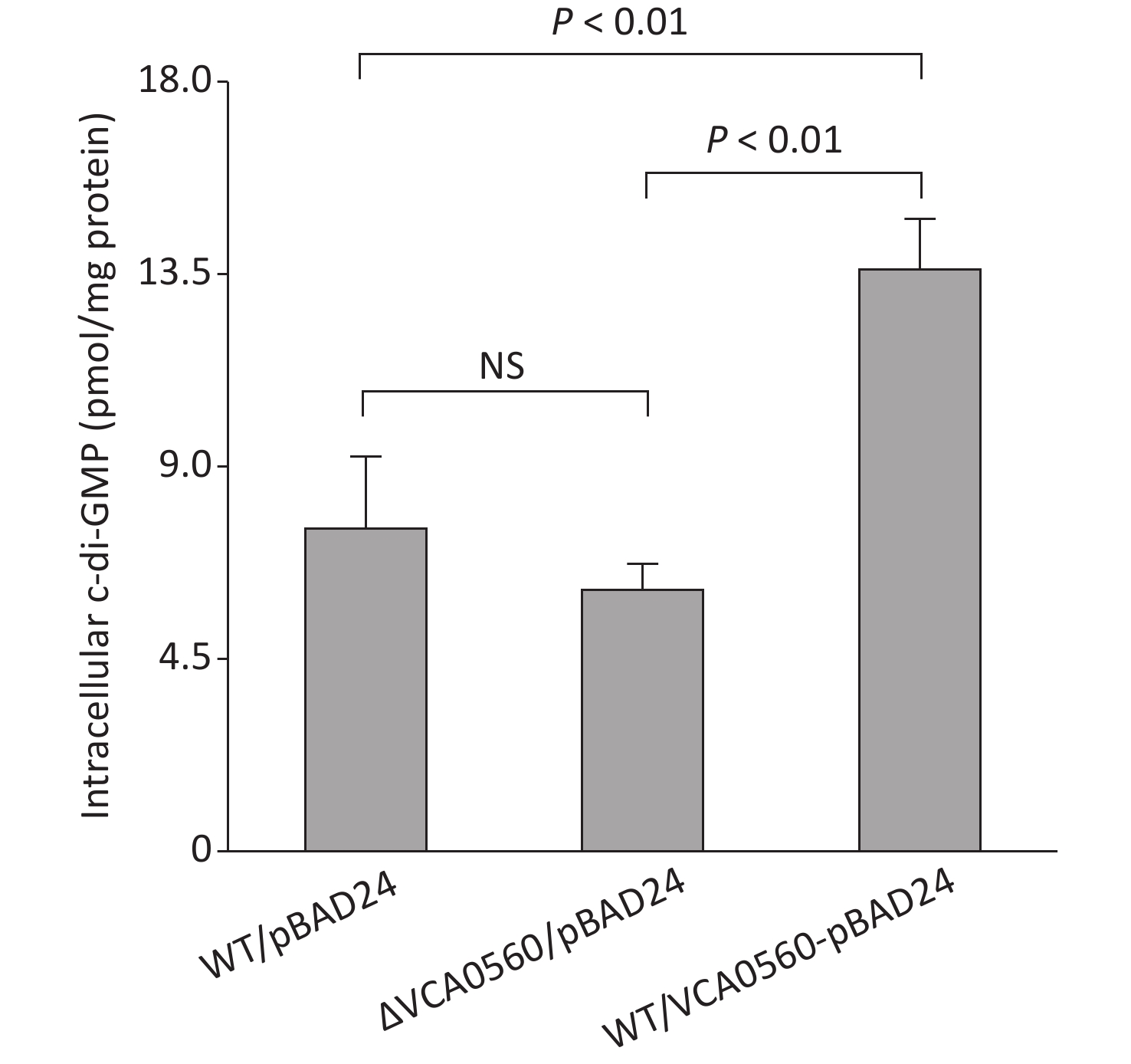
Elevated c-di-GMP levels in bacterial cells lead to decreased motility but increased biofilm formation[5]. Thus, we further detected whether VCA0560 regulates c-di-GMP production in V. cholerae. The concentration of c-di-GMP in WT/pBAD24-VCA0560 was higher than that in WT/pBAD24 and ΔVCA0560/pBAD24. Meanwhile, ΔVCA0560/pBAD24 had similar c-di-GMP levels to WT/pBAD24 (Figure 4). These results indicated that VCA0560 mutation did not influence c-di-GMP production, but VCA0560 overexpression induced c-di-GMP synthesis in V. cholerae.
