

 下载:
下载:




-
With an aging population, Intracranial hemorrhage (ICH) has become the second most common subtype of stroke, surpassed only by ischemic stroke, and its fatality rate is more than twice that of ischemic stroke[1,2]. Following ICH, hematoma expansion and edema are accompanied by intricate processes that involve inflammation, vascular dynamics, and pathological metabolic clearance. In particular, vasogenic edema arises from the disruption of tight endothelial junctions on the microvascular side of the blood-brain barrier (BBB), leading to increased accumulation of fluid due to extravasation and posing a serious threat to the health of affected individuals[3,4]. Therefore, preserving the ability of the BBB to remove hematoma metabolites is essential to maintain brain health and function, and to ensure an optimal cerebral environment.
In recent years, researchers have discovered that the optimal functioning of a healthy brain significantly depends on the dynamic interactions between neurons and non-neuronal cells. Consequently, the concept of a neurovascular unit (NVU) has been introduced as a focal point in nervous system research[5]. The NVU comprises neurons, astrocytes, microglia, the blood-brain barrier, and the extracellular matrix, which bridges these components. Together, they maintain the internal environmental integrity of brain tissue and dynamically regulate local cerebral blood flow and metabolic clearance through neurovascular time[6,7].
In the NVU, the primary function of neurons is to initiate local vascular responses. Neurons regulate cerebral blood flow (CBF) by producing signaling molecules, such as glutamate and adenosine, which act directly or indirectly on local blood vessels, inducing their dilation or contraction of the blood vessels[8]. Astrocell foot processes are anchored at the vascular interface and serve as the gatekeepers of the BBB, controlling and clearing cerebrospinal fluid from the perivascular space[9,10]. Aquaporin 4 (AQP4), expressed in astrocyte foot processes, functions as a water gate, facilitating the entry of cerebrospinal fluid into the brain parenchyma for metabolite exchange[11,12]. Previous research has also shown the involvement of scavenger receptors such as low-density lipoprotein receptor-related protein 1 (LRP1) and CD163 on the surfaces of microglia and astrocytes in the clearance of hematomas after ICH[13-15].
Considering the vital role of the BBB in preserving central nervous system homeostasis, and the intricate connections between circadian rhythms and functional NVU, disruptions in circadian rhythms have been suggested to contribute to the pathology of various chronic brain diseases[16]. In sleep disorders, there is a reduction in the flow of cerebrospinal fluid within the perivascular space. Sleep disorders have been speculated to result in depolarization of perivascular AQP4 and increased resistance to fluid outflow between brain tissues. This decreases the efficiency of cerebral lymphatic drainage system clearance, which could impact circadian regulation of the lymphatic system and clearance function throughout the BBB via AQP4[17]. Moreover, short-wavelength blue light causes biological and psychological internal rhythms to remain highly synchronized with circadian rhythms. Exposure to blue light can inhibit the secretion of melatonin, and exposure to blue light during the day is important for maintaining the health, alertness, and cognitive ability of the body. However, direct exposure to low-intensity blue light at night may have a serious impact on sleep quality and circadian rhythm[18,19].
In this study, we established an in vitro NVU model by co-culturing hemoglobin and simulated circadian rhythms using short-wavelength blue-light exposure. We aimed to investigate the impact of circadian rhythms on BBB function, neuronal damage, and BBB clearance, along with potential underlying mechanisms.
-
Sprague-Dawley (SD) rats were obtained from the Experimental Animal Center of Shanxi Medical University (Shanxi, China). All procedures involving the use of SD rats were approved by the Ethics Committee of the Animal Transformation Center of Shanxi Medical University (No. DW2022029).
-
Primary BMECs were isolated from 3-day-old SD rats[20,21]. Briefly, large vessels, meninges, and white matter were removed, and the forebrain tissue was minced into small pieces of approximately 1.0 mm3. After centrifugation at 150 ×g for 3 min, the precipitate layer was treated with trypsin (2.5 mg/mL, Amresco, USA) and digested at 37 °C for 1.5 h. Dulbecco’s modified eagle medium/nutrient mixture F-12 (DMEM/F12) (Gibco, Invitrogen Corporation, USA) with 10% fetal bovine serum (FBS, Hyclone, Thermo Scientific, USA) was added to stop digestion, followed by centrifugation at 150 ×g for 5 min. The precipitate was resuspended in an equal volume of 25% (w/v) bovine serum albumin (BSA, Sigma, USA) and centrifuged at 1,600 ×g for 5 min. The obtained microvessels were digested with type-2 collagenase (1.0 mg/mL, Sigma, USA) for 1 h and cultured in DMEM/F12 supplemented with 20% FBS and 1% penicillin/streptomycin. BMECs were collected and seeded in 25 cm2 flasks pre-coated with 2% gelatin. Primary BMECs were characterized for the von Willebrand factor (vWf) using fluorescence imaging.
-
Neurons were isolated from 1-day-old rats following established protocols[20,21]. Briefly, after removal of the meninges and white matter, the purified cerebral cortices were digested with 0.125% trypsin-ethylenediaminetetraacetic acid (Sigma, USA) and resuspended in neurobasal-A medium (Gibco, Grand Island, USA) containing 10% FBS, 2% B27 supplement (Invitrogen, Carlsbad, USA), and 1% penicillin/streptomycin. Cells were seeded at a density of 2 × 106 cells/mL in dishes coated with 0.01% poly-L-lysine (Sigma, USA) and cultured at 37 °C in a 5% CO2 incubator (Thermo Fisher, Fremont, USA). On the third day, the medium was replaced with fresh medium supplemented with Ara-C (5.0 mg/mL, Sigma) for 24 h to inhibit non-neuronal cell growth. The culture medium was replaced every 48 h. Primary neurons were characterized by immunofluorescence labeling for tubulin.
