

 下载:
下载:

-
Tuberculosis (TB) is a serious infectious disease and the ninth leading cause of death worldwide. Therefore, it remains one of the most important public health problems in the world. According to the Global TB Report 2019, there were approximately 10 million active cases of TB worldwide in 2018, of which approximately 1.24 million died of the disease. In the meantime, a quarter of the world’s population has been latently infected with Mycobacterium tuberculosis[1]. Furthermore, co-infection of M. tuberculosis and human immunodeficiency virus (HIV), prevalence of multidrug-resistant TB (MDR-TB) and emergence of extensively drug-resistant TB (XDR-TB), controlling the current TB epidemic remains a severe challenge[2]. Vaccination is the most efficient method for controlling communicable diseases; the Bacillus Calmette-Guérin (BCG) vaccine has been the first and only licensed vaccine used for TB prevention since 1921 globally. It was developed after long-term subculture of M. bovis, whereby the cells almost completely lost all pathogenicity to humans, but maintained strong immunogenicity, making it an ideal vaccine at the time[3]. According to current statistics, more than 3 billion people have been vaccinated with the BCG vaccine worldwide; however, its immune protection effect has always been a controversial topic. It is generally considered that the BCG vaccine is effective for the prevention and control of TB, especially tuberculous meningitis in infants, while its immune protection against adult TB is inferior or even ineffective[4]. Therefore, there is an urgent need to develop new and effective TB vaccines to end the epidemic, and researchers worldwide are working towards this goal using different approaches[5-7]. The key to these different approaches depends on finding new and effective antigens[8-11]. The effectiveness of the BCG vaccine against TB depends on the cross-reactivity of immune responses induced by both M. bovis and M. tuberculosis in humans[4]. Therefore, scientists have hypothesized that identifying new antigens from non-tuberculous mycobacteria (NTM) such as M. vaccae is a promising approach for determining suitable antigens for use in vaccines[12, 13]. M. intracellulare, named by Runyon in the 1960s, is one of the most common pathogenic NTM and is widely distributed worldwide. M. intracellulare infection usually causes lung damage in the host. M. intracellulare, M. avium, and M. tuberculosis have been shown to induce greater, albeit similar, levels of chemokines compared to those induced by M. smegmatis and M. abscessus, whereas M. intracellulare showed higher levels of phagosome-lysosome fusion and apoptosis than M. tuberculosis in macrophages in vitro[14]. However, studies on the immune mechanisms induced by M. intracellulare infection are required, especially to characterize the cross-immunity with M. tuberculosis.
In this study, we aimed to identify antigens that induce cross-immunity between M. intracellulare and M. tuberculosis. First, we used whole bacterial protein extracts from M. intracellulare to immunize mice and evaluated the antigen performance using cellular and humoral immunoassays and the capability of macrophages to control intracellular M. tuberculosis growth. We then determined cross-reactive antigens between M. intracellulare and M. tuberculosis by comparative genomic analysis and immunoproteome microarray hybridization. The results of this study will help identify effective antigens among M. intracellulare proteins and may provide potential candidates for the development of new types of TB vaccines and immunotherapies against M. intracellulare and M. tuberculosis infection.
-
All animal experiments conducted in the study were approved by the Ethics Committee and the Animal Experimental Ethical Committee of the National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention.
-
M. intracellulare (ATCC13950) was cultured in Löwenstein–Jensen medium (ENCODE, China) at 37 °C. The bacterial colonies were washed in PBS buffer three times before being harvested into lysis buffer (10 mmol/L Tris-HCl + 100 mmol/L NaH2P04 + 8 mol/L urea + 50 mmol/L IAA + 1× protease inhibitor cocktail). The harvested bacteria were then shock-crushed with magnetic beads four times (6.5 m/s, 2 min, ice for 1 min) to release the whole bacterial protein extract. Protein concentration was determined using a BCA kit (TransGen Biotech, China). All protein products were stored at −80 °C until use.
-
Female BALB/c mice at 6–8 weeks were used in this study. Five mice were included in both experimental group and control group, and at least three mice were included to ensure reliability and statistical analysis. The experimental group was immunized with a 200 μL mixture containing 100 μg whole M. intracellulare protein extract, 50 μL 0.5 mg/mL PolyI:C, and 100 μL 2.5 mg/mL Dimethyl-dioctyldecylammonium bromide (DDA) as adjuvants. The control group was immunized with adjuvant mixture only (50 μL 0.5 mg/mL PolyI:C and 100 μL 2.5 mg/mL DDA). Each mouse was immunized 3 times, with subcutaneous injections at ten-day intervals.
-
On the 10th day after the last vaccination, spleens from immunized mice were surgically removed, crushed with a syringe plunger into Roswell Park Memorial Institute (RPMI) 1,640 medium (with 1% penicillin-streptomycin) (Gibco, USA), and filtered into centrifuge tubes with a cell strainer. Cell suspensions were centrifuged at 1,000 rpm (r = 8.5 cm) for 5 min, and erythrocytes were lysed with ACK lysis buffer (Solarbio, China). Finally, the splenocytes were collected and quantified using Millipore ScepterTM 2.0 (merckmillipore, Germany).
For each well, splenocytes (2 × 106 cells/mL, 500 μL) were co-cultured with 10 μg of corresponding bacterial antigens for 48 h. RPMI 1640 + ConA (5 μg/mL) or just RPMI 1640 were used as positive and negative controls, respectively. The BD OptEIA ELISA kits were used for detection of cytokines, including IFN-γ, IL-2, IL-4, IL-6, and IL-12, based on absorbance at 450 nm.
-
Ten days after the first, second, and third immunization, 120 μL mouse blood was collected from the orbital vein. Blood samples were stored at 4 °C overnight and then centrifuged at 2,000 rpm (r = 8.5 cm) for 10 min. The sera were collected and stored at −80 °C. ELISA was performed as follows: The 96-well ELISA plates were coated with antigens (the whole bacterial protein extract) at a final concentration of 10 μg/mL at 4 °C overnight. Blocking was achieved by adding 200 μL 3% BSA (Saibao, China) in each well, and extracts were incubated at 37 °C for 2 h. Sera (diluted 1:100, 1:1,000, 1:10,000, 2:10,000, 4:10,000, 8:10,000, and 1:100,000) were added and incubated for 1 h. Next, 100 μL 1:3,000 diluted secondary antibody (Southern Biotech, USA) was added and incubated at 37 °C for 50 min. TMB substrate (100 μL/well) (InnoReagents, China) was then added and the reaction was terminated by adding 2 mol/L H2SO4 (50 μL/well). Absorbance was read at 450 nm after 10 min.
-
1) Isolation of bone marrow-derived macrophage Bone marrow-derived macrophages (BMDMs) were isolated from mouse femurs at day 10 after the last immunization. First, the bone was cut at the hip joint, the entire leg was removed, and the skin and muscle were removed very carefully. Then, the bone was cut with scissors at the knee joint. Femurs were crushed using sterilized mortar pestle in 5 mL complete Dulbecco’s Modified Eagle Media (DMEM) containing 10% FBS and 1% penicillin-streptomycin (Gibco, USA) and the cells were cultured in complete DMEM (with 10 μg/mL M-CSF) to differentiate BMDMs at 37 °C in 5% CO2. Mature macrophages could be observed under the microscope by checking the morphology of cells after approximately 10 days.