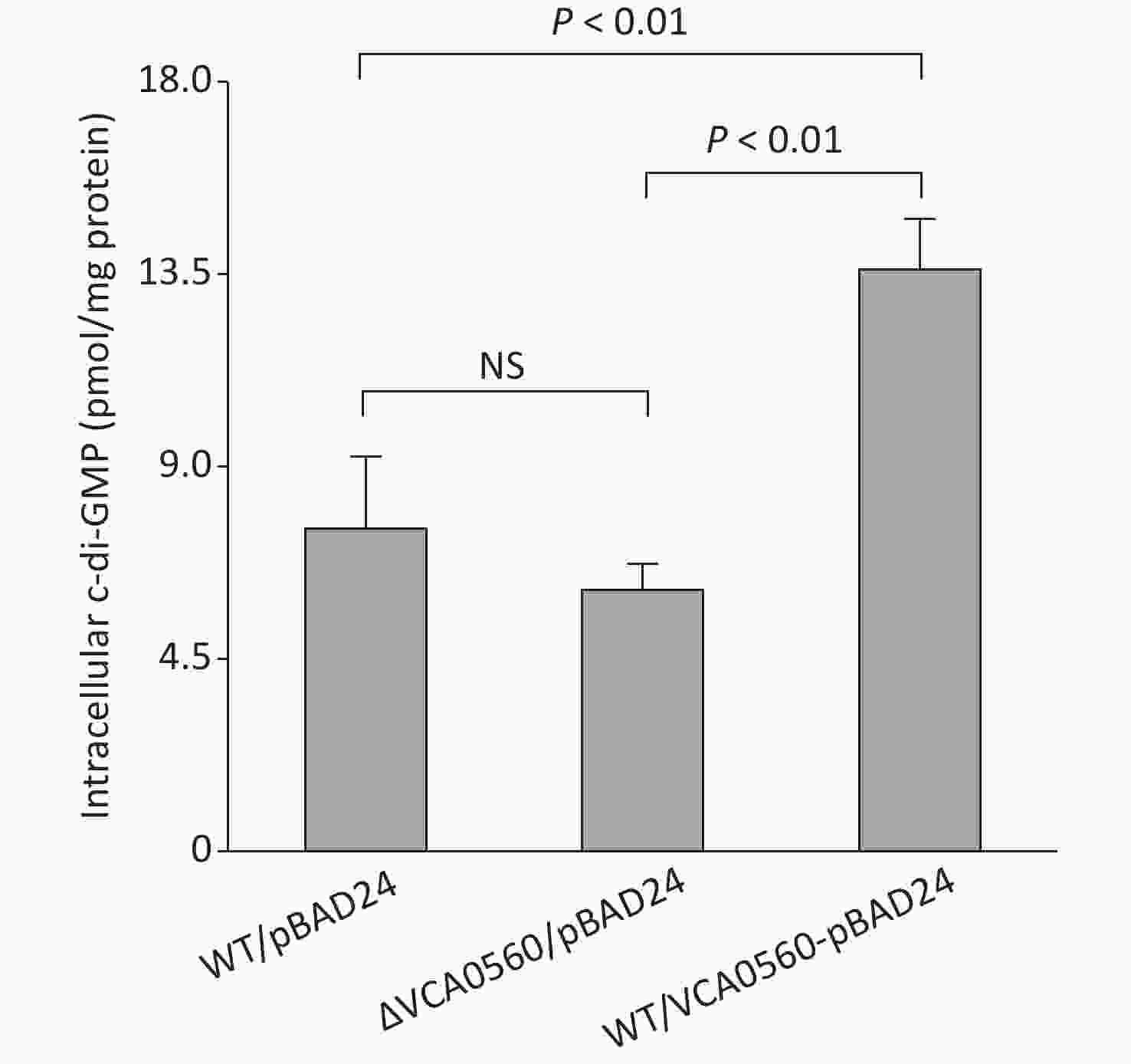
Figure 4. Intracellular c-di-GMP levels in V. cholerae strains. V. cholerae strains were cultured in LB broth supplemented with 100 mg/mL ampicillin and 0.1% arabinose at 30 °C and harvested at OD600 = 0.6. Intracellular c-di-GMP was extracted by ultrasonication, and its concentration was measured by a c-di-GMP ELISA kit (Mskbio, Beijing, China) and expressed as pmol/mg. NS represents no significant difference.
-
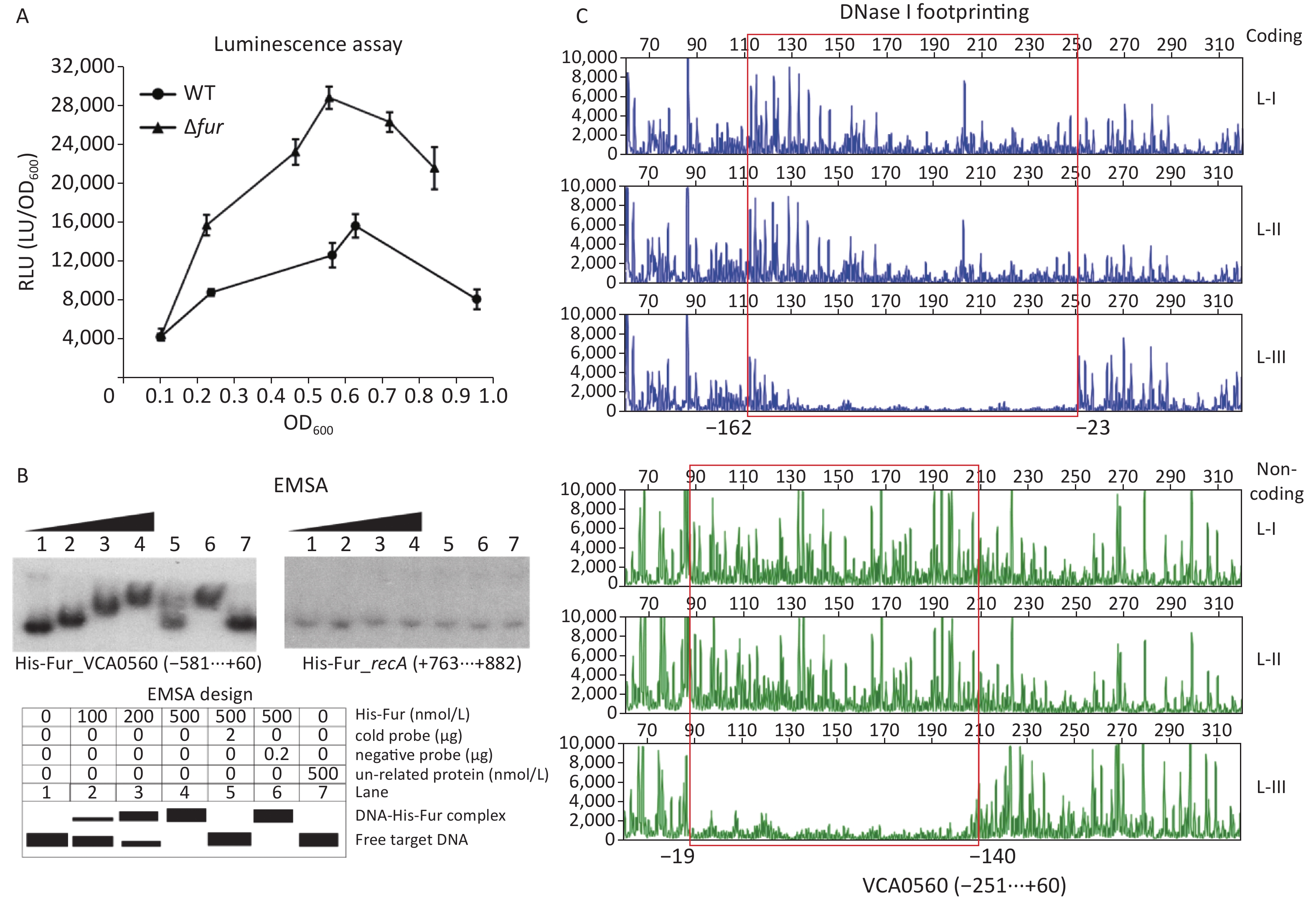
A Fur consensus-like sequence AATAAGCATAGTTTTCATC with a weighted score of 8.1 located from −126 bp to −108 bp was detected within the upstream of the VCA0560 gene[38,52], suggesting that VCA0560 transcription might be directly regulated by Fur in V. cholerae. In a previous study, Fur has been demonstrated to repress c-di-GMP production and biofilm formation by V. cholerae independent of iron[13]. Thus, the bacterium was cultured in LB broth and then harvested to investigate the Fur-dependent transcription of VCA0560. As shown in Figure 5A, the luminescence activity under the direct action of the VCA0560 promoter was higher in Δfur than in WT across the growth periods. EMSA results indicated that His-Fur dose-dependently bound to the 581 bp upstream of the VCA0560 gene but was incapable of binding to the negative control DNA from recA (Figure 5B). DNase I footprinting assay revealed that His-Fur protected a single DNA region located from −162 bp to −19 bp upstream of VCA0560 (Figure 5C). Thus, VCA0560 transcription was directly suppressed by Fur.