-
Primary astrocytes were obtained from 2-day-old rats following established procedures[20]. Briefly, isolated cells were incubated in flasks pre-coated with poly-L-lysine in DMEM/F12 containing 20% FBS and 1% penicillin-streptomycin. The fibroblasts were removed after differential attachment. On day 5, the cultures were shaken overnight to eliminate microglia[21]. Primary astrocytes were characterized by immunofluorescence labeling for glial fibrillary acidic protein (GFAP).
-
Rat microvascular microglial cells (Procell Life Science and Technology Co., Ltd., CP-R110) were obtained and cultured according to the manufacturer’s instructions. Primary microglial cells were characterized by immunofluorescence labeling with CD11b.
-
Cells were seeded in 6-well plates containing cell carriers. After adherence, cell-covered carriers were removed, rinsed with phosphate-buffered solution (PBS), fixed with 4% paraformaldehyde (PFA) for 15 min, and permeabilized with 1% X-Triton for 20 min. Subsequently, the cells were blocked with 10% goat serum (Sigma) for 1 h. The cells were then incubated overnight with rabbit anti-von Willebrand factor (1:100; ab6994, Abcam), rabbit anti-Tubulin (1:100; ab18207, Abcam), mouse anti-GFAP (1:100; ab10062, Abcam), and mouse anti-CD11b (1:100; ab1211, Abcam) antibodies at 4 °C.
The following day, cells were incubated with fluorescently-labeled secondary antibodies (FITC-conjugated goat anti-rabbit IgG, 1:200, ab6717, Abcam; Alexa Fluor 647-conjugated goat anti-mouse IgG, 1:200, ab150115, Abcam) for 1 h at 37 °C. The imaging was performed using a fluorescence microscope (Fluoview 1000, Olympus, Tokyo, Japan). The number of cell bodies and nuclei were quantified using ImageJ software. The purity was calculated as the ratio of the number of cell bodies to the number of nuclei.
-
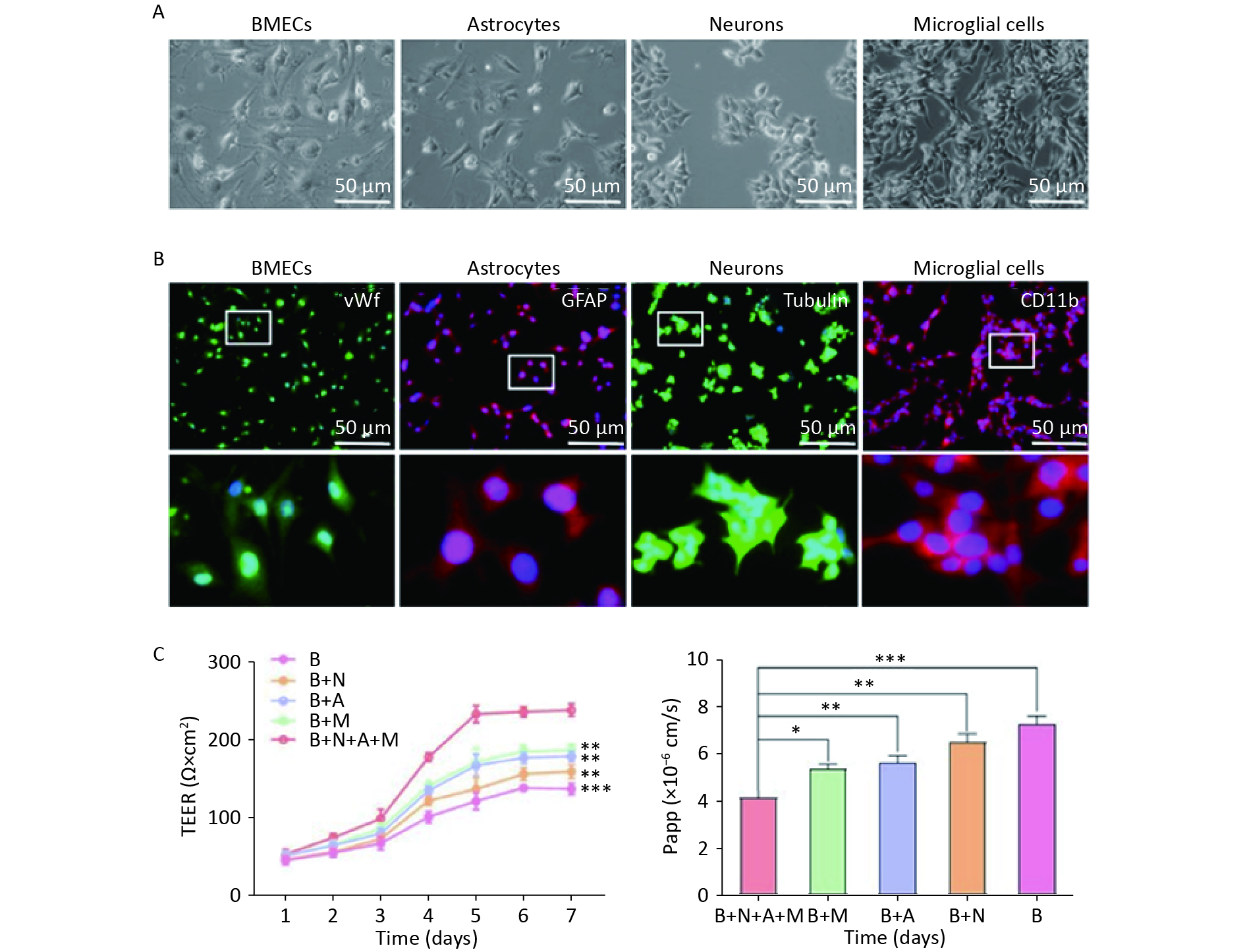
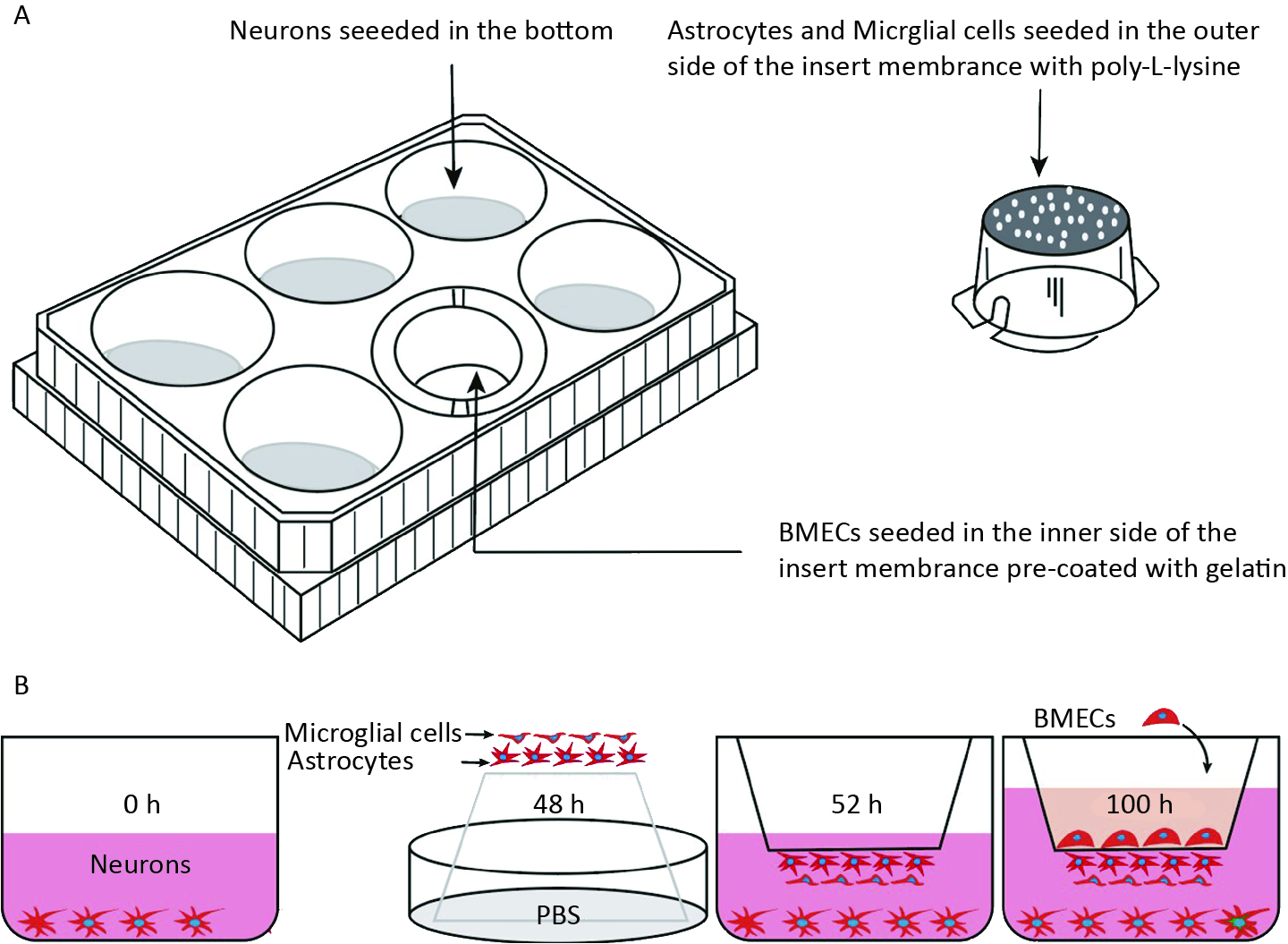
As illustrated in Figure 1A, a four-cell co-culture system was established based on a previous study with modifications. Neurons were seeded at the bottom of transwell plates (0.4 μm, Corning, New York, NY, USA) and maintained in Neurobasal-A medium containing 2% B27 and 1% penicillin/streptomycin for 48 h. Astrocytes and microglial cells (2 × 105 cells/cm2) were seeded beneath the insert membrane in a Petri dish for 48 h. After 4 h, the transwell inserts were inverted and transferred to the plates. After an additional 48 h, the BMECs (5 × 105 cells/cm2) were seeded on the inner side of the insert membrane coated with gelatin (30.0 mg/mL). For the control groups, BMECs, neurons, astrocytes, and microglial cells were cultured individually in transwell chambers. BMECs were also cultured with neurons alone, astrocytes alone, microglial cells alone, or without the other three primary cell types, designated as the B + N, B + A, B + M, and B groups, respectively. The day the BMECs were seeded was defined as day zero in vitro (day 0). All cells were cultured for 3−5 days with a daily change in the medium before analysis. The procedure used to establish the model is shown in Figure 1B.
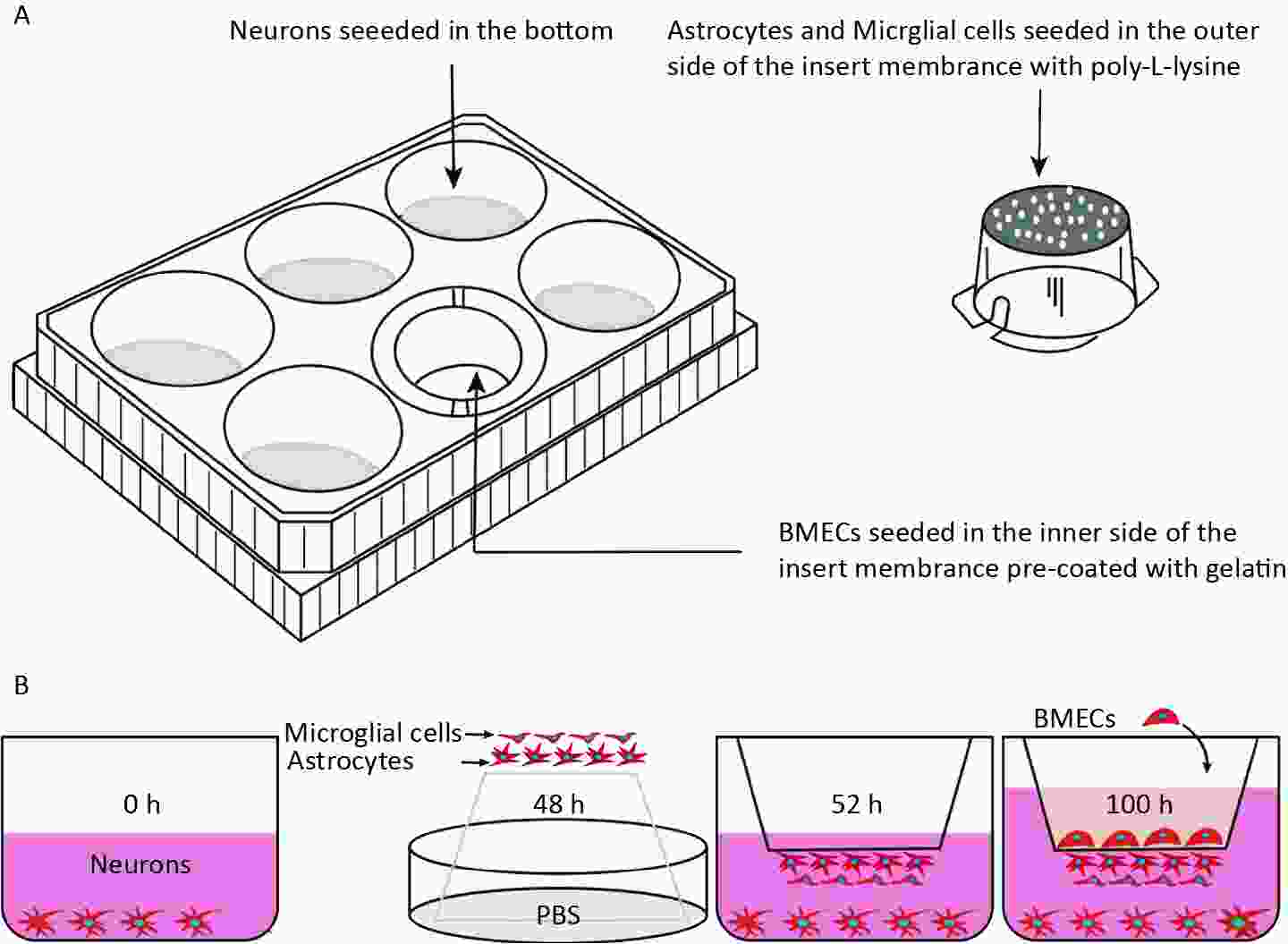
Figure 1. Establishment of an in vitro NVU model. (A) Schematic drawing of the four-cell co-culture system. (B) The procedure for establishing an in vitro four-cell co-culture model. NUV, neurovascular unit; BMECs, brain microvascular endothelial cells.
-
The TEER values for different cell co-cultures were assessed using an epithelial-volt-ohm resistance meter (ERS-2, Millipore, USA) according to the manufacturer’s protocol. The background TEER value was measured in the same well under identical conditions but without seeded cells. The final result was calculated by subtracting the corresponding background TEER value from the TEER value of different cells and multiplying it by the area of the insert membrane. The values are presented as Ω × cm2.
-
The permeability of the BMECs was assessed using a 4-h leak experiment. After 5 days of co-culture, the insert, which serves as the donor pool, was filled with medium, while the medium level in the 12-well plates, which acts as the receptor pool, was maintained 0.5 cm lower than an insert. Inserts without cell cultures were used as controls. The medium levels in the donor pool were compared before and after a 4-h incubation.
-
We assessed the apparent permeability (Papp) of small-molecule SF to evaluate tight junction formation by BMECs. SF (100 μg/mL) was added to the upper chamber. A 100-µL volume was collected from the lower chamber at 30, 60, 90, and 120 min, and subsequently replenished with pre-equilibrated culture media. The absorbances of the samples were measured using a fluorescence spectrophotometer (Fluostar Omega; BMG Labtech, Offenburg, Germany). Papp was calculated as (dM/dt)/(A•C), where dM/dt represents the cumulative measured fluorescence intensity in the plate per unit time, A refers to the bottom area of the insert (1.12 cm2), and C refers to the SF intensity in the upper insert (941).
-
Cell viability was assessed using the cell counting kit-8 (CCK8) technique. For the cell counting kit-8 assay (AR1160, Boster), cultures were incubated in a culture medium containing 10% (v/v) cell counting kit-8 solution for 2 h. The absorbance was measured at 450 nm using a microplate reader (Biotek, Winooski, USA).