2) H37Rv infection model BMDMs (2 × 106 cells) were plated in 24-well plates (Nunc, Denmark) in complete DMEM (10% FBS, 1% penicillin-streptomycin) per well and left to adhere for 12 h. BMDMs were then infected with log phase M. tuberculosis H37Rv for 4 h (day 0) and 3 and 5 days in a BSL-3 laboratory. M. tuberculosis H37Rv infected BMDMs were lysed with 1 mL ddH2O and serially diluted (1:10, 1:100, and 1:1,000). Then, 100 µL from each preparation were inoculated on 7H10 media containing 50 µg/mL cycloheximide, 25 µg/mL polymixin B, 50 µg/mL carbenicillin, and 20 µg/mL trimethoprim and incubated at 37 °C. BMDMs isolated from adjuvant-immunized mice were used as controls. The multiplicity of infection (MOI) was set as 3 and 5. The count of intracellular M. tuberculosis was determined after 3 weeks.
-
The complete genome sequences and coding sequences (CDSs) of M. intracellulare (NC_016946) and M. tuberculosis (NC_000962) were downloaded from the National Center of Biotechnology Information (NCBI) genome website. BLASTN in the NCBI BLAST+ software package (Version 2.6.0+) was used to determine the sequence similarity (parameters were set as follows: -perc_identity 85.00, -qcov_hsp_perc 90.00, -outfmt 5, -num_threads 32, and -evalue 1e-5) between the two species. The genes that were common between the two species were classified into functional categories according to S. T. Cole’s classification[15].
-
Antigens common between the species were determined using protein microarrays (CapitalBio, China) spotted with 3791 H37Rv proteins and 428 CCDC1551 proteins. First, 3 mL serum sample diluted 1:200 with PBST was overlaid on the arrays and incubated at room temperature (RT) for 1 h. After washing three times with PBST, goat anti-mouse IgG (H+L) antibodies with Alexa-Fluor 532 or IgM with Alexa-Fluor 635 (diluted 1:1,000 with PBST) were added and incubated at RT for 1 h. Finally, arrays were washed with PBST, dried in a SlideWasher (CapitalBio, China), and scanned with GenePix 4200A (Molecular Devices, USA). Data were analyzed using GenePix 6.0 (Molecular Devices, USA). The signal-to-noise ratio (SNR) of each spot was defined as the ratio of the foreground to the background median intensity. To eliminate the systematic error between the protein arrays and different serum samples, quantile normalization was employed between the arrays. Spots with IgG-SNR > 3 or IgM-SNR > 5 were determined as positive. Gene ontology (GO) enrichment analysis and KEGG pathway analysis were performed at
http://geneontology.org/ andhttps://www.genome.jp/kegg/tool/map_pathway2.html , respectively. Protein-protein interaction analysis was performed using STRING 11.0 online (http://string-db.org/ ). Venn diagrams were generated using the R VennDiagram packagehttps://rdrr.io/bioc/limma/man/venn.html . -
All statistical analyses were performed using GraphPad Prism8 (GraphPad software, USA). The immunological data were compared using a two-tailed t-test. A two-sided P value ≤ 0.05 was used to determine statistical significance in all analyses.
-
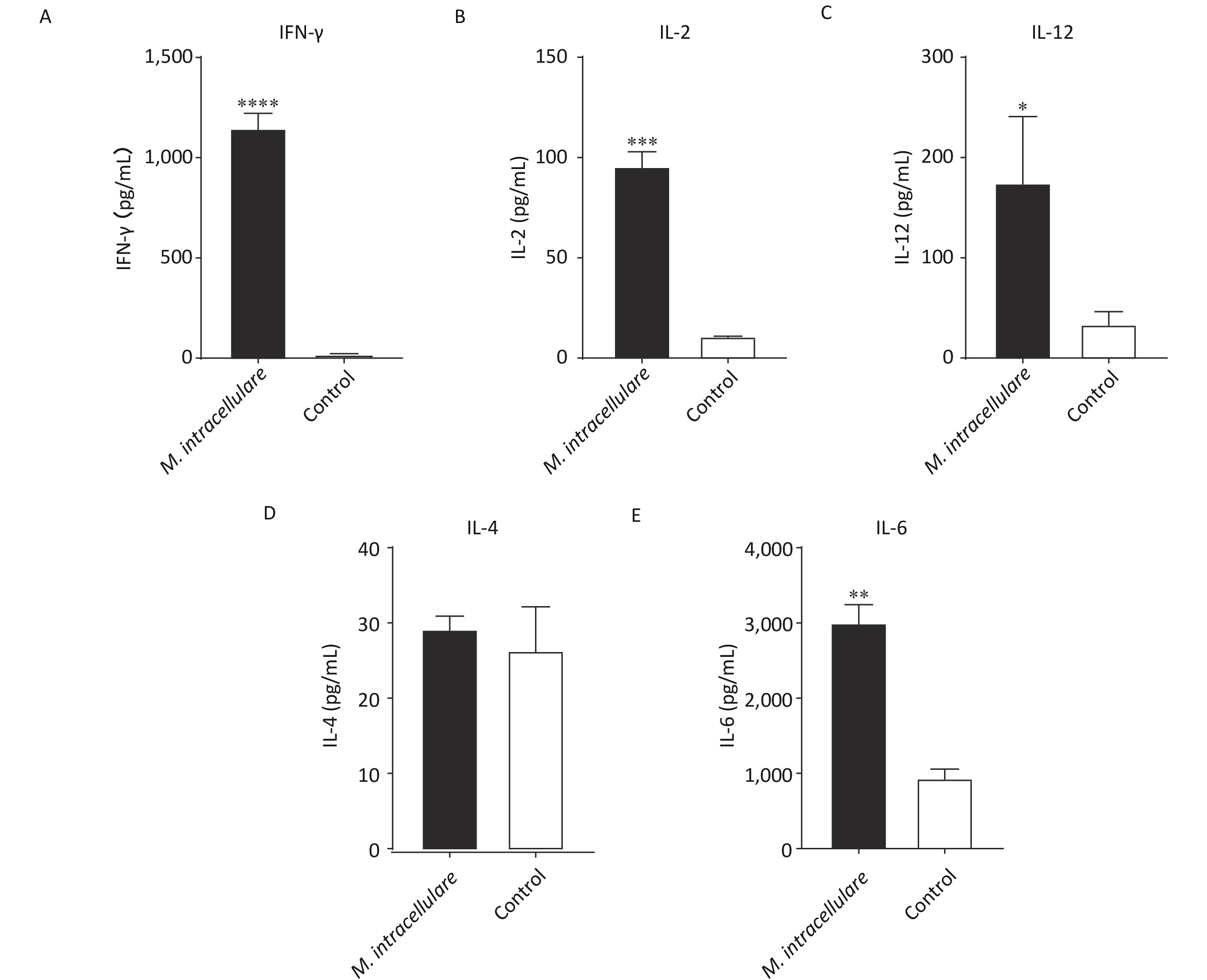
After immunizing mice three times with M. intracellulare protein extracts or adjuvants only, splenic lymphocytes were isolated and stimulated with M. intracellulare proteins. M. intracellulare protein extracts induced significantly higher levels of IFN-γ, IL-2, IL-12, and IL-6 (P < 0.0001, P < 0.001, P < 0.05, and P < 0.01, respectively), compared with control mice; however, IL-4 levels were similar in these mice (P > 0.05). Further, IL-4 concentrations obtained using both the immunization methods were low (< 30 pg/mL) (Figure 1).