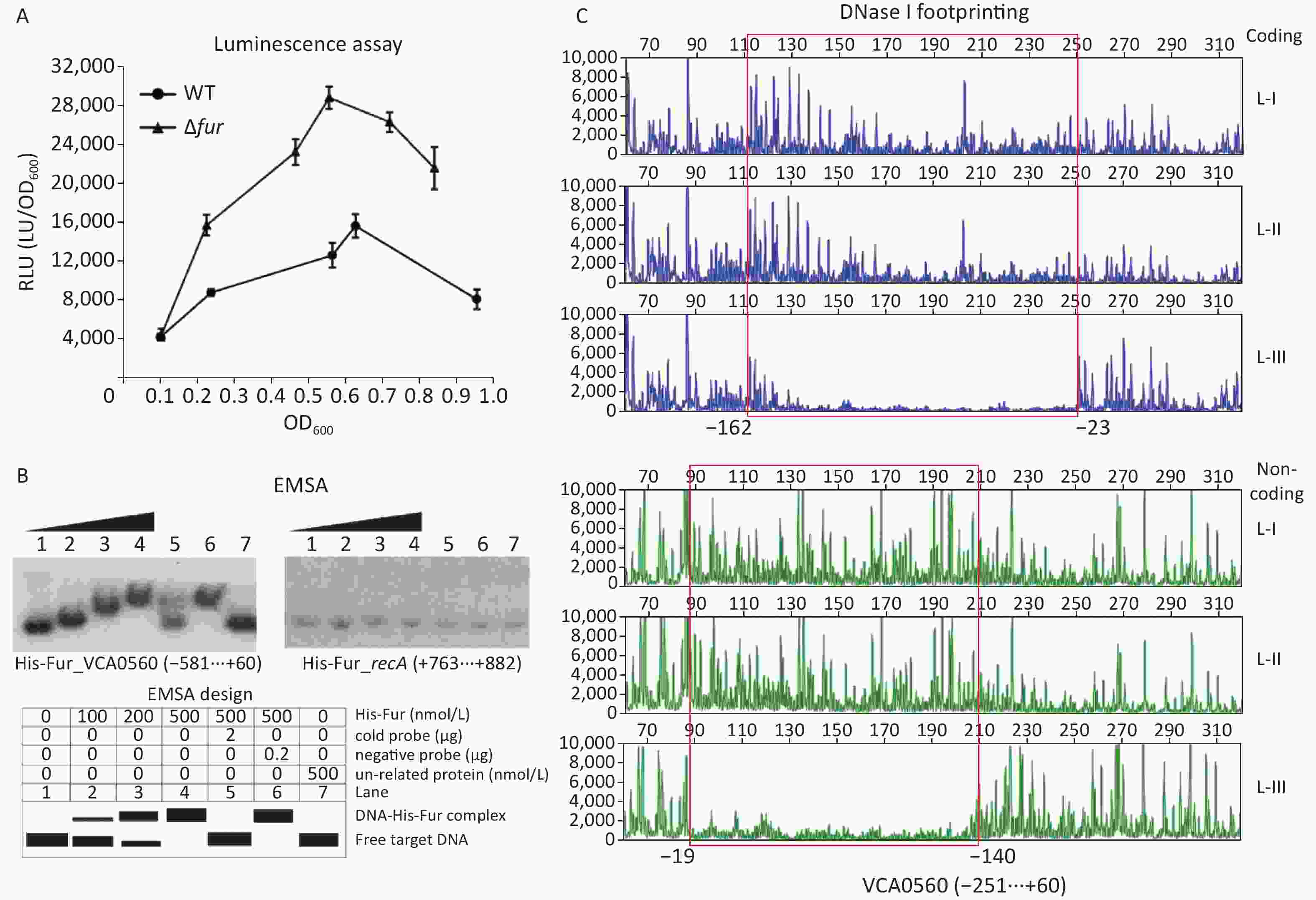
Figure 5. Fur regulated the transcription of the VCA0560 gene. The first base of the start codon of VCA0650 was defined as +1. (A) Luminescence assay. The recombinant pBBRlux plasmid containing the regulatory DNA region of VCA0560 was introduced into Δfur and WT to measure the lux activity (RLU; the light units/OD600) for each strain. (B) EMSA. The [γ-32P]-ATP-labeled DNA probe of VCA0560 was incubated with different amounts of His-Fur and then analyzed by 4% (w/v) polyacrylamide gel electrophoresis. The results were detected by autoradiography. (C) DNase I footprinting. Fluorescently-labeled DNA probe of VCA0560 was incubated with different amounts of His-Fur (L-I, II, and III containing 0, 2.95, and 8.85 pmol His-Fur, respectively) and then digested with DNase I. The protected regions are marked with boxes and positions.