-
A phagocytosis assay kit (ab234054, Abcam) was used to quantify the phagocytic capacity of astrocytes in the NVU model. Following treatment, cells were incubated with 5 µL of prelabeled zymosan particles for 2 h and then washed by adding cold phagocytosis assay buffer. Subsequently, the cells were analyzed using a confocal fluorescence microscope (SP8 Leica microscope).
-
A reactive oxygen species assay kit (Yesen, Shanghai, China) was used to detect oxidative stress injury in neurons in the NVU model. Following treatment, cells were laser irradiated and subsequently stained with 2,7-dichlorodihydroflfluorescein diacetate (DCFH-DA, in green), followed by an incubation at 37 °C for 1 h. Finally, the cells were analyzed using a confocal fluorescence microscope (SP8 Leica microscope).
-
Neuronal apoptosis levels were assessed using the annexin V-FITC/PI double staining kit (E-CK-A211, Elabscience®). After the intervention, the neuronal cells were collected using EDTA-free trypsin, rinsed with PBS, and processed according to the manufacturer’s instructions. Briefly, cells were suspended in 100 μL of binding buffer, transferred to a centrifuge tube, 2.5 μL of dye was added, and the mixture was incubated for 30 min in the dark. Finally, 400 μL of binding buffer was added to the tube for flow cytometry detection.
-
Proteins were extracted from cells using RIPA lysis buffer (RIPA, AR0105, Boster), separated using SDS-PAGE (EpiZyme Biotechnology, China), and transferred to polyvinylidene fluoride (PVDF, HVLP09050, Millipore®) membranes. The PVDF membranes were incubated with primary antibodies at 4 °C overnight, followed by secondary antibodies for 1 h at 25 °C. Visualization was performed using a Bio-Rad CHEMIDoc XRS+ system. The primary antibodies used for protein analysis included AQP4 (1:1,000; ab259318, Abcam), LRP1 (1:1,000; ab92544, Abcam), occludin (1:1,000; ab216327, Abcam), melatonin receptor 1A (MTNR1A, 1:1,000; ab158923, Abcam), and β-Actin (1:10,000; ab8226, Abcam).
-
Cells were washed three times with PBS and fixed with 4% paraformaldehyde (PFA) at room temperature for 15 min. Cell membranes were permeabilized with 1% Triton X-100 for 20 min, followed by blocking of nonspecific binding sites with 5% BSA (Beyotime, Shanghai) for 1 h. Cultured cells were then incubated overnight at 4 °C with primary antibodies against AQP4 (1:100; ab259318; Abcam), LRP1 (1:100; ab92544; Abcam), occludin (1:100; ab216327; Abcam), and MTNR1A (1:100; ab158923; Abcam). The following day, fluorescent secondary antibodies were added dropwise to coverslips in a dark room and incubated for 1 h at room temperature, followed by staining of the nuclei with 4’,6-diamidino-2-phenylindole (DAPI, AR1176, Boster) for 30 min. Finally, the images were acquired and analyzed under a microscope.
-
All experiments were carried out at least three times and data were presented as means ± standard deviation (SD). Data analysis was performed using GraphPad Prism 8 software (San Diego, Calif., USA) or SPSS 18.0 statistical software (IBM Corp). Unpaired two-tailed Student’s t-tests (for two groups) or one-way ANOVA (for multiple groups) were used to assess statistical significance (P < 0.05).
-
The BBB function in the NVU model is shown in Figure 2. Four types of rat cerebral cells (BMECs, neurons, astrocytes, and microglia) were used to establish an NVU model (Figure 2A). Cell purity was assessed using immunofluorescence, which showed that the purity of BMECs, astrocytes, neurons, and microglia in the rat brain exceeded 92%, 96%, 97%, and 98%, respectively (Figure 2B). Subsequently, an in vitro NVU model was established, and the BBB properties were measured.
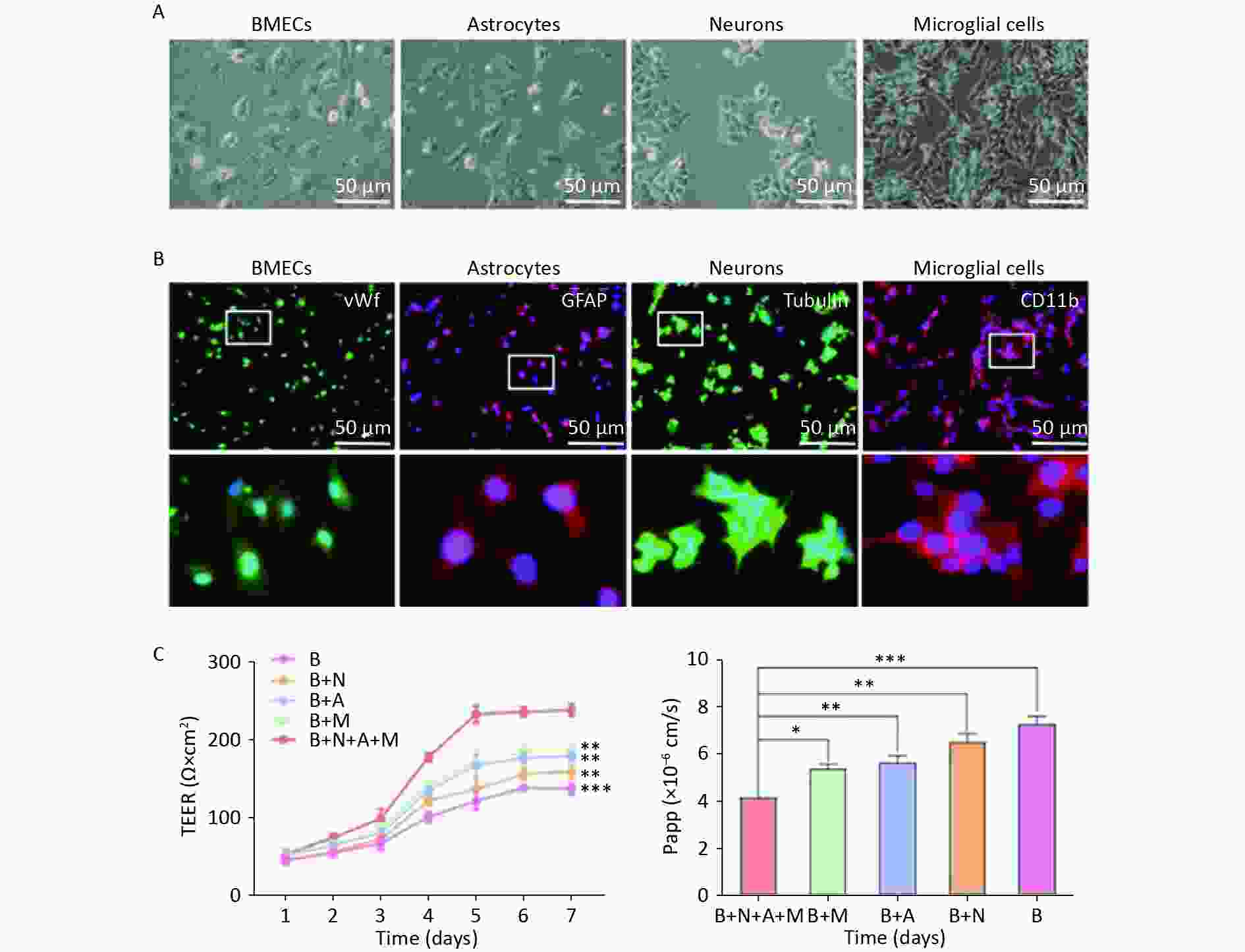
Figure 2. Functional features of the in vitro neurovascular unit (NVU) model. (A) Four-cell morphologies for the establishment of an in vitro NVU model. (B) Immunostaining for all cell types in the NVU model. Brain microvascular endothelial cells (BMECs) are stained for von Willebrand factor (vWF), astrocytes stained for glial fibrillary acidic protein (GFAP), neurons stained for Tubulin and microglial cells stained for CD11b. (C) Transendothelial electrical resistance (TEER) values for 7 days. (D) The permeability coefficient of sodium fluorescein in the B+N+A+M, B+M, B+A, B+N, and B groups. B+N+A+M group: BMECs cultured with neurons, astrocytes, and microglial cells; B+M group: BMECs cultured with microglial cells; B+A group: BMECs cultured with astrocytes; B+N group: BMECs cultured with neurons; B group: BMECs cultured alone in the transwell chamber. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001.
TEER measurements and the endothelial permeability coefficient for SF were used to compare the physical properties of the in vitro BBB model in various cultures. The NVU model achieved maximum TEER at 5 days, significantly exceeding the values in monoculture systems or in BMECs co-cultured with neurons, astrocytes, or microglial cells (Figure 2C). The low paracellular Papp of SF in the co-culture system confirmed the establishment of an impermeable barrier in vitro. The permeability coefficients of SF in the NVU system were notably minimal compared to those of the single culture of BMECs and BMECs co-cultured with neurons, astrocytes, or microglia (Figure 2D).