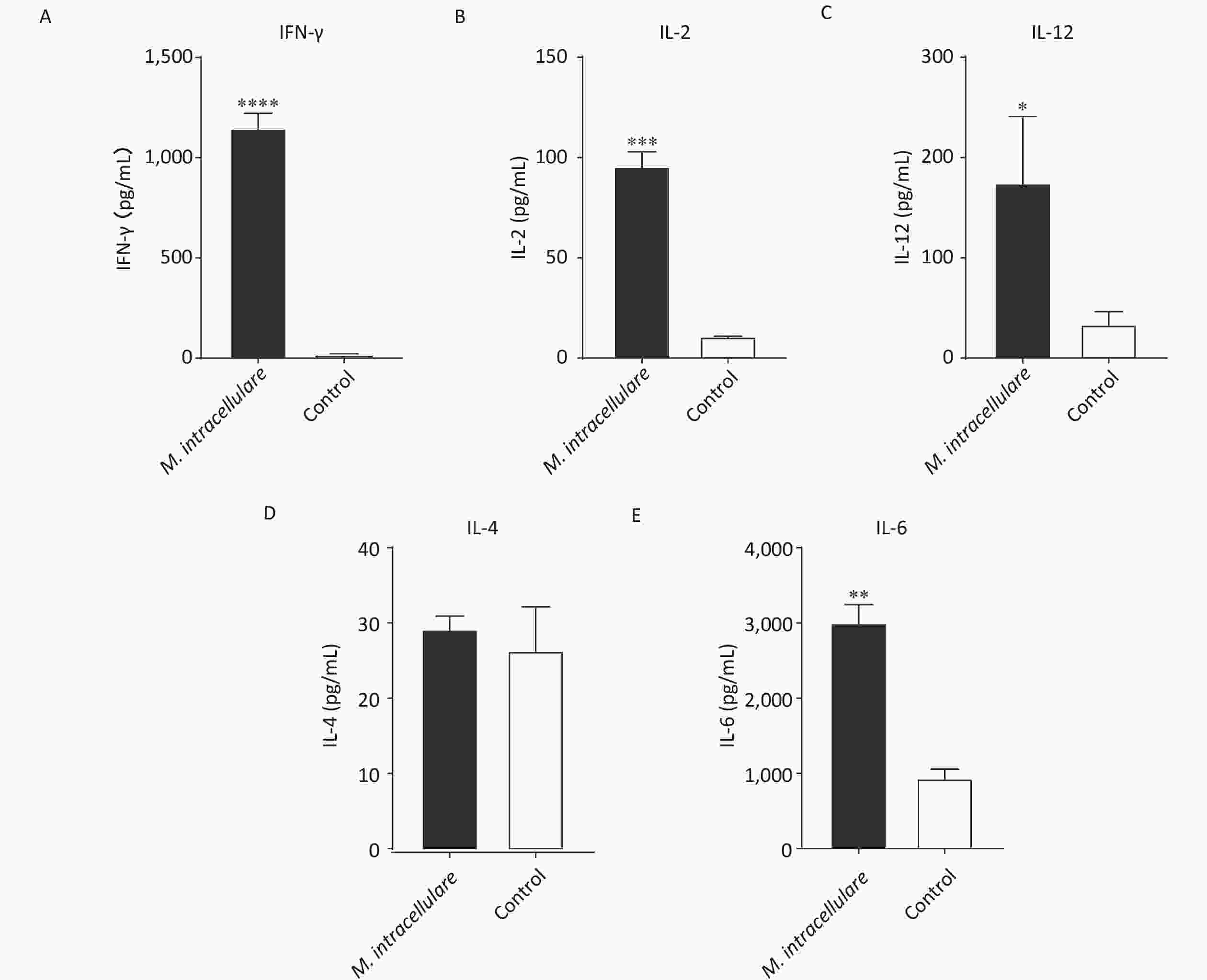
Figure 1. Cellular immune responses in immunized BALB/c mice. Lymphocytes isolated from BALB/c mice immunized with M. intracellulare protein extracts or adjuvants only were stimulated with M. intracellulare protein extracts, and the concentration of five different cytokines (A) IFN-γ, (B) IL-2, (C) IL-12, (D) IL-4, and (E) IL-6 were determined after 48 h. Capped line with asterisk * indicates significant difference between the two immunization methods, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Columns indicate the mean from at least three samples, and error bars denote the standard deviation (SD)
-
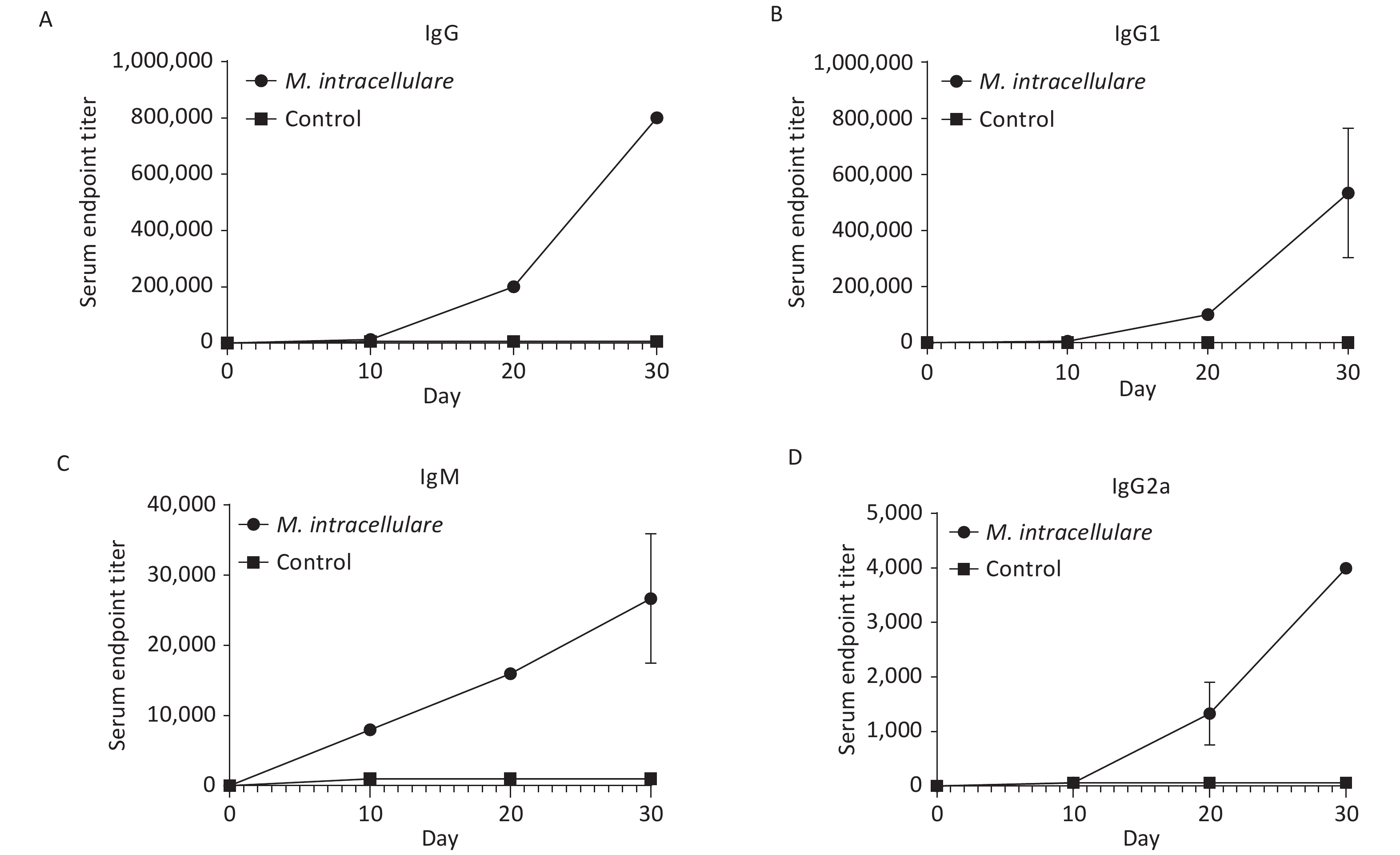
As shown in Figure 2, the prime and two prime-boost immunizations with M. intracellulare protein extracts induced strong and significant increase in IgG, IgG1, IgM, and IgG2a titers compared with pre-immunization and the adjuvant alone group at each detection point (P values were all < 0.001).