-
An MQSR consensus-like sequence TAATTGTAGAATTATCATTA with a weighted score of 8.3 located from −46 bp to −27 bp was detected within the upstream of the VCA0560 gene[48], suggesting that VCA0560 transcription might be directly regulated by HapR. As shown in Figure 6A, VCA0560 expressed was enhanced significantly in ΔhapR compared with that in WT across the growth periods. EMSA results demonstrated that His-HapR dose-dependently bound to the regulatory DNA region of the VCA0560 gene but was incapable of binding to the negative control DNA fragment from recA (Figure 6B). DNase I footprinting assay results showed that His-HapR bound one DNA region located from −59 bp to −11 bp upstream of the coding region of VCA0560 (Figure 6C). Thus, VCA0560 transcription was directly inhibited by HapR.
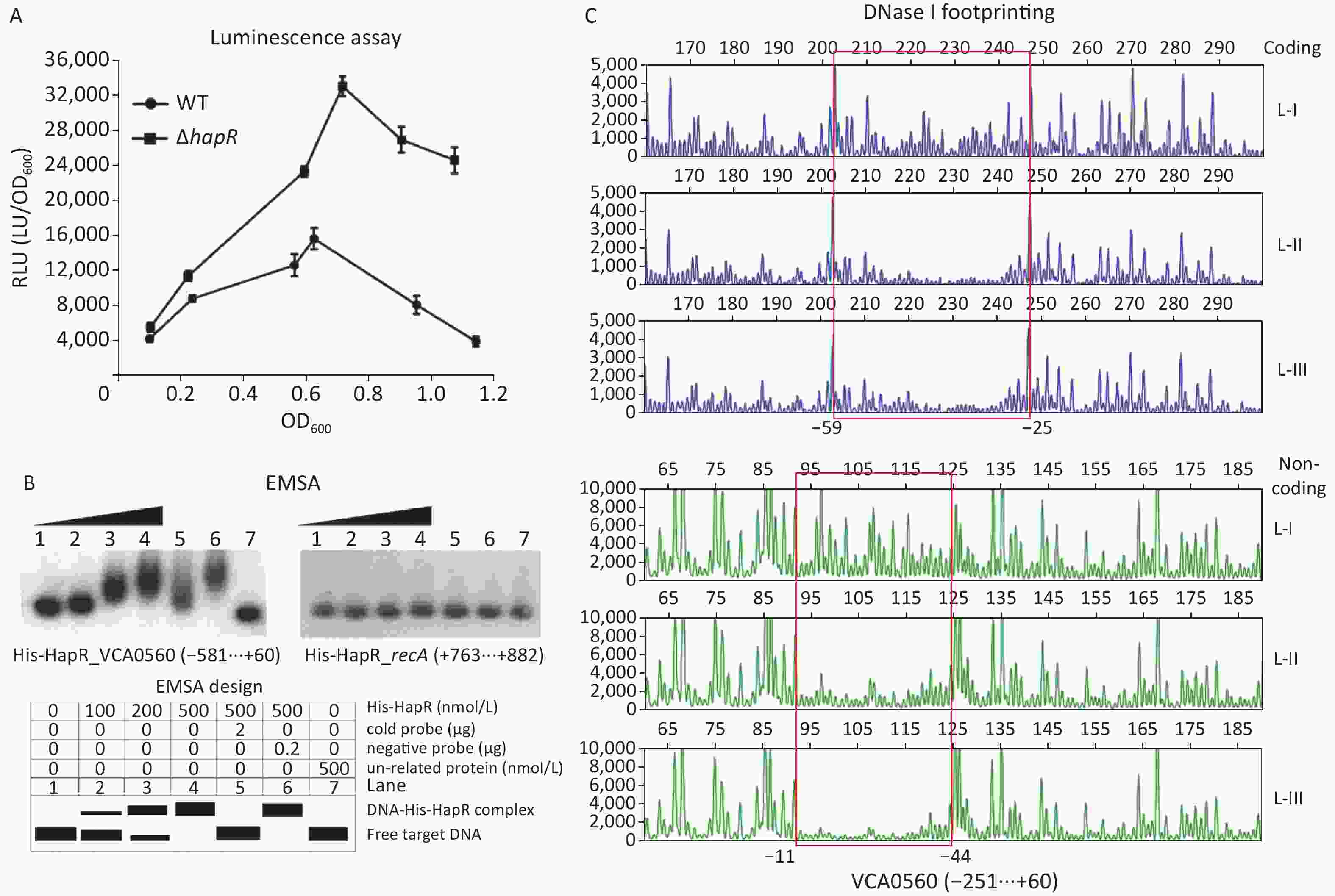
Figure 6. HapR regulated the transcription of the VCA0560 gene. The first base of the start codon of VCA0650 was defined as +1. Luminescence assay (A), EMSA (B), and DNase I footprinting (C) were performed as described in Figure 5.
-
In this work, we showed that the overexpression of the VCA0560 gene, which encodes a GGDEF-type protein in V. cholerae, induced biofilm formation and c-di-GMP production but inhibited the motility of V. cholerae. Meanwhile, the deletion of the VCA0560 gene had no influence on c-di-GMP synthesis, biofilm formation, and motility (Figures 2–4). These results suggested that the VCA0560 protein acts as an active DGC in V. cholerae, but its activity is low. A previous study also showed that VCA0560 overexpression seems to have only a weakly positive effect on the biofilm formation by V. cholerae[36]. The deletion of a DGC gene does not alter the phenotype changes because more than 30 GGDEF-containing proteins with a high level of functional redundancy can be found in V. cholerae[20]. The VCA0560 protein is perhaps not the main enzyme that modulates c-di-GMP synthesis under the growth conditions in the current work. VCA0560 expression is significantly induced in the early stage than in the late stage of cholera infection[53]. V. cholerae forms biofilms almost immediately after it adheres to the intestinal cells[2,3]. Thus, the VCA0560 protein might play some roles in the early stage of V. cholerae infection by promoting c-di-GMP synthesis and biofilm formation. Further investigations are warranted to verify this hypothesis.