-
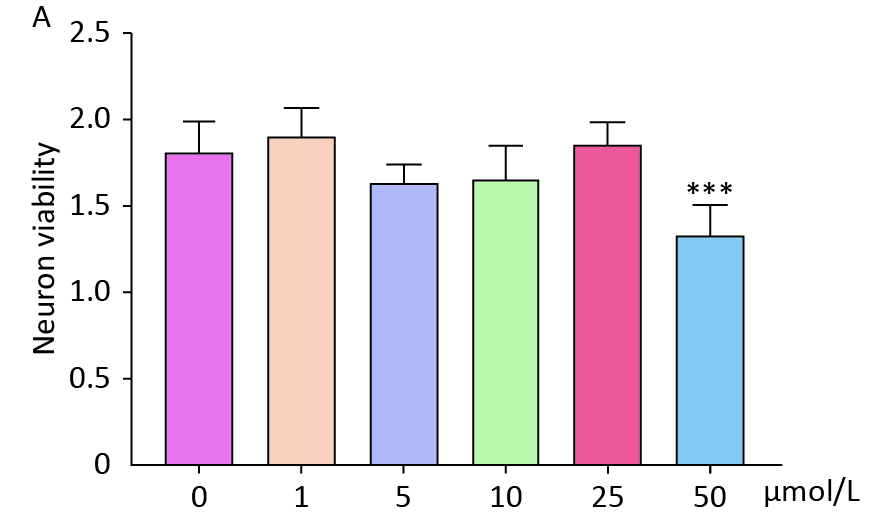
To investigate the impact of circadian rhythms on BBB function, we established an in vitro NVU model and co-cultured it with hemoglobin. The optimal concentration of hemoglobin for co-culture was determined to be 25 μmol/L (
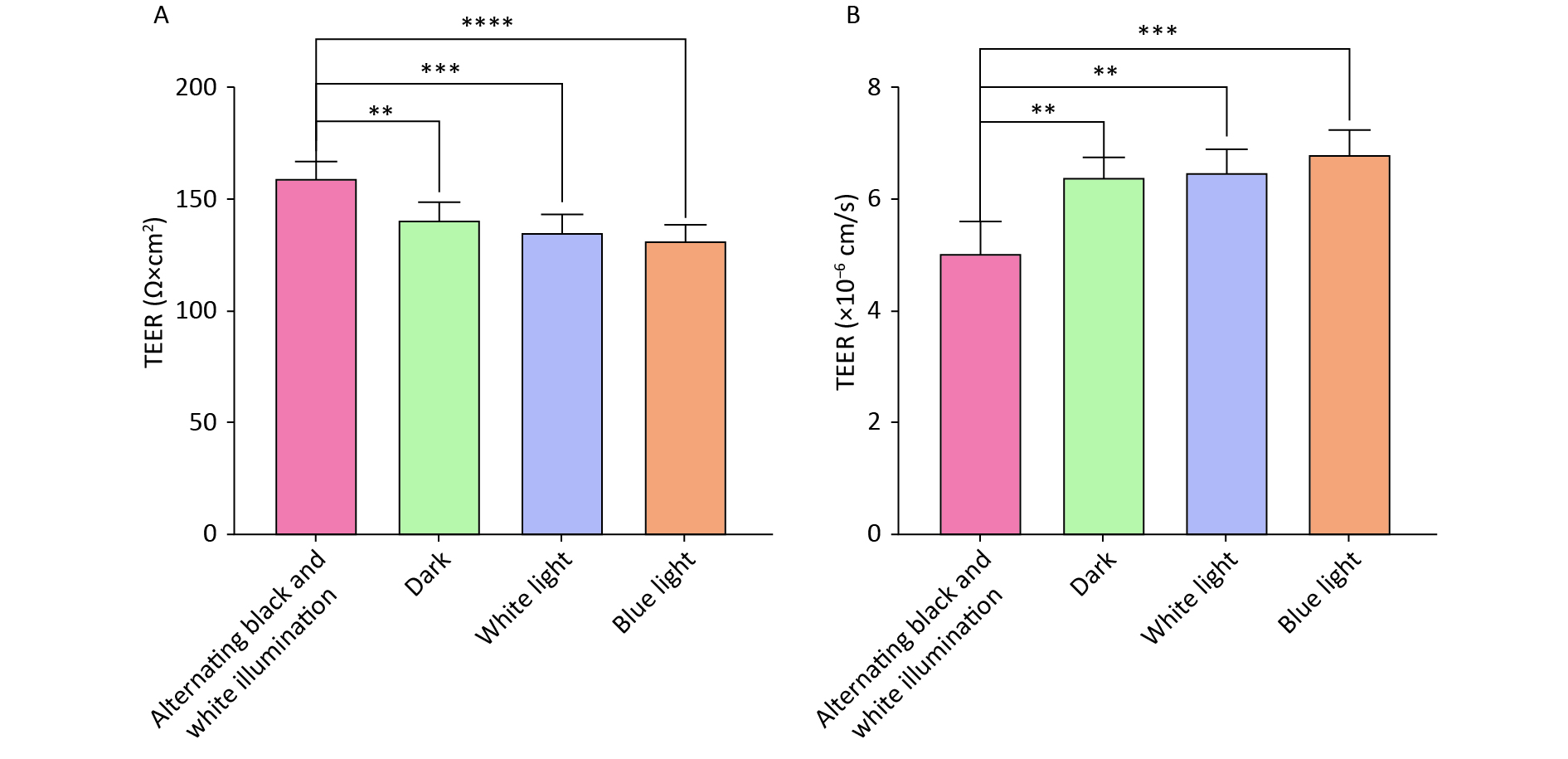
Supplementary Figure S1 , available in www.besjournal.com). Subsequently, the NVU co-cultured with hemoglobin was subjected to different illumination conditions (alternating black and white light, dark, white light, and blue light). The TEER values of the alternating black and white light group, which simulate circadian rhythm, were significantly higher than those of the other three groups (Figure 3A).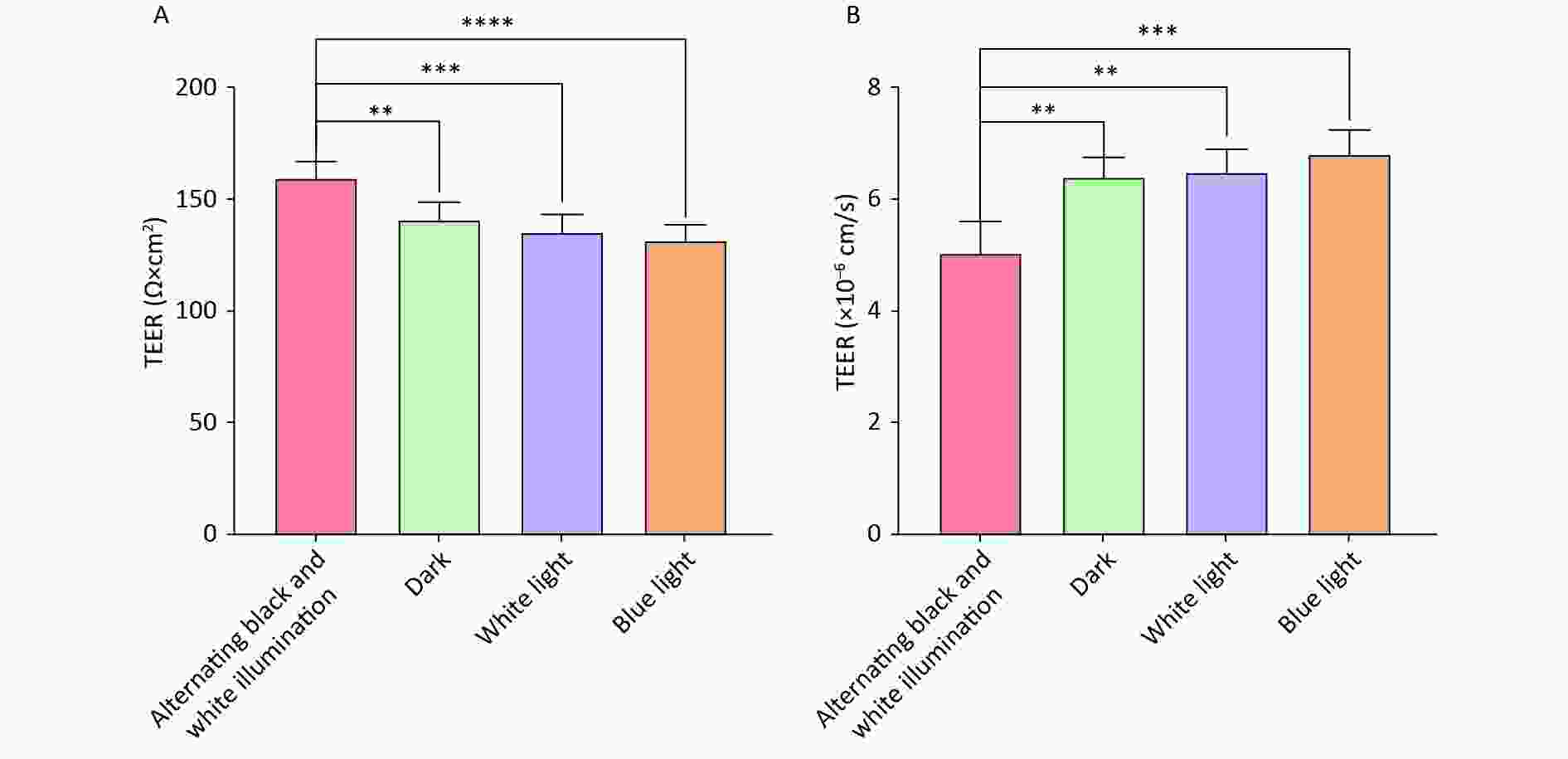
Figure 3. neurovascular unit (NVU) barrier function co-cultured with hemoglobin under different illumination. (A) The TEER values in the alternating black and white light, dark, white light, and blue light groups. (B) The permeability coefficient of sodium fluorescein in the alternating black and white light, dark, white light, and blue light groups. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
Furthermore, the low paracellular Papp of SF confirmed the establishment of an impermeable barrier in vitro. The permeability coefficients of SF in the alternating black and white light group were minimal compared to those of the dark, white light, and blue light groups (Figure 3B). These findings suggest that circadian rhythms can preserve the BBB function of the nervous system.
Circadian Rhythm Alleviates Neuronal Damage and Decreases Phagocytosis of Glial Cells in NVU Co-cultured with Hemoglobin
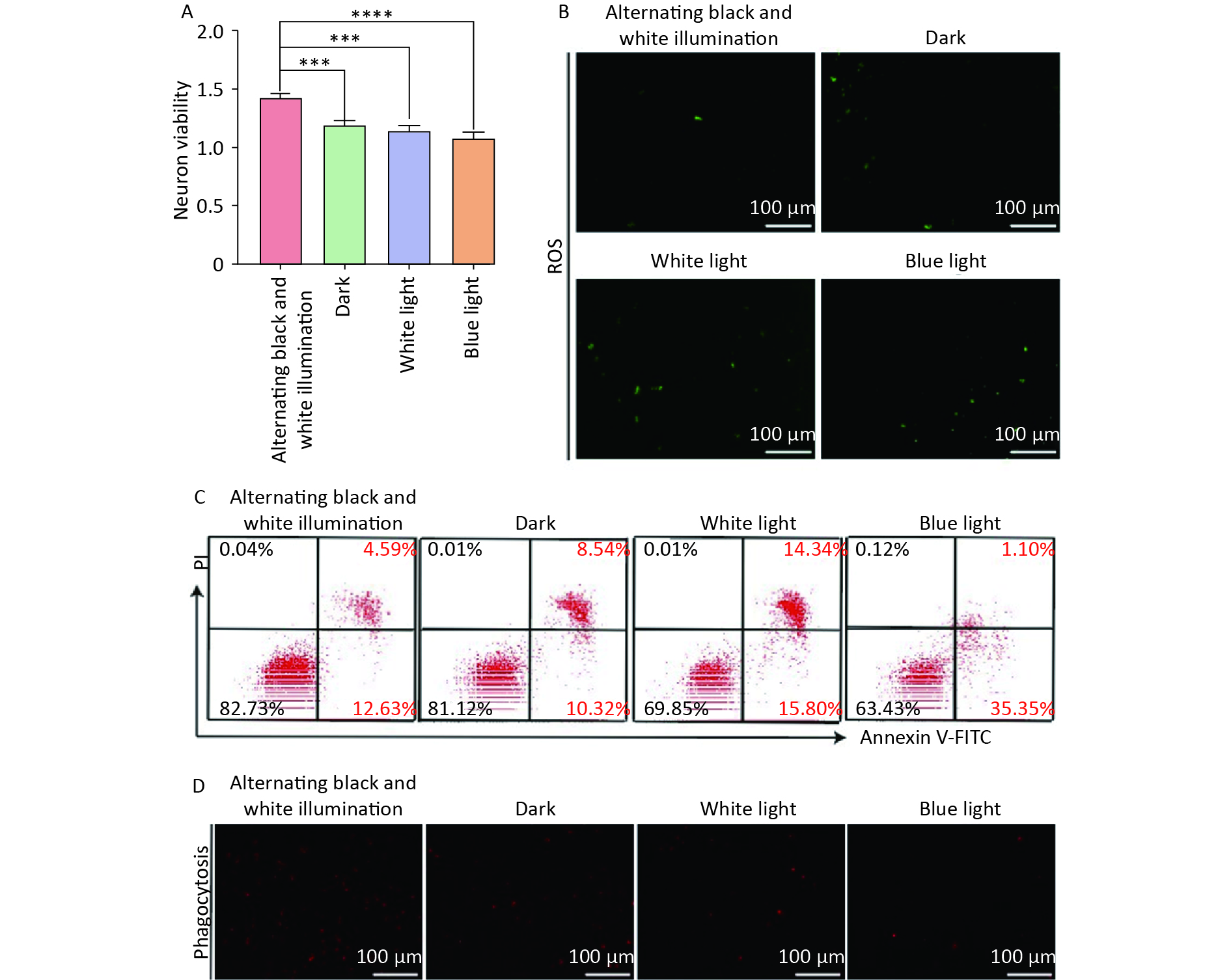
Subsequently, we investigated the effect of circadian rhythms on NVU function by treating NVU co-cultured with hemoglobin under different illumination conditions (alternating black and white light, dark, white light, and blue light). Our findings revealed that the alternating black and white light group showed the highest neuronal survival rate, along with the lowest levels of oxidative stress injury and apoptosis compared to the dark, white light, and blue light groups (Figure 4A–C).