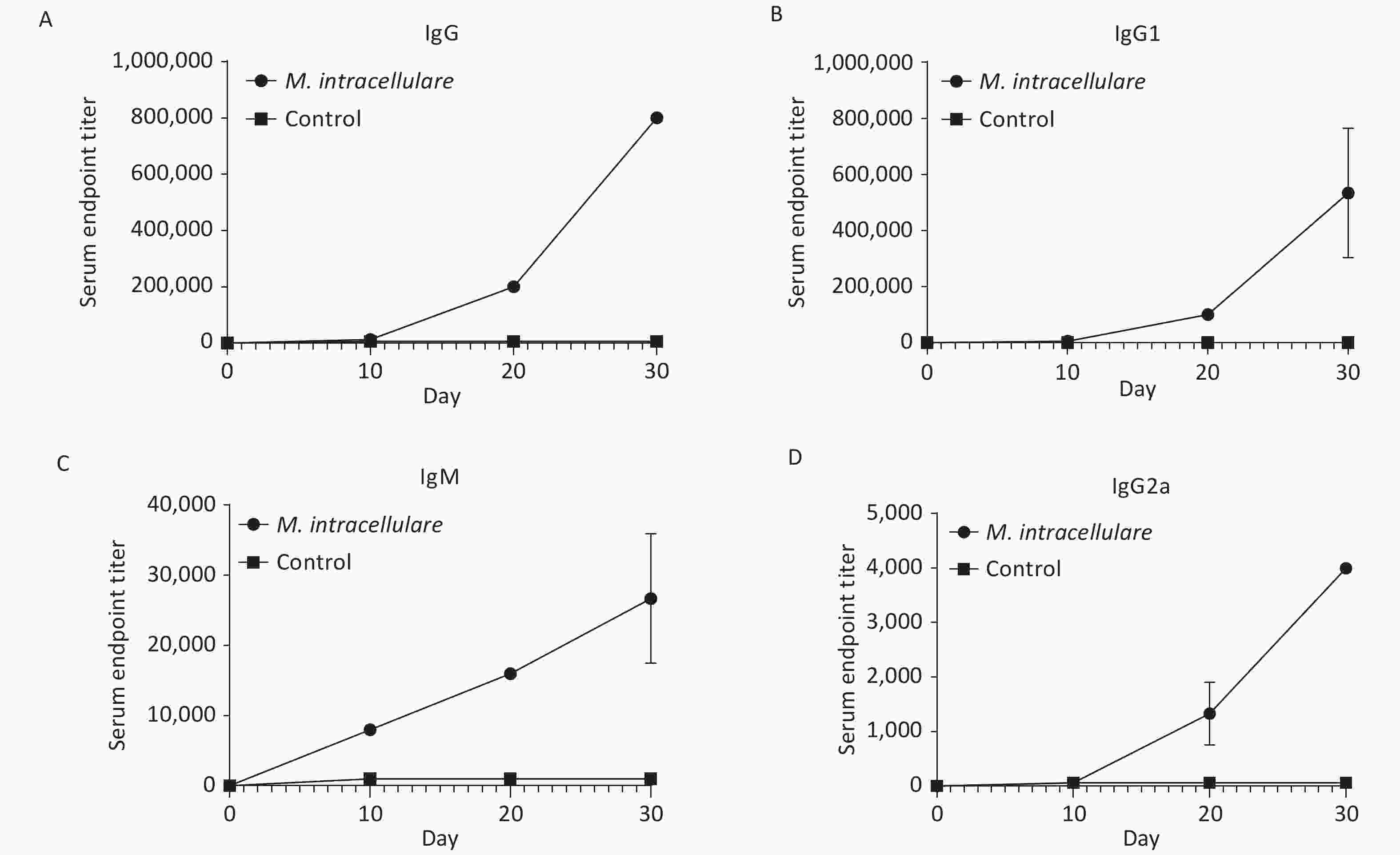
Figure 2. Antibody level monitoring in mice that had received three interval immunizations with M. intracellulare protein extracts or adjuvants. The change trends of titer levels of IgG, IgG1, IgM, and IgG2a are shown in (A), (B), (C), and (D), respectively. Dotted lines show the antibody titer levels in BALB/c mice immunized with M. intracellulare bacterial proteins + PolyI:C and DDA as an adjuvant; lines with squares show the antibody titer levels in BALB/c mice immunized with PolyI:C and DDA. The antibody titer levels were monitored at day 0, day 10 (10 days after the first immunization), day 20 (10 days after the second immunization), and day 30 (10 days after the third immunization). Points indicate the mean of at least three different serum samples in a group, and error bars denote standard deviation (SD).
By day 30, after the second boost immunization, the IgG titer reached 1:800,000 and the titer of IgG1 also reached a high level of 1:400,000, while the titer of IgM was 1:32,000 and that of IgG2a was only 1:4,000 (Figure 2). All IgG, IgG1, IgM, and IgG2a titers were significantly higher than those acquired from the prime and the first boost immunization.