A Fur consensus-like sequence and an MQSR consensus-like sequence were detected in the regulatory DNA region of the VCA0560 gene. Thus, we further investigated the transcriptional regulation of the VCA0560 gene by Fur and HapR. The data showed that VCA0560 transcription was directly repressed by HapR and Fur (Figures 5 and 6). In the presence and absence of HapR and Fur, the promoter activity of the VCA0560 gene increased with cell density until OD600 = 0.6 and then decreased over time (Figures 5A and 6A). Thus, VCA0560 might be repressed strongly by some other unknown regulators operating at low and high cell densities. In addition, primer extension[13,39] and 5ʹ-RACE[13] did not detect any transcription starts for VCA0560 (data not shown). Only one transcription start located at 124 bp upstream of the VCA0560 gene was predicted using the online tool SoftBerry (
http://linux1.softberry.com/ ) (Figure 7). The putative −10 element was well matched to the −10 consensus, but the putative −35 element possessed three bases mismatch with the consensus (Figure 7), suggesting that the promoter is relatively weak. The Fur site overlapped with the putative −10 and −35 elements and the predicted transcription start, indicating that Fur represses VCA0560 transcription possibly via its direct interference with RNA polymerase (RNAP) function. In contrast, the HapR site was located downstream of the predicted transcription start. Thus, HapR represses VCA0560 transcription, possibly by blocking RNAP elongation. The HapR site almost entirely overlapped with the Fur site (Figure 7). Whether HapR cooperates with Fur to repress VCA0560 transcription or not warrants further investigation.
Figure 7. Promoter organization of VCA0560 gene. The predicted transcription and translation start sites are marked with boxes and bent arrows. The putative −10/−35 elements are enclosed in boxes. The Fur site is marked with a solid line, and the HapR site is marked with a dotted line.
HapR acts as a negative regulator of biofilm formation, intracellular c-di-GMP synthesis, and many other gene loci in V. cholerae[15,54-56]. In addition, HapR regulates the transcription of 14 genes involved in c-di-GMP metabolism in V. cholerae, but the VCA0560 gene is not included[15]. The above data were obtained in the genetic background of luxO mutant[15]. LuxO is a central quorum regulator that controls the QS circuit and many other genes[57,58]. Therefore, the discrepancies between previous and present data may be mainly due to the bacterial genetic background. Moreover, Fur acts as a repressor of biofilm formation and c-di-GMP synthesis in V. cholerae[13], and its transcription is directly repressed by HapR[39]. The current study showed that Fur and HapR directly repressed the transcription of VCA0560 encoding a GGDEF-type protein that acts as an active DGC in V. cholerae. Thus, Fur and HapR compose a regulatory circuit to tightly regulate biofilm formation at multiple genes and levels.
In conclusion, VCA0560 overexpression significantly enhances biofilm formation and intracellular c-di-GMP levels but reduces the motility capacity of V. cholerae. Fur and HapR bind to the regulatory DNA region of the VCA0560 gene to repress its transcription. Therefore, overexpressed VCA0560 acts as an active DGC in V. cholerae, and its transcription is repressed by Fur and HapR.
doi: 10.3967/bes2023.080
Protein Containing the GGDEF Domain Affects Motility and Biofilm Formation in Vibrio cholerae and is Negatively Regulated by Fur and HapR
-
Abstract:
Objective This study aimed to investigate whether the VCA0560 gene acts as an active diguanylate cyclase (DGC) in Vibrio cholerae and how its transcription is regulated by Fur and HapR. Methods The roles of VCA0560 was investigated by utilizing various phenotypic assays, including colony morphological characterization, crystal violet staining, Cyclic di-GMP (c-di-GMP) quantification, and swimming motility assay. The regulation of the VCA0560 gene by Fur and HapR was analyzed by luminescence assay, electrophoretic mobility shift assay, and DNase I footprinting. Results VCA0560 gene mutation did not affect biofilm formation, motility, and c-di-GMP synthesis in V. cholerae, and its overexpression remarkably enhanced biofilm formation and intracellular c-di-GMP level but reduced motility capacity. The transcription of the VCA0560 gene was directly repressed by Fur and the master quorum sensing regulator HapR. Conclusion Overexpressed VCA0560 functions as an active DGC in V. cholerae, and its transcription is repressed by Fur and HapR. -
Key words:
- Vibrio cholera /
- Cyclic di-GMP /
- VCA0560 /
- Fur /
- HapR
-
Figure 1. Growth curves of V. cholerae strains. The overnight growth cultures of WT/pBAD24, ΔVCA0560/pBAD24, and WT/pBAD24-VCA0560 were diluted at 1:100 into 20 mL of LB broth supplemented with 100 mg/mL ampicillin and 0.1% arabinose, mixed well, and then divided into a microwell plate with 300 μL in each well. V. cholerae strains were grown at 30 °C with shaking at 200 rpm, and the OD600 values were monitored at 1 h intervals. The experiment was conducted twice, with five replicates per trial for each strain.
Figure 2. VCA0560 overexpression induced biofilm formation by V. cholerae. (A) Colony morphological characterization. In brief, 2 μL of overnight bacterial culture was spotted on the LB agar supplemented with 100 mg/mL ampicillin and 0.1% arabinose and then statically incubated at 30 °C for 48 h. (B) Crystal violet (CV) staining. Bacterial cells were inoculated into 2 mL of fresh LB broth supplemented with 100 mg/mL ampicillin and 0.1% arabinose in glass tubes and allowed to grow at 30 °C with shaking at 100 rpm for 48 h. The surface-attached cells were stained with 0.1% CV. The bound CV was dissolved with dimethylsulfoxide (DMSO), and the OD570 values were measured as the index of CV staining. NS represents no significant difference.
Figure 3. Regulation of swimming motility by VCA0560 protein in V. cholerae. In brief, 2 μL of bacterial cells were inoculated into semisolid swim plates (1% tryptone, 0.5% yeast extract, 1% NaCl, and 0.35% bacterial agar) supplemented with 100 mg/mL ampicillin and 0.1% arabinose. The diameter of the swimming areas was measured after incubation at 30 °C for 10 h.