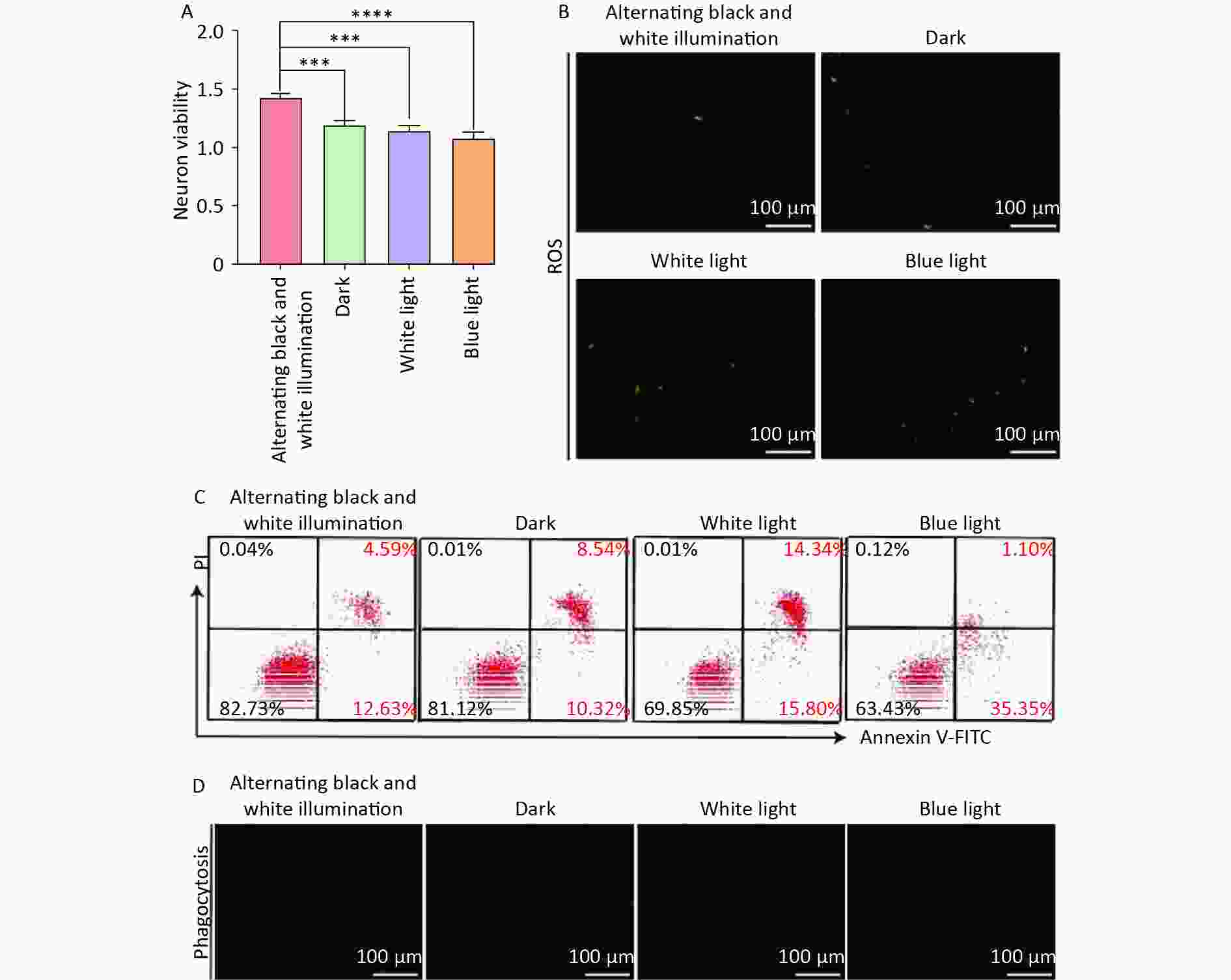
Figure 4. Neuron damage and microglia phagocytosis decreased in neurovascular unit (NVU) co-cultured with hemoglobin under different illumination. (A) The neuron survival rate in the alternating black and white light, dark, white light, and blue light groups. (B) Neuron oxidative stress injury in the alternating black and white light, dark, white light, and blue light groups. (C) The apoptosis rate of neurons in the alternating black and white light, dark, white light, and blue light groups. (D) Microglial phagocytosis in the alternating black and white light, dark, white light, and blue light groups. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
We also investigated the effects of different light irradiations on the phagocytosis of microglia in NVU co-cultured with hemoglobin. The results showed a significant inhibition of microglial phagocytosis in the dark, white light, and blue light groups compared to the alternating black and white light group (Figure 4D). These results highlight that circadian rhythm can mitigate oxidative stress injury and neuronal apoptosis, and decelerate the decline in microglial phagocytosis, thus protecting NVU function.
-
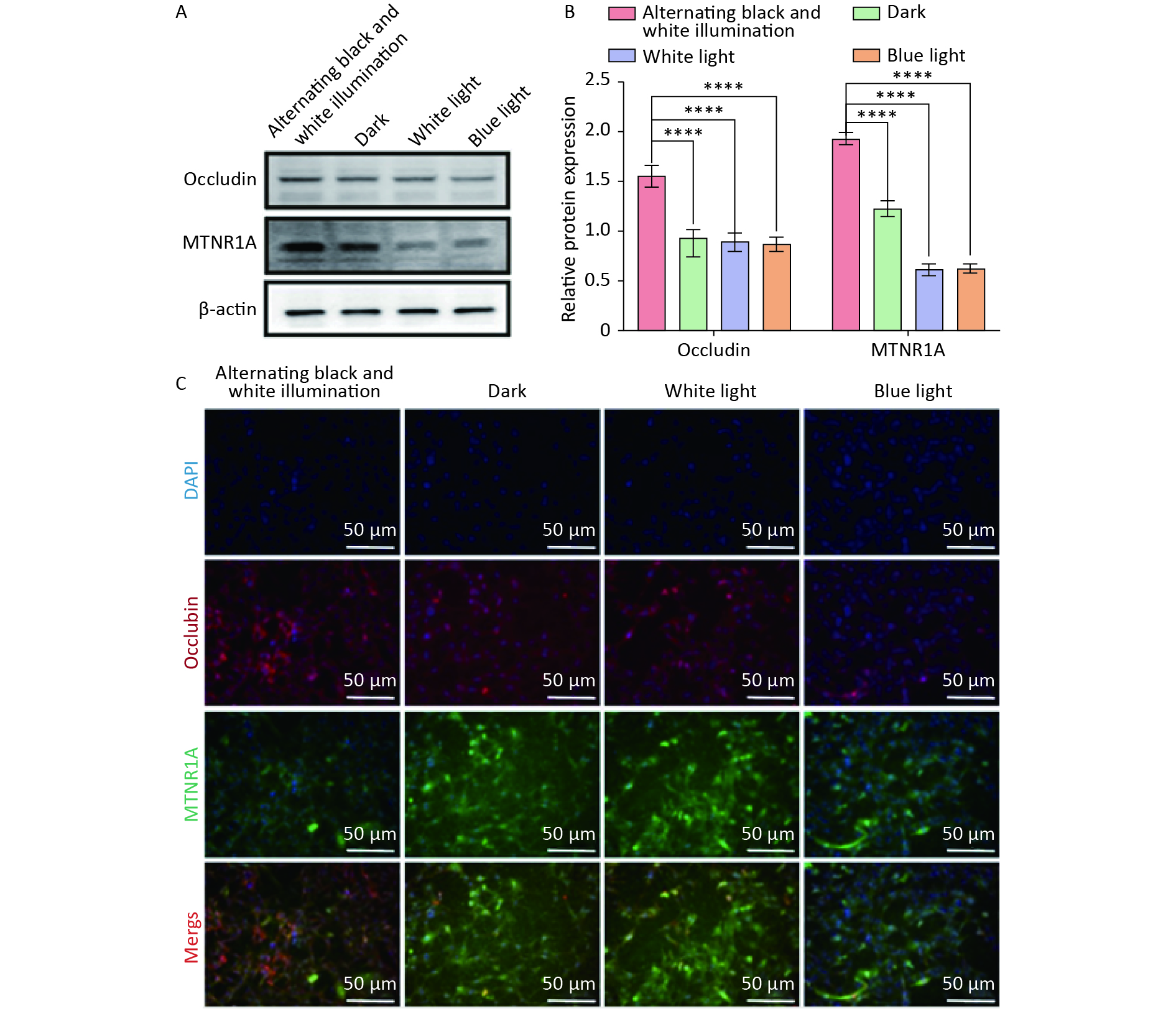
Occludin and MTNR1A are located at tight junctions of the BBB and play crucial roles in maintaining physiological functions. Subsequently, we examined the expression of occludin and MTNR1A in NVU co-cultured with hemoglobin under different light irradiation conditions. Western blot results indicated that the expression of occludin and MTNR1A in the dark, white light, and blue light groups was significantly reduced compared to the alternating black and white light group (Figure 5A, B). Similarly, immunofluorescence staining revealed a substantial decrease in the expression of occludin and MTNR1A in the dark, white light, and blue light groups compared to the alternating black and white light group (Figure 5C). These results suggest that circadian rhythm can mitigate the reduction in the expression of occludin and MTNR1A in NVU.