-
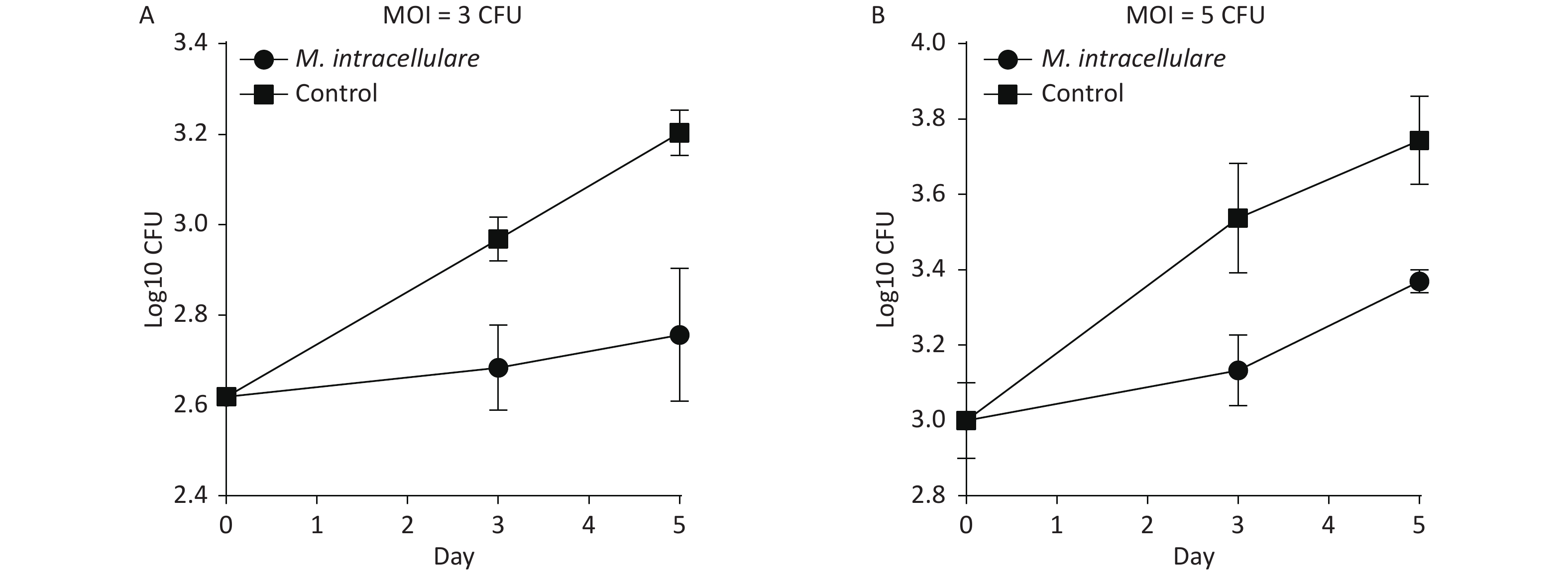
BMDMs isolated from mice immunized with M. intracellulare protein antigens were found to display significantly lower bacterial loads than those isolated from mice immunized only with adjuvants (control group) and inhibited the growth of intracellular M. tuberculosis in a dose- and time-dependent manner (Figure 3). Figure 3A shows that in MOI = 3 with 2.62 CFUs/mL (log10) H37Rv in day 0, the colony counts of the immunized group increased to 2.68 CFUs/mL (log10) when BMDMs were infected with H37Rv for 3 days, which was less than those of the control group [2.97 CFUs/mL (log10)] by 0.28 CFU/mL (log10) (P < 0.01); when BMDMs were infected with H37Rv for 5 days, the colony counts of the immunized group increased to 2.76 CFUs/mL (log10), which was significantly less than those obtained for 3.20 CFUs/mL (log10) of the control group by 0.45 CFUs/mL (log10) (P < 0.01). Figure 3B shows that in MOI = 5 with 3.01 CFUs/mL (log10) on day 0, the colony counts of the immunized group increased to 3.13 CFUs/mL (log10) when BMDMs were infected with H37Rv for 3 days, which was less than those of the control group [3.54 CFUs/mL (log10)] by 0.40 CFUs/mL (log10) (P < 0.05). When BMDMs were infected with H37Rv for 5 days, the colony counts of the immunized group increased to 3.37 CFUs/mL (log10), significantly less than those of the control group, which was 3.74 CFUs/mL (log10) by 0.3 CFUs/mL (log10) (P < 0.01).
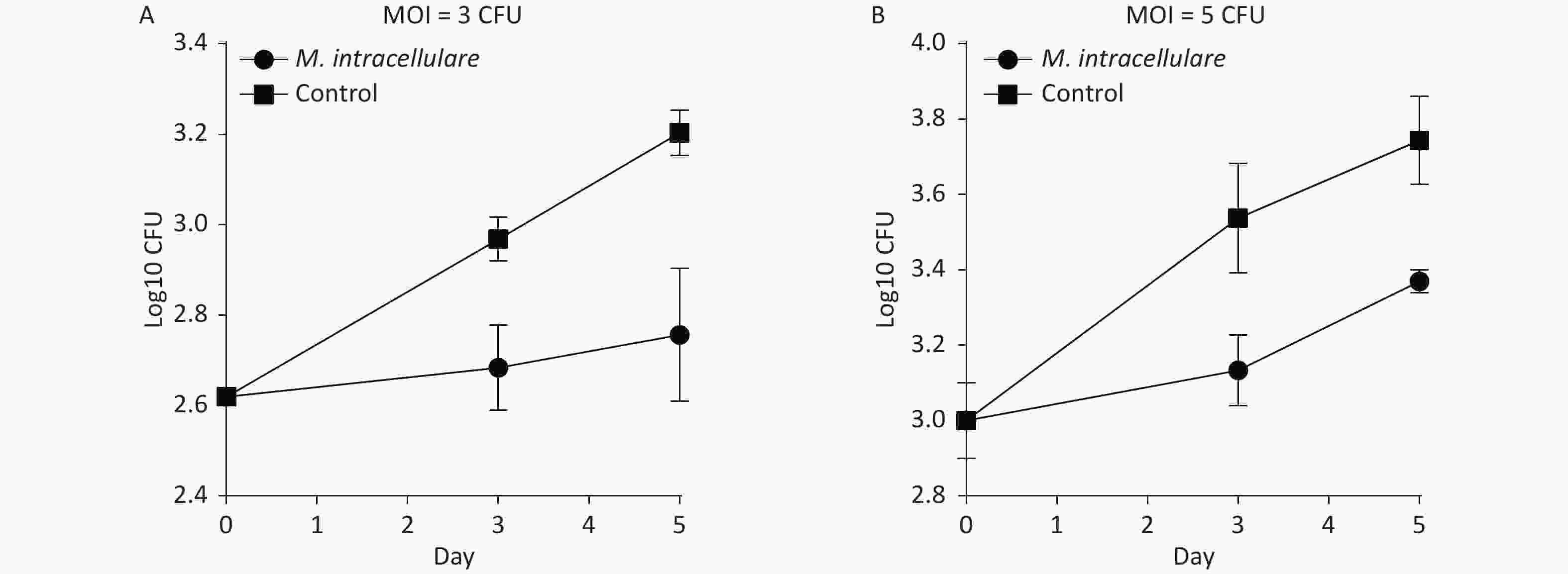
Figure 3. Immunization with M. intracellulare protein extracts enhanced the ability of bone marrow-derived macrophages to inhibit M. tuberculosis growth ex vivo. (A) BMDMs were infected with H37Rv at MOI = 3. (B) BMDMs were infected with H37Rv at MOI = 5. Colony counts were obtained for 0, 3, and 5 days of BMDM infection were compared with those obtained for H37RV. Points indicate the mean from triplicate cultures, and error bars denote standard deviation (SD). P-values were determined with two-tailed Student's t test.
-
Basic information regarding the two downloaded genomic sequences is shown in Table 1. In total, 369 common genes were found between these two genomes; the classification of these genes is shown in Table 2.