Figure 4. Intracellular c-di-GMP levels in V. cholerae strains. V. cholerae strains were cultured in LB broth supplemented with 100 mg/mL ampicillin and 0.1% arabinose at 30 °C and harvested at OD600 = 0.6. Intracellular c-di-GMP was extracted by ultrasonication, and its concentration was measured by a c-di-GMP ELISA kit (Mskbio, Beijing, China) and expressed as pmol/mg. NS represents no significant difference.
Figure 5. Fur regulated the transcription of the VCA0560 gene. The first base of the start codon of VCA0650 was defined as +1. (A) Luminescence assay. The recombinant pBBRlux plasmid containing the regulatory DNA region of VCA0560 was introduced into Δfur and WT to measure the lux activity (RLU; the light units/OD600) for each strain. (B) EMSA. The [γ-32P]-ATP-labeled DNA probe of VCA0560 was incubated with different amounts of His-Fur and then analyzed by 4% (w/v) polyacrylamide gel electrophoresis. The results were detected by autoradiography. (C) DNase I footprinting. Fluorescently-labeled DNA probe of VCA0560 was incubated with different amounts of His-Fur (L-I, II, and III containing 0, 2.95, and 8.85 pmol His-Fur, respectively) and then digested with DNase I. The protected regions are marked with boxes and positions.
Figure 6. HapR regulated the transcription of the VCA0560 gene. The first base of the start codon of VCA0650 was defined as +1. Luminescence assay (A), EMSA (B), and DNase I footprinting (C) were performed as described in Figure 5.
Figure 7. Promoter organization of VCA0560 gene. The predicted transcription and translation start sites are marked with boxes and bent arrows. The putative −10/−35 elements are enclosed in boxes. The Fur site is marked with a solid line, and the HapR site is marked with a dotted line.
Table 1. Oligonucleotide primers used in this study
Target Primers (forward/reverse, 5'–3’) Construction of mutant VCA0560 GCGGGATCCGAACAGCGATCAGAGCATG/CCTCCTTTGCCAATTTTGAATTCCTGAGGTTCTACATTG CAATGTAGAACCTCAGGAATTCAAAATTGGCAAAGGAGG/GGACTAGTGCGAGTTGATGTGGGTAG GCGGGATCCGAACAGCGATCAGAGCATG/GGACTAGTGCGAGTTGATGTGGGTAG Construction of overexpressed plasmid VCA0560 GATTCTAGAATGTCTTCTAGCTTTGTTAC/GCGAAGCTTTTAGCTAGCGACTTTGACAC Luminescence assay VCA0560 GCGGAGCTCGAACAGCGATCAGAGCAT/GCGGGATCCCAGCAGCAATAGAGGAAAA EMSA VCA0560 GAACAGCGATCAGAGCAT/CAGCAGCAATAGAGGAAAA recA AAGATTGGTGTGATGTTTGGTA/CACTTCTTCGCCTTCTTTGA DNase I footprinting VCA0560 GTAAAACGACGGCCAGTCAAGCTGCAACGGCAAGTAG/CAGGAAACAGCTATGACCAGCAGCAATAG
AGGAAAAM13 FAM-GTAAAACGACGGCCAGT/CAGGAAACAGCTATGAC-HEX  下载: 导出CSV
下载: 导出CSV
-
[1] Clemens JD, Nair GB, Ahmed T, et al. Cholera. Lancet, 2017; 390, 1539−49. doi: 10.1016/S0140-6736(17)30559-7 [2] Tamayo R, Patimalla B, Camilli A. Growth in a biofilm induces a hyperinfectious phenotype in Vibrio cholerae. Infect Immun, 2010; 78, 3560−9. doi: 10.1128/IAI.00048-10 [3] Sengupta C, Mukherjee O, Chowdhury R. Adherence to intestinal cells promotes biofilm formation in Vibrio cholerae. J Infect Dis, 2016; 214, 1571−8. doi: 10.1093/infdis/jiw435 [4] Pruzzo C, Vezzulli L, Colwell RR. Global impact of Vibrio cholerae interactions with chitin. Environ Microbiol, 2008; 10, 1400−10. doi: 10.1111/j.1462-2920.2007.01559.x [5] Yildiz FH, Visick KL. Vibrio biofilms: so much the same yet so different. Trends Microbiol, 2009; 17, 109−18. doi: 10.1016/j.tim.2008.12.004 [6] Zamorano-Sánchez D, Fong JCN, Kilic S, et al. Identification and characterization of VpsR and VpsT binding sites in Vibrio cholerae. J Bacteriol, 2015; 197, 1221−35. doi: 10.1128/JB.02439-14 [7] Beyhan S, Bilecen K, Salama SR, et al. Regulation of rugosity and biofilm formation in Vibrio cholerae: comparison of VpsT and VpsR regulons and epistasis analysis of vpsT, vpsR, and hapR. J Bacteriol, 2007; 189, 388−402. doi: 10.1128/JB.00981-06 [8] Teschler JK, Cheng AT, Yildiz FH. The two-component signal transduction system VxrAB positively regulates Vibrio cholerae biofilm formation. J Bacteriol, 2017; 199, e00139−17. [9] Ayala JC, Wang HX, Silva AJ, et al. Repression by H-NS of genes required for the biosynthesis of the Vibrio cholerae biofilm matrix is modulated by the second messenger cyclic diguanylic acid. Mol Microbiol, 2015; 97, 630−45. doi: 10.1111/mmi.13058 [10] Fong JCN, Yildiz FH. Interplay between cyclic AMP-cyclic AMP receptor protein and cyclic di-GMP signaling in Vibrio cholerae biofilm formation. J Bacteriol, 2008; 190, 6646−59. doi: 10.1128/JB.00466-08 [11] Bilecen K, Yildiz FH. Identification of a calcium-controlled negative regulatory system affecting Vibrio cholerae biofilm formation. Environ Microbiol, 2009; 11, 2015−29. doi: 10.1111/j.1462-2920.2009.01923.x [12] Sultan SZ, Silva AJ, Benitez JA. The PhoB regulatory system modulates biofilm formation and stress response in El Tor biotype Vibrio cholerae. FEMS Microbiol Lett, 2010; 302, 