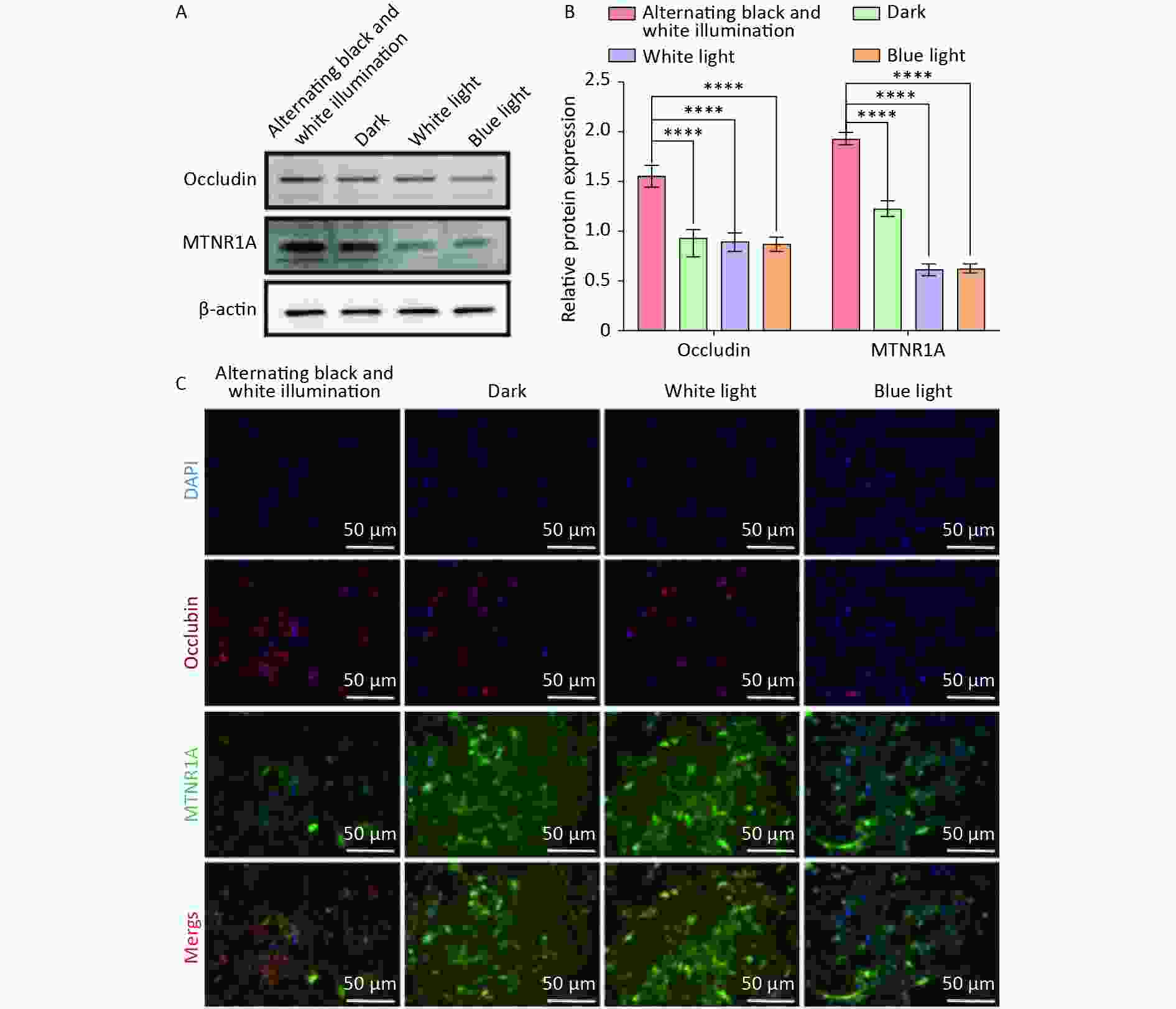
Figure 5. The expression of occludin and MTNR1A in neurovascular unit (NVU) co-cultured with hemoglobin under different illumination. (A, B) Western blot and quantification of occludin and MTNR1A in the alternating black and white light, dark, white light, and blue light groups, with β-actin as endogenous control. (C) Immunofluorescent staining of occludin and MTNR1A in the alternating black and white light, dark, white light, and blue light groups. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
Circadian Rhythm Preserved the Decrease in Expression of AQP4 and LRP1 in NVU Co-cultured with Hemoglobin.
AQP4 and LRP1 play crucial roles in eliminating metabolic waste and exogenous toxins from the brain, thus facilitating the removal of components of the hematoma after ICH. Finally, we investigated the expression of AQP4 and LRP1 in NVU co-cultured with hemoglobin under different light conditions. Western blot showed that the expression of AQP4 and LRP1 in the dark, white light, and blue light groups was significantly reduced compared to the alternating black and white light group (Figure 6A, B). Similarly, immunofluorescence staining revealed a substantial decrease in the expression of AQP4 and LRP1 in the dark, white light, and blue light groups compared to that of the alternating black and white light group (Figure 6C). These findings suggest that circadian rhythm may mitigate the reduction in AQP4 and LRP1 expression in the NVU.
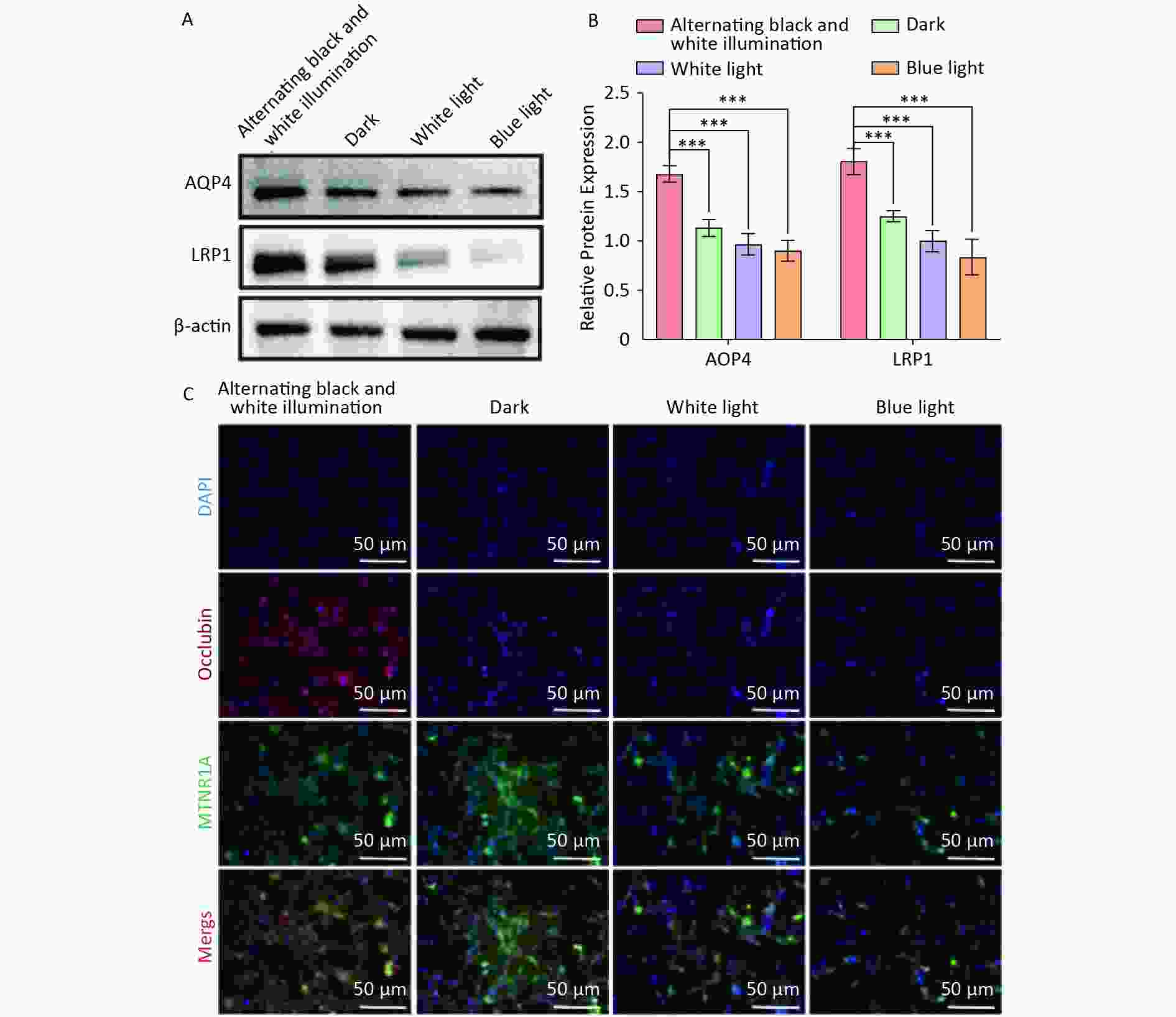
Figure 6. The expression of AQP4 and LRP1 in NVU co-cultured with hemoglobin under different illumination. (A, B) Western blot and quantification of AQP4 and LRP1 in the alternating black and white light, dark, white light, and blue light groups, with β-actin as endogenous control. (C) Immunofluorescent staining of AQP4 and LRP1 in the alternating black and white light, dark, white light, and blue light groups. aquaporin 4; LRP1, lipoprotein receptor-related protein 1.
-
Cerebral hemorrhage (CH) is a severe cerebrovascular disorder that is usually caused by the rupture of blood vessels in the brain. CH includes intracerebral hemorrhage (ICH), subarachnoid, and intraventricular hemorrhage[22]. Disruption of the BBB caused by ICH leads to vasogenic edema, resulting in extravasation of plasma proteins. This may potentially lead to neurotoxicity and the entry of vasoactive compounds into the brain[23,24]. Therefore, the cerebral clearance process plays a crucial role in maintaining homeostasis of the brain environment, limiting excessive inflammation, and promoting recovery[25,26].
The NVU is made up of various components, including neurons, astrocytes, microglial cells, BBB, and extracellular matrix, which collectively maintain the integrity of the brain tissue environment. Through neurovascular coupling, the NVU dynamically regulates local CBF and metabolic clearance in real time[27,28]. In recent years, research focusing on neurological disorders using the NVU as the primary investigative unit has become a prominent and influential area of study. The NVU model, established through a co-culture system involving primary BMECs, neurons, and astrocytes, has been used in research on various cerebrovascular diseases, including ischemic stroke[29,30]. To date, a limited number of studies have explored the use of triple-cell co-culture models to investigate the permeability of the BBB[20]. Building on previous studies, we used a four-cell co-culture model involving neurons, astrocytes, microglial cells, and BMECs to establish an NVU model. Additionally, compared with the monoculture of BMECs or BMECs co-cultured with the other three cell types, our four-cell co-culture model showed significantly higher TEER values and lower SF permeability. Our results suggest that the four-cell co-culture model established in this study not only recapitulates the diversity of NVU cell types, but also effectively simulates in vivo conditions.