Table 1. Annotation information for the two genomic sequences
Characters NC_000962* NC_016946** Genome size (Mb) 4.41 5.4 GC (%) 65.60 68.10 No. of proteins 3,906 5,000 No. of rRNA 3 3 No. of tRNA 45 46 No. of other RNA 22 3 No. of genes 4,008 5,104 No. of pseudogenes 30 52 Note. *https://www.ncbi.nlm.nih.gov/genome/166?genome_assembly_id=159857. **https://www.ncbi.nlm.nih.gov/genome/1703?genome_assembly_id=171542. Table 2. Functional classification of the 369 common genes found in the genomes of M. tuberculosis and M. intracellulare
No. Function Number 1 Virulence, detoxification, adaptation 10 2 Lipid metabolism 35 3 Information pathways 73 4 Cell wall and cell processes 39 5 Intermediary metabolism and respiration 143 6 Regulatory proteins 24 7 Conserved hypotheticals 41 -
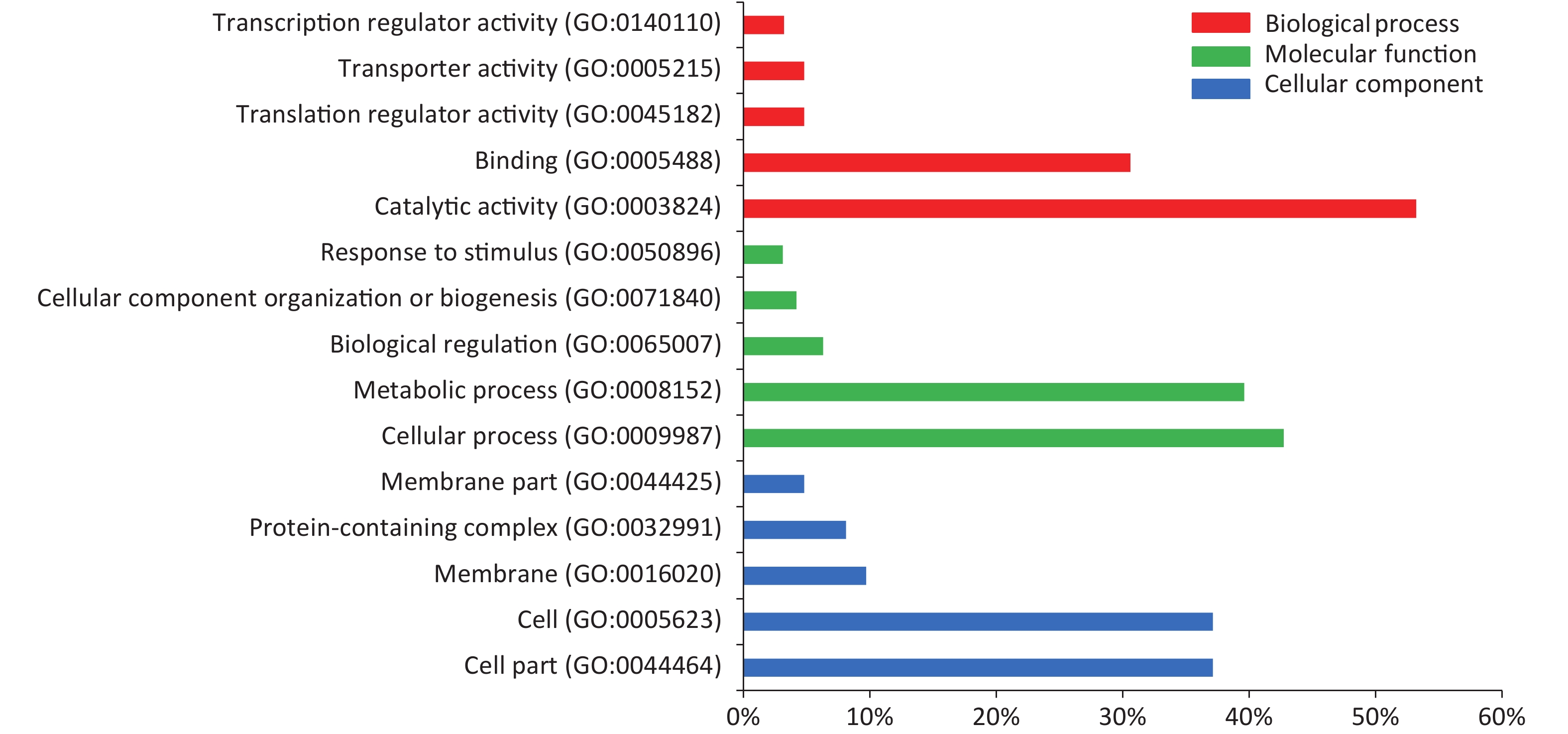
In the serum of M. intracellulare-immunized mice, 478 proteins were recognized by IgG and IgM antibodies in the protein microarray. GO enrichment analysis based on the Gene Ontology database revealed the top 15 GO terms to be in three categories according to the GO classification, of which “binding” “catalytic activity” “metabolic process” “cellular process” “cell” “cell part” were dominant, as shown in Figure 4.
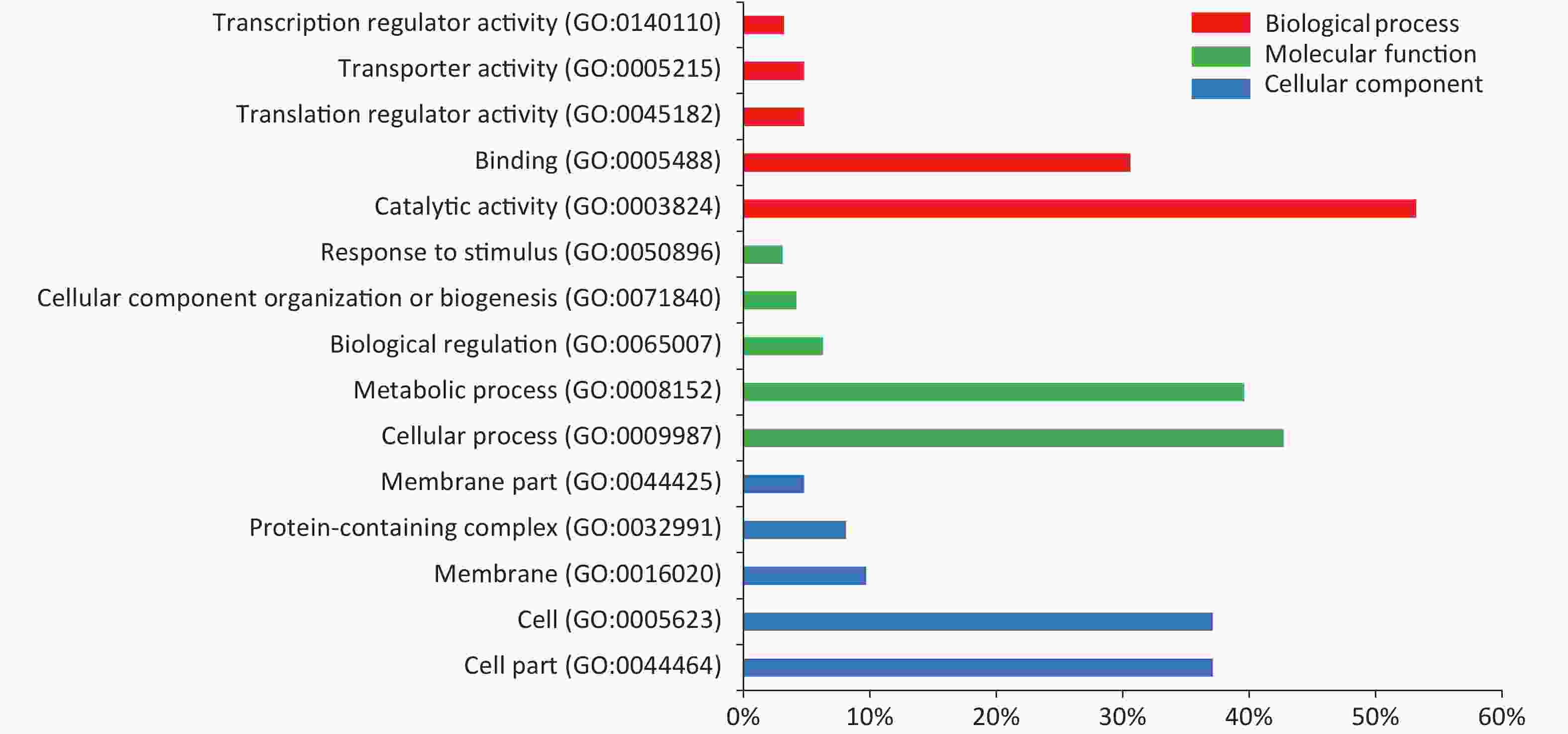
Figure 4. Top 15 GO terms identified based on the proteins found in the serum of M. intracellulare immunized mice. The results include three main categories: red bars represent biological processes, green bars represent molecular functions, and blue bars represent cellular components. The x-axis shows the percentage of the specific GO term in each category. GO, gene ontology.