22−31. doi: 10.1111/j.1574-6968.2009.01837.x [13] Gao H, Ma LZ, Qin Q, et al. Fur represses Vibrio cholerae biofilm formation via direct regulation of vieSAB, cdgD, vpsU, and vpsA-K transcription. Front Microbiol, 2020; 11, 587159. doi: 10.3389/fmicb.2020.587159 [14] Zhu J, Mekalanos JJ. Quorum sensing-dependent biofilms enhance colonization in Vibrio cholerae. Dev Cell, 2003; 5, 647−56. doi: 10.1016/S1534-5807(03)00295-8 [15] Waters CM, Lu WY, Rabinowitz JD, et al. Quorum sensing controls biofilm formation in Vibrio cholerae through modulation of cyclic di-GMP levels and repression of vpsT. J Bacteriol, 2008; 190, 2527−36. doi: 10.1128/JB.01756-07 [16] Mills E, Pultz IS, Kulasekara HD, et al. The bacterial second messenger c-di-GMP: mechanisms of signalling. Cell Microbiol, 2011; 13, 1122−9. doi: 10.1111/j.1462-5822.2011.01619.x [17] Fernandez NL, Srivastava D, Ngouajio AL, et al. Cyclic di-GMP positively regulates DNA repair in Vibrio cholerae. J Bacteriol, 2018; 200, e00005−18. [18] Fernandez NL, Hsueh BY, Nhu NTQ, et al. Vibrio cholerae adapts to sessile and motile lifestyles by cyclic di-GMP regulation of cell shape. Proc Natl Acad Sci USA, 2020; 117, 29046−54. doi: 10.1073/pnas.2010199117 [19] Beyhan S, Tischler AD, Camilli A, et al. Transcriptome and phenotypic responses of Vibrio cholerae to increased cyclic di-GMP level. J Bacteriol, 2006; 188, 3600−13. doi: 10.1128/JB.188.10.3600-3613.2006 [20] Seshasayee ASN, Fraser GM, Luscombe NM. Comparative genomics of cyclic-di-GMP signalling in bacteria: post-translational regulation and catalytic activity. Nucl Acids Res, 2010; 38, 5970−81. doi: 10.1093/nar/gkq382 [21] Kovacikova G, Lin W, Skorupski K. Dual regulation of genes involved in acetoin biosynthesis and motility/biofilm formation by the virulence activator AphA and the acetate-responsive LysR-type regulator AlsR in Vibrio cholerae. Mol Microbiol, 2005; 57, 420−33. doi: 10.1111/j.1365-2958.2005.04700.x [22] Lim B, Beyhan S, Meir J, et al. Cyclic-diGMP signal transduction systems in Vibrio cholerae: modulation of rugosity and biofilm formation. Mol Microbiol, 2006; 60, 331−48. doi: 10.1111/j.1365-2958.2006.05106.x [23] Lim B, Beyhan S, Yildiz FH. Regulation of Vibrio polysaccharide synthesis and virulence factor production by CdgC, a GGDEF-EAL domain protein, in Vibrio cholerae. J Bacteriol, 2007; 189, 717−29. doi: 10.1128/JB.00834-06 [24] Syed KA, Beyhan S, Correa N, et al. The Vibrio cholerae flagellar regulatory hierarchy controls expression of virulence factors. J Bacteriol, 2009; 191, 6555−70. doi: 10.1128/JB.00949-09 [25] Beyhan S, Odell LS, Yildiz FH. Identification and characterization of cyclic diguanylate signaling systems controlling rugosity in Vibrio cholerae. J Bacteriol, 2008; 190, 7392−405. doi: 10.1128/JB.00564-08 [26] Liu XX, Beyhan S, Lim B, et al. Identification and characterization of a phosphodiesterase that inversely regulates motility and biofilm formation in Vibrio cholerae. J Bacteriol, 2010; 192, 4541−52. doi: 10.1128/JB.00209-10 [27] Tamayo R, Schild S, Pratt JT, et al. Role of cyclic Di-GMP during el tor biotype Vibrio cholerae infection: characterization of the in vivo-induced cyclic Di-GMP phosphodiesterase CdpA. Infect Immun, 2008; 76, 1617−27. doi: 10.1128/IAI.01337-07 [28] Bordeleau E, Brouillette E, Robichaud N, et al. Beyond antibiotic resistance: integrating conjugative elements of the SXT/R391 family that encode novel diguanylate cyclases participate to c-di-GMP signalling in Vibrio cholerae. Environ Microbiol, 2010; 12, 510−23. doi: 10.1111/j.1462-2920.2009.02094.x [29] Bomchil N, Watnick P, Kolter R. Identification and characterization of a Vibrio cholerae gene, mbaA, involved in maintenance of biofilm architecture. J Bacteriol, 2003; 185, 1384−90. doi: 10.1128/JB.185.4.1384-1390.2003 [30] Chouhan OP, Bandekar D, Hazra M, et al. Effect of site-directed mutagenesis at the GGEEF domain of the biofilm forming GGEEF protein from Vibrio cholerae. AMB Express, 2016; 6, 2. doi: 10.1186/s13568-015-0168-6 [31] Dey AK, Bhagat A, Chowdhury R. Host cell contact induces expression of virulence factors and VieA, a cyclic di-GMP phosphodiesterase, in Vibrio cholerae. J Bacteriol, 2013; 195, 2004−10. doi: 10.1128/JB.02127-12 [32] Tamayo R, Tischler AD, Camilli A. The EAL domain protein VieA is a cyclic diguanylate phosphodiesterase. J Biol Chem, 2005; 280, 33324−30. doi: 10.1074/jbc.M506500200 [33] Beyhan S, Yildiz FH. Smooth to rugose phase variation in Vibrio cholerae can be mediated by a single nucleotide change that targets c-di-GMP signalling pathway. Mol Microbiol, 2007; 63, 995−1007. doi: 10.1111/j.1365-2958.2006.05568.x [34] Roelofs KG, Jones CJ, Helman SR, et al. Systematic identification of cyclic-di-GMP binding proteins in Vibrio cholerae reveals a novel class of cyclic-di-GMP-binding