Circadian rhythm is a biological process with rhythms of 24-h cycle that can influence neuropeptide release, pineal gland melatonin, and adrenal cortex glucocorticoid secretion[31,32]. Irregular eating and sleeping patterns, sudden changes in temperature, and prolonged exposure to blue light can disrupt circadian rhythms[33]. Certain molecules, such as tumor necrosis factor-alpha (TNF-α), β-amyloid, and prostaglandin D2 (PGD2), exhibit circadian rhythmic oscillations. These molecules are probably regulated according to rhythmic changes in BBB transport[34-36]. A study carried out in Drosophila revealed that the permeability of the BBB is regulated by circadian rhythms, with increased permeability observed at night[37]. In this study, we used alternating black and white light to simulate circadian rhythms. The experimental results revealed that, compared to individual treatments with dark light, white light, and blue light, the TEER values of the NVU model subjected to alternating black-white light were significantly higher, with a minimal SF permeability coefficient. Our findings consistently suggest that circadian rhythms can effectively preserve BBB function.
-
Neurons play a pivotal role in initiating local vascular responses in NVU structures. Neurons modulate CBF by generating signaling molecules such as glutamate and adenosine. These signaling molecules exert direct or indirect effects on local blood vessels by inducing vasodilation or vasoconstriction[7]. In the hours following ICH, activated microglia and infiltrating immune cells play a crucial role in protecting surrounding cells and tissues by clearing blood and damaged neurons[38]. Insufficient clearance capability of microglial cells can prolong damage and further deteriorate the patient’s condition[39]. In our experiments, we observed that the neuronal survival rate was the highest in the hemoglobin co-culture NVU model subjected to alternating black and white light treatment, with lower levels of oxidative stress damage and apoptosis. Furthermore, compared to the group exposed to alternating black and white light, the phagocytic activity of microglial cells in the dark, white light, and blue light groups was significantly inhibited. These findings suggest that circadian rhythms can alleviate oxidative stress damage and apoptosis in neurons and improve the phagocytic capability of microglial cells, thus protecting the normal function of the NVU.
Occludin is present at the tight junctions of the BBB and plays a crucial role in maintaining the physiological function of the BBB, serving as a specific indicator of BBB integrity. The disruption of the BBB during ICH leads to decreased occludin expressions[40]. Melatonin, produced by the pineal gland, serves as an effector of the circadian rhythm-biological clock and plays a crucial role in circadian regulation[41]. An animal experimental study revealed that the application of melatonin in transgenic Alzheimer’s disease animal models improves the glial lymphatic clearance capacity for Aβ, consequently reducing the deposition of Aβ in the brain[42]. Therefore, we hypothesized that circadian rhythms influence BBB function through modulation of melatonin secretion. The results of this study revealed that alternating black-white light treatment significantly improved the expression of occludin and melatonin receptor 1A (MTNR1A) in the NVU, suggesting that circadian rhythms regulate the integrity of BBB function, possibly through the modulation of melatonin.
AQP4 is expressed in astrocytic foot processes and functions as a gate to control the entry of cerebrospinal fluid into the brain parenchyma for metabolic exchange[11]. Studies have revealed that the inhibition of AQP4 leads to a reduction in the clearance capacity of the brain, highlighting the critical role of AQP4 in the elimination of cerebral metabolic waste and exogenous toxins[43]. Convective activities of the glymphatic system stimulated by AQP4 enhance the clearance function of the BBB, particularly in astrocytes[44]. LRP1, which serves as the main endocytic receptor in perivascular glial cells of the BBB, is also a main scavenger receptor that facilitates the clearance of Aβ in the BBB[45]. In previous studies, we have shown that LRP1 plays a crucial role in promoting the clearance of components of the hematoma after ICH[14,46]. This study, for the first time, revealed that alternating black-white light treatment significantly rescued the decreased expression of AQP4 and LRP1 in NVU co-cultured with hemoglobin. These findings suggest that circadian rhythms can mitigate the reduction in the expression of AQP4 and LRP1 in NVU.
In summary, this study established an NVU model through a four-cell co-culture, simulating the BBB environment by co-culturing with hemoglobin. Using alternating black-white light treatment in the NVU model mimicked circadian rhythms, and it was observed that circadian rhythms could protect BBB integrity and enhance the ability of the endogenous brain clearance system to remove hematomas after ICH. This study focused on the NVU as the research unit, unraveling the role of circadian rhythm in the function of the brain clearance system and providing new ideas to improve the brain clearance ability of patients after ICH.
-
Sprague-Dawley (SD) rats were obtained from the Experimental Animal Center of Shanxi Medical University (Shanxi, China). All procedures involving the use of SD rats were approved by the Ethics Committee of the Animal Transformation Center of Shanxi Medical University (No. DW2022029).
-
The authors declare that they have no competing interests.
-
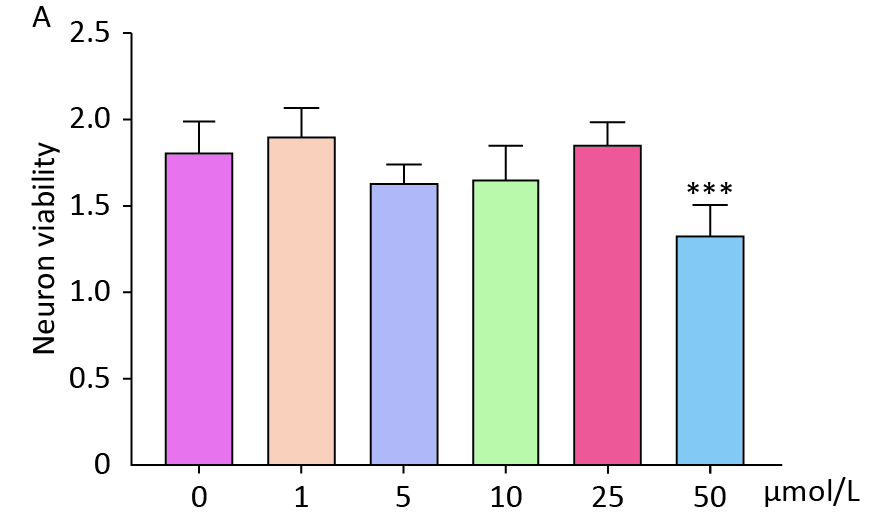
Figure S1. Effects of different concentrations of hemoglobin on neuronal cell activity. (A) CCK8 results showed that different concentrations of hemoglobin affected the activity of neurons. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
doi: 10.3967/bes2024.090
The Regulatory Role and Mechanism of Circadian Rhythm in Hemoglobin Co-cultured Neurovascular Unit
-
Abstract:
Objective Intracranial hemorrhage (ICH), the second most common subtype of stroke, exacerbates the disruption of the blood-brain barrier (BBB), leading to vasogenic edema, plasma protein extravasation, and infiltration of neurotoxic substances. The clearance capacity of the brain plays a crucial role in maintaining BBB homeostasis and facilitating patient recovery after hemorrhage. This study aimed to investigate the effect of circadian rhythms on BBB function, neuronal damage, and clearance capabilities. Methods The transwell model and hemoglobin were co-cultured to simulate the BBB environment after ICH. After intervention with different light groups, neuronal apoptosis was determined, glial phagocytosis was analyzed, the expression of endogenous clearing-related proteins aquaporin 4 (AQP4) and low-density lipoprotein receptor-related protein 1 (LRP1) was detected by western blotting and immunofluorescence dual standard method, and the expression of the tight junction protein occludin and melatonin receptor 1A (MTNR1A) was quantitatively analyzed. Results Circadian rhythms play a key role in maintaining the integrity of the BBB, reducing oxidative stress-induced neuronal damage, and improving microglial phagocytosis. Meanwhile, the expression of occludin and MTNR1A in neurovascular unit (NVU) co-cultured with hemoglobin improved the expression of AQP4 and LRP1, the key proteins in the NVU’s endogenous brain clearance system. Conclusion Circadian rhythm (alternating black and white light) protects the NVU BBB function after ICH, promotes the expression of proteins related to the clearance of the hematoma, provides new evidence for the clinical treatment of patients recovering from ICH, and improves the circadian rhythm to promote brain metabolism and hematoma clearance. -
Key words:
- Blood-brain barrier /
- Circadian rhythm /
- Neurovascular unit /
- Melatonin receptor 1A /
- Aquaporin-4
-
Figure 1. Establishment of an in vitro NVU model. (A) Schematic drawing of the four-cell co-culture system. (B) The procedure for establishing an in vitro four-cell co-culture model. NUV, neurovascular unit; BMECs, brain microvascular endothelial cells.
Figure 2. Functional features of the in vitro neurovascular unit (NVU) model. (A) Four-cell morphologies for the establishment of an in vitro NVU model. (B) Immunostaining for all cell types in the NVU model. Brain microvascular endothelial cells (BMECs) are stained for von Willebrand factor (vWF), astrocytes stained for glial fibrillary acidic protein (GFAP), neurons stained for Tubulin and microglial cells stained for CD11b. (C) Transendothelial electrical resistance (TEER) values for 7 days. (D) The permeability coefficient of sodium fluorescein in the B+N+A+M, B+M, B+A, B+N, and B groups. B+N+A+M group: BMECs cultured with neurons, astrocytes, and microglial cells; B+M group: BMECs cultured with microglial cells; B+A group: BMECs cultured with astrocytes; B+N group: BMECs cultured with neurons; B group: BMECs cultured alone in the transwell chamber. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001.
Figure 3. neurovascular unit (NVU) barrier function co-cultured with hemoglobin under different illumination. (A) The TEER values in the alternating black and white light, dark, white light, and blue light groups. (B) The permeability coefficient of sodium fluorescein in the alternating black and white light, dark, white light, and blue light groups. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 4. Neuron damage and microglia phagocytosis decreased in neurovascular unit (NVU) co-cultured with hemoglobin under different illumination. (A) The neuron survival rate in the alternating black and white light, dark, white light, and blue light groups. (B) Neuron oxidative stress injury in the alternating black and white light, dark, white light, and blue light groups. (C) The apoptosis rate of neurons in the alternating black and white light, dark, white light, and blue light groups. (D) Microglial phagocytosis in the alternating black and white light, dark, white light, and blue light groups. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 5. The expression of occludin and MTNR1A in neurovascular unit (NVU) co-cultured with hemoglobin under different illumination. (A, B) Western blot and quantification of occludin and MTNR1A in the alternating black and white light, dark, white light, and blue light groups, with β-actin as endogenous control. (C) Immunofluorescent staining of occludin and MTNR1A in the alternating black and white light, dark, white light, and blue light groups. Data are expressed as mean ± SD. **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 6. The expression of AQP4 and LRP1 in NVU co-cultured with hemoglobin under different illumination. (A, B) Western blot and quantification of AQP4 and LRP1 in the alternating black and white light, dark, white light, and blue light groups, with β-actin as endogenous control. (C) Immunofluorescent staining of AQP4 and LRP1 in the alternating black and white light, dark, white light, and blue light groups. aquaporin 4; LRP1, lipoprotein receptor-related protein 1.