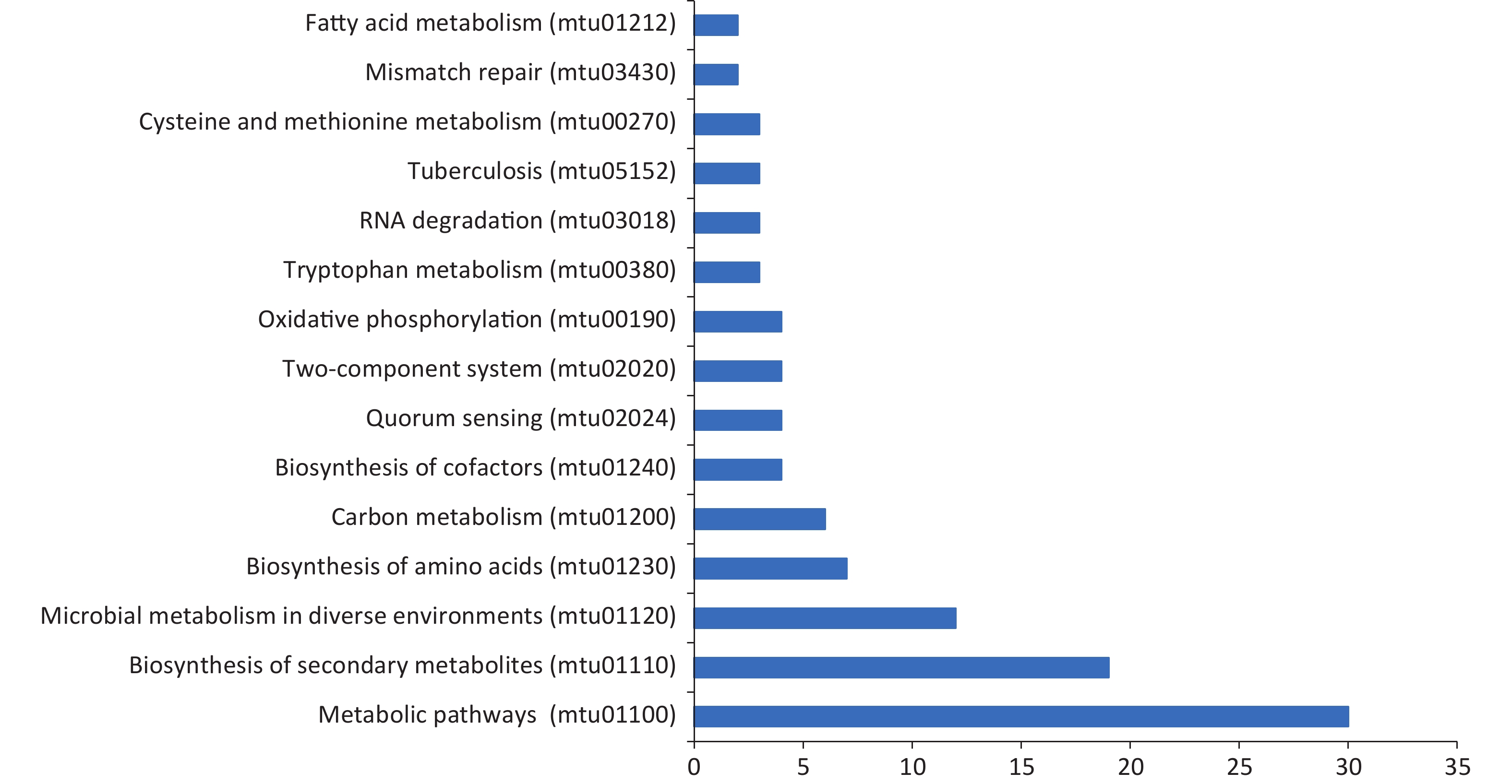
We also conducted KEGG pathway analysis on these microarray-screened proteins. The names, functions, and pathways of 478 proteins are listed in Supplementary Table S1, available in www.besjournal.com. The top 15 enrichment pathways classified based on the screened proteins are displayed in Figure 5. Of the enriched pathways, “metabolic pathways” was the predominant pathway with 30 proteins, followed by “biosynthesis of secondary metabolites” with 19 proteins and “microbial metabolism in diverse environments” with 12 proteins.