ATPases associated with type II secretion systems. PLoS Pathog, 2015; 11, e1005232. doi: 10.1371/journal.ppat.1005232 [35] Hunter JL, Severin GB, Koestler BJ, et al. The Vibrio cholerae diguanylate cyclase VCA0965 has an AGDEF active site and synthesizes cyclic di-GMP. BMC Microbiol, 2014; 14, 22. doi: 10.1186/1471-2180-14-22 [36] Massie JP, Reynolds EL, Koestler BJ, et al. Quantification of high-specificity cyclic diguanylate signaling. Proc Natl Acad Sci USA, 2012; 109, 12746−51. doi: 10.1073/pnas.1115663109 [37] Ng WL, Bassler BL. Bacterial quorum-sensing network architectures. Annu Rev Genet, 2009; 43, 197−222. doi: 10.1146/annurev-genet-102108-134304 [38] Davies BW, Bogard RW, Mekalanos JJ. Mapping the regulon of Vibrio cholerae ferric uptake regulator expands its known network of gene regulation. Proc Natl Acad Sci USA, 2011; 108, 12467−72. doi: 10.1073/pnas.1107894108 [39] Gao H, Xu JL, Lu X, et al. Expression of hemolysin is regulated under the collective actions of HapR, Fur, and HlyU in Vibrio cholerae El tor serogroup O1. Front Microbiol, 2018; 9, 1310. doi: 10.3389/fmicb.2018.01310 [40] Wu R, Zhao M, Li J, et al. Direct regulation of the natural competence regulator gene tfoX by cyclic AMP (cAMP) and cAMP receptor protein (CRP) in Vibrios. Sci Rep, 2015; 5, 14921. doi: 10.1038/srep14921 [41] Sun FJ, Zhang YQ, Qiu YF, et al. H-NS is a repressor of major virulence gene loci in Vibrio parahaemolyticus. Front Microbiol, 2014; 5, 675. [42] Fang N, Gao H, Wang L, et al. Optimized methods for biofilm analysis in Yersinia pestis. Biomed Environ Sci, 2013; 26, 408−11. [43] Xue XF, Zhnag MM, Sun JF, et al. H-NS represses biofilm formation and c-di-GMP synthesis in Vibrio parahaemolyticus. Biomed Environ Sci, 2022; 35, 821−9. [44] Wang L, Ling Y, Jiang HW, et al. AphA is required for biofilm formation, motility, and virulence in pandemic Vibrio parahaemolyticus. Int J Food Microbiol, 2013; 160, 245−51. doi: 10.1016/j.ijfoodmicro.2012.11.004 [45] Xu X, Stern AM, Liu Z, et al. Virulence regulator AphB enhances toxR transcription in Vibrio cholerae. BMC Microbiol, 2010; 10, 3. doi: 10.1186/1471-2180-10-3 [46] Kleber-Janke T, Becker WM. Use of modified BL21(DE3) Escherichia coli cells for high-level expression of recombinant peanut allergens affected by poor codon usage. Protein Expr Purif, 2000; 19, 419−24. doi: 10.1006/prep.2000.1265 [47] Gao H, Zhou DS, Li YL, et al. The iron-responsive Fur regulon in Yersinia pestis. J Bacteriol, 2008; 190, 3063−75. doi: 10.1128/JB.01910-07 [48] Zhang YQ, Qiu YF, Tan YF, et al. Transcriptional regulation of opaR, qrr2-4 and aphA by the master quorum-sensing regulator OpaR in Vibrio parahaemolyticus. PLoS One, 2012; 7, e34622. doi: 10.1371/journal.pone.0034622 [49] Zhang YQ, Zhang Y, Gao H, et al. Vibrio parahaemolyticus CalR down regulates the thermostable direct hemolysin (TDH) gene transcription and thereby inhibits hemolytic activity. Gene, 2017; 613, 39−44. doi: 10.1016/j.gene.2017.03.001 [50] Osei-Adjei G, Gao H, Zhang Y, et al. Regulatory actions of ToxR and CalR on their own genes and type III secretion system 1 in Vibrio parahaemolyticus. Oncotarget, 2017; 8, 65809−22. doi: 10.18632/oncotarget.19498 [51] Gao H, Zhang LY, Osei-Adjei G, et al. Transcriptional regulation of cpsQ-mfpABC and mfpABC by CalR in Vibrio parahaemolyticus. Microbiologyopen, 2017; 6, e00470. doi: 10.1002/mbo3.470 [52] Mey AR, Wyckoff EE, Kanukurthy V, et al. Iron and fur regulation in Vibrio cholerae and the role of fur in virulence. Infect Immun, 2005; 73, 8167−78. doi: 10.1128/IAI.73.12.8167-8178.2005 [53] Larocque RC, Harris JB, Dziejman M, et al. Transcriptional profiling of Vibrio cholerae recovered directly from patient specimens during early and late stages of human infection. Infect Immun, 2005; 73, 4488−93. doi: 10.1128/IAI.73.8.4488-4493.2005 [54] Hammer BK, Bassler BL. Quorum sensing controls biofilm formation in Vibrio cholerae. Mol Microbiol, 2003; 50, 101−4. doi: 10.1046/j.1365-2958.2003.03688.x [55] Ball AS, Chaparian RR, van Kessel JC. Quorum sensing gene regulation by LuxR/HapR master regulators in Vibrios. J Bacteriol, 2017; 199, e00105−17. [56] Zhang YQ, Ma LZ, Gao Y, et al. Master quorum sensing regulator HapR acts as a repressor of the mannitol phosphotransferase system operon in Vibrio cholerae. Biomed Environ Sci, 2022; 35, 69−72. [57] Milton DL. Quorum sensing in vibrios: complexity for diversification. Int J Med Microbiol, 2006; 296, 61−71. doi: 10.1016/j.ijmm.2006.01.044 [58] Zhu J, Miller MB, Vance RE, et al. Quorum-sensing regulators control virulence gene expression in Vibrio cholerae. Proc Natl Acad Sci USA, 2002; 99, 3129−34. doi: 10.1073/pnas.052694299 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1841
- HTML全文浏览量: 796
- PDF下载量: 86
- 被引次数: 0

 Quick Links
Quick Links