-
[1] Owolabi MO, Thrift AG, Mahal A, et al. Primary stroke prevention worldwide: translating evidence into action. Lancet Public Health, 2022; 7, e74−85. doi: 10.1016/S2468-2667(21)00230-9 [2] Saldana S, Breslin J, Hanify J, et al. Comparison of clevidipine and nicardipine for acute blood pressure reduction in hemorrhagic stroke. Neurocrit Care, 2022; 36, 983−92. doi: 10.1007/s12028-021-01407-w [3] Xiao MJ, Xiao ZJ, Yang BB, et al. Blood-brain barrier: more contributor to disruption of central nervous system homeostasis than victim in neurological disorders. Front Neurosci, 2020; 14, 764. doi: 10.3389/fnins.2020.00764 [4] Cui Y, Zhao Y, Chen SY, et al. Association of serum biomarkers with post-thrombolytic symptomatic intracranial hemorrhage in stroke: a comprehensive protein microarray analysis from INTRECIS study. Front Neurol, 2022; 13, 751912. doi: 10.3389/fneur.2022.751912 [5] Schaeffer S, Iadecola C. Revisiting the neurovascular unit. Nat Neurosci, 2021; 24, 1198−209. doi: 10.1038/s41593-021-00904-7 [6] McConnell HL, Mishra A. Cells of the blood-brain barrier: an overview of the neurovascular unit in health and disease. In: Stone N. The Blood-Brain Barrier: Methods and Protocols. Humana. 2022, 3-24. [7] Dormanns K, Brown RG, David T. Neurovascular coupling: a parallel implementation. Front Comput Neurosci, 2015; 9, 109. [8] Iadecola C. The neurovascular unit coming of age: a journey through neurovascular coupling in health and disease. Neuron, 2017; 96, 17−42. doi: 10.1016/j.neuron.2017.07.030 [9] Liu JC, Guo YZ, Zhang CY, et al. Clearance systems in the brain, from structure to function. Front Cell Neurosci, 2022; 15, 729706. doi: 10.3389/fncel.2021.729706 [10] Nedergaard M, Goldman SA. Glymphatic failure as a final common pathway to dementia. Science, 2020; 370, 50−6. doi: 10.1126/science.abb8739 [11] Rasmussen MK, Mestre H, Nedergaard M. The glymphatic pathway in neurological disorders. Lancet Neurol, 2018; 17, 1016−24. doi: 10.1016/S1474-4422(18)30318-1 [12] Kitchen P, Salman MM, Halsey AM, et al. Targeting aquaporin-4 subcellular localization to treat central nervous system edema. Cell, 2020; 181, 784-99. e19. [13] Qing WG, Dong YQ, Ping TQ, et al. Brain edema after intracerebral hemorrhage in rats: the role of iron overload and aquaporin 4. J Neurosurg, 2009; 110, 462−8. doi: 10.3171/2008.4.JNS17512 [14] Wang GQ, Manaenko A, Shao AW, et al. Low-density lipoprotein receptor-related protein-1 facilitates heme scavenging after intracerebral hemorrhage in mice. J Cereb Blood Flow Metab, 2017; 37, 1299−310. doi: 10.1177/0271678X16654494 [15] Wang GQ, Guo ZN, Tong LS, et al. TLR7 (toll-like receptor 7) facilitates heme scavenging through the BTK (Bruton tyrosine kinase)-CRT (calreticulin)-LRP1 (low-density lipoprotein receptor-related protein-1)-Hx (hemopexin) pathway in murine intracerebral hemorrhage. Stroke, 2018; 49, 3020−9. doi: 10.1161/STROKEAHA.118.022155 [16] Mitchell JW, Gillette MU. Development of circadian neurovascular function and its implications. Front Neurosci, 2023; 17, 1196606. doi: 10.3389/fnins.2023.1196606 [17] Hablitz LM, Plá V, Giannetto M, et al. Circadian control of brain glymphatic and lymphatic fluid flow. Nat Commun, 2020; 11, 4411. doi: 10.1038/s41467-020-18115-2 [18] Cardinali DP. Melatonin and healthy aging. Vitam Horm, 2021; 115, 67−88. [19] Wahl S, Engelhardt M, Schaupp P, et al. The inner clock-blue light sets the human rhythm. J Biophotonics, 2019; 12, e201900102. doi: 10.1002/jbio.201900102 [20] Xue Q, Liu Y, Qi HY, et al. A novel brain neurovascular unit model with neurons, astrocytes and microvascular endothelial cells of rat. Int J Biol Sci, 2013; 9, 174−89. doi: 10.7150/ijbs.5115 [21] Li CX, Wang XQ, Cheng FF, et al. Hyodeoxycholic acid protects the neurovascular unit against oxygen-glucose deprivation and reoxygenation-induced injury in vitro. Neural Regen Res, 2019; 14, 1941−9. doi: 10.4103/1673-5374.259617 [22] Keep RF, Andjelkovic AV, Xiang JM, et al. Brain endothelial cell junctions after cerebral hemorrhage: changes, mechanisms and therapeutic targets. J Cereb Blood Flow Metab, 2018; 38, 1255−75. doi: 10.1177/0271678X18774666 [23] Stokum JA, Gerzanich V, Simard JM. Molecular pathophysiology of cerebral edema. J Cereb Blood Flow Metab, 2016; 36, 513−38. doi: 10.1177/0271678X15617172 [24] Chen SP, Li LZ, Peng C, et al. Targeting oxidative stress and inflammatory response for blood-brain barrier protection in intracerebral hemorrhage. Antioxid Redox Signal, 2022; 37, 115−34. doi: 10.1089/ars.2021.0072 [25] Da Mesquita S, Papadopoulos Z, Dykstra T, et al. Meningeal lymphatics affect microglia responses and anti-Aβ immunotherapy. Nature, 2021; 593, 255−60. doi: 10.1038/s41586-021-03489-0 [26] Liu LR, Liu JC, Bao JS, et al. Interaction of microglia and astrocytes in the neurovascular unit. Front Immunol, 2020; 11, 1024. doi: 10.3389/fimmu.2020.01024 [27] Tiret P, Chaigneau E, Lecoq J, et al. Two-photon imaging of capillary blood flow in olfactory bulb glomeruli. In: Hyder F. Dynamic Brain Imaging: Multi-Modal Methods and In Vivo Applications. Humana. 2009, 81-91. [28] LeDoux JE, Thompson ME, Iadecola C, et al. Local cerebral blood flow increases during auditory and emotional processing in the conscious rat. Science, 1983; 221, 576−8. doi: 10.1126/science.6867731 [29] Zhang WJ, Zhao XJ, Qi XW, et al. Induced pluripotent stem cell model revealed impaired neurovascular interaction in genetic small vessel disease cerebral autosomal dominant arteriopathy with subcortical infarcts and leukoencephalopathy. Front Cell Neurosci, 2023; 17, 1195470. doi: 10.3389/fncel.2023.1195470 [30] Wang C, Gu W, Zhao K, et al. A new neurotoxicity model composed of a neurovascular unit in vitro. Cell Mol Biol, 2016; 62, 21−6. doi: 10.14715/cmb/2016.62.13.5 [31] Greco CM, Sassone-Corsi P. Circadian blueprint of metabolic pathways in the brain. Nat Rev Neurosci, 2019; 20, 71−82. [32] Neves AR, Albuquerque T, Quintela T, et al. Circadian rhythm and disease: relationship, new insights, and future perspectives. J Cell Physiol, 2022; 237, 3239−56. doi: 10.1002/jcp.30815 [33] Deaver JA, Eum SY, Toborek M. Circadian disruption changes gut microbiome taxa and functional gene composition. Front Microbiol, 2018; 9, 737. doi: 10.3389/fmicb.2018.00737 [34] Kress GJ, Liao F, Dimitry J, et al. Regulation of amyloid-β dynamics and pathology by the circadian clock. J Exp Med, 2018; 215, 1059−68. doi: 10.1084/jem.20172347 [35] Yang DF, Huang WC, Wu CW, et al. Acute sleep deprivation exacerbates systemic inflammation and psychiatry disorders through gut microbiota dysbiosis and disruption of circadian rhythms. Microbiol Res, 2023; 268, 127292. doi: 10.1016/j.micres.2022.127292 [36] Kawaguchi C, Shintani N, Hayata-Takano A, et al. Lipocalin-type prostaglandin D synthase regulates light-induced phase advance of the central circadian rhythm in mice. Commun Biol, 2020; 3, 557. doi: 10.1038/s42003-020-01281-w [37] Zhang SL, Yue ZF, Arnold DM, et al. A circadian clock in the blood-brain barrier regulates xenobiotic efflux. Cell, 2018; 173, 130-9. e10. [38] Lan X, Han XN, Li Q, et al. (−)-Epicatechin, a natural flavonoid compound, protects astrocytes against hemoglobin toxicity via Nrf2 and AP-1 signaling pathways. Mol Neurobiol, 2017; 54, 7898−907. doi: 10.1007/s12035-016-0271-y [39] Li Q, Lan X, Han XN, et al. Microglia-derived interleukin-10 accelerates post-intracerebral hemorrhage hematoma clearance by regulating CD36. Brain Behav Immun, 2021; 94, 437−57. doi: 10.1016/j.bbi.2021.02.001 [40] Feldman GJ, Mullin JM, Ryan MP. Occludin: structure, function and regulation. Adv Drug Deliv Rev, 2005; 57, 883−917. doi: 10.1016/j.addr.2005.01.009 [41] Spinedi E, Cardinali DP. Neuroendocrine-metabolic dysfunction and sleep disturbances in neurodegenerative disorders: focus on Alzheimer's disease and melatonin. Neuroendocrinology, 2019; 108, 354−64. doi: 10.1159/000494889 [42] Li YM, Zhang J, Wan JL, et al. Melatonin regulates Aβ production/clearance balance and Aβ neurotoxicity: a potential therapeutic molecule for Alzheimer's disease. Biomed Pharmacother, 2020; 132, 110887. doi: 10.1016/j.biopha.2020.110887 [43] Zhao ZA, Li P, Ye SY, et al. Perivascular AQP4 dysregulation in the hippocampal CA1 area after traumatic brain injury is alleviated by adenosine A2A receptor inactivation. Sci Rep, 2017; 7, 2254. doi: 10.1038/s41598-017-02505-6 [44] Semyachkina-Glushkovskaya OV, Karavaev AS, Prokhorov MD, et al. EEG biomarkers of activation of the lymphatic drainage system of the brain during sleep and opening of the blood-brain barrier. Comput Struct Biotechnol J, 2023; 21, 758−68. doi: 10.1016/j.csbj.2022.12.019 [45] Ma QY, Zhao Z, Sagare AP, et al. Blood-brain barrier-associated pericytes internalize and clear aggregated amyloid-β42 by LRP1-dependent apolipoprotein E isoform-specific mechanism. Mol Neurodegener, 2018; 13, 57. doi: 10.1186/s13024-018-0286-0 [46] Wang GQ, Li T, Duan SN, et al. PPAR-γ promotes hematoma clearance through haptoglobin-hemoglobin-CD163 in a rat model of intracerebral hemorrhage. Behav Neurol, 2018; 2018, 7646104. -
 24041NEW+Supplementary Materials.pdf
24041NEW+Supplementary Materials.pdf

-

 点击查看大图
点击查看大图
计量
- 文章访问数: 1101
- HTML全文浏览量: 491
- PDF下载量: 72
- 被引次数: 0

 Quick Links
Quick Links