Table S1. Names, functions, and pathways of 478 proteins identified in the sera of M. intracellulare-immunized mice by protein microarrays
ID KEGG pathway No. of genes Genes mapped mtu01100 Metabolic
pathways30 mtu:Rv0091 mtn; 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase mtu:Rv0500 proC; pyrroline-5-carboxylate reductase mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0769 oxidoreductase mtu:Rv0848 cysK2; cysteine synthase CysK mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv0956 purN; phosphoribosylglycinamide formyltransferase PurN mtu:Rv1092c coaA; pantothenate kinase mtu:Rv1164 narI; nitrate reductase subunit gamma mtu:Rv1257c oxidoreductase mtu:Rv1263 amiB2; amidase AmiB mtu:Rv1306 atpF; ATP synthase subunit B mtu:Rv1308 atpA; ATP synthase subunit alpha mtu:Rv1448c tal; transaldolase mtu:Rv1530 adh; alcohol dehydrogenase mtu:Rv1604 impA; inositol-monophosphatase ImpA mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2195 qcrA; ubiquinol-cytochrome C reductase rieske iron-sulfur subunit mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2350c plcB; membrane-associated phospholipase B mtu:Rv2377c hypothetical protein mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3009c gatB; aspartyl/glutamyl-tRNA(Asn/Gln) amidotransferase subunit B mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu:Rv3106 fprA; NADPH-ferredoxin reductase FprA mtu:Rv3465 rmlC; dTDP-4-dehydrorhamnose 3,5-epimerase mtu:Rv3600c type III pantothenate kinase mtu:Rv3703c etgB; iron(II)-dependent oxidoreductase EgtB mtu01110 Biosynthesis of
secondary
metabolites19 mtu:Rv0500 proC; pyrroline-5-carboxylate reductase mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0769 oxidoreductase mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv0956 purN; phosphoribosylglycinamide formyltransferase PurN mtu:Rv1086 (2Z,6E)-farnesyl diphosphate synthase mtu:Rv1257c oxidoreductase mtu:Rv1448c tal; transaldolase mtu:Rv1530 adh; alcohol dehydrogenase mtu:Rv1604 impA; inositol-monophosphatase ImpA mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2350c plcB; membrane-associated phospholipase B mtu:Rv2377c hypothetical protein mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu:Rv3465 rmlC; dTDP-4-dehydrorhamnose 3,5-epimerase mtu01120 Microbial
metabolism in
diverse
environments12 mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv1164 narI; nitrate reductase subunit gamma mtu:Rv1257c oxidoreductase mtu:Rv1263 amiB2; amidase AmiB mtu:Rv1448c tal; transaldolase mtu:Rv1530 adh; alcohol dehydrogenase mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu01230 Biosynthesis of
amino acids7 mtu:Rv0091 mtn; 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase mtu:Rv0500 proC; pyrroline-5-carboxylate reductase mtu:Rv1448c tal; transaldolase mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu01200 Carbon
metabolism6 mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv1448c tal; transaldolase mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu01240 Biosynthesis of
cofactors4 mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0769 oxidoreductase mtu:Rv1092c coaA; pantothenate kinase mtu:Rv3600c type III pantothenate kinase mtu02024 Quorum sensing 4 mtu:Rv0732 secY; preprotein translocase SecY mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2350c plcB; membrane-associated phospholipase B mtu:Rv3676 crp; cAMP receptor protein mtu00190 Oxidative
phosphorylation4 mtu:Rv1306 atpF; ATP synthase subunit B mtu:Rv1308 atpA; ATP synthase subunit alpha mtu:Rv2195 qcrA; ubiquinol-cytochrome C reductase rieske iron-sulfur subunit mtu:Rv3628 ppa; inorganic pyrophosphatase mtu02020 Two-component
system4 mtu:Rv1164 narI; nitrate reductase subunit gamma mtu:Rv2234 ptpA; protein-tyrosine-phosphatase mtu:Rv3132c devS; two component sensor histidine kinase DevS mtu:Rv3676 crp; cAMP receptor protein mtu00270 Cysteine and
methionine
metabolism3 mtu:Rv0091 mtn; 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase mtu:Rv0848 cysK2; cysteine synthase CysK mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu00380 Tryptophan
metabolism3 mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv1263 amiB2; amidase AmiB mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu03018 RNA
degradation3 mtu:Rv0350 dnaK; chaperone protein DnaK mtu:Rv0440 groEL2; molecular chaperone GroEL mtu:Rv3417c groEL1; chaperonin GroEL mtu05152 Tuberculosis 3 mtu:Rv0350 dnaK; chaperone protein DnaK mtu:Rv0440 groEL2; molecular chaperone GroEL mtu:Rv3417c groEL1; chaperonin GroEL mtu03430 Mismatch repair 2 mtu:Rv0002 dnaN; DNA polymerase III subunit beta mtu:Rv2413c hypothetical protein mtu01212 Fatty acid
metabolism2 mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0769 oxidoreductase 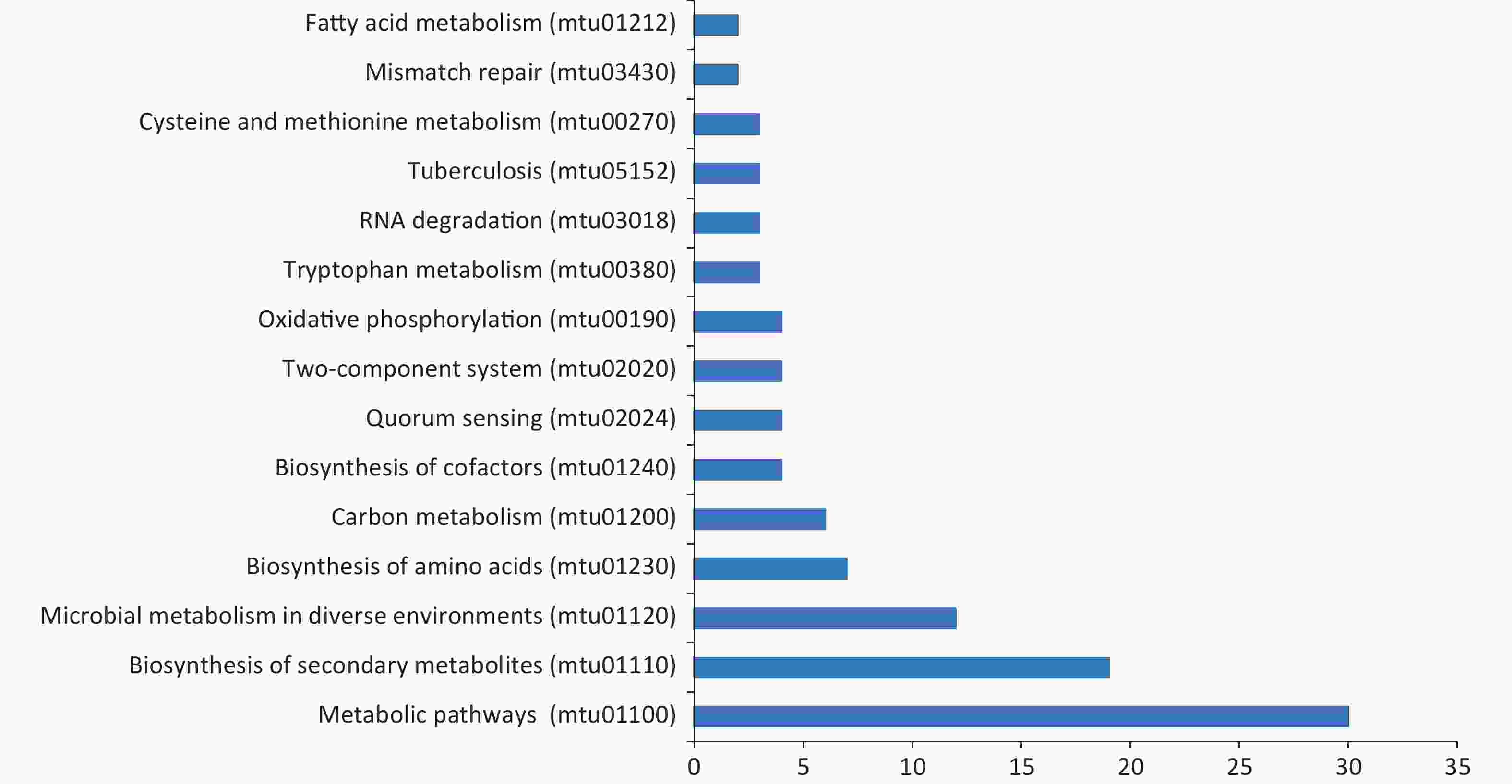
Figure 5. Kyoto encyclopedia of genes and genomes (KEGG) pathway analysis for screened antigens. Blue bars represent each category with the name labeled in the left; x-axis shows the number of antigens in each specific KEGG term in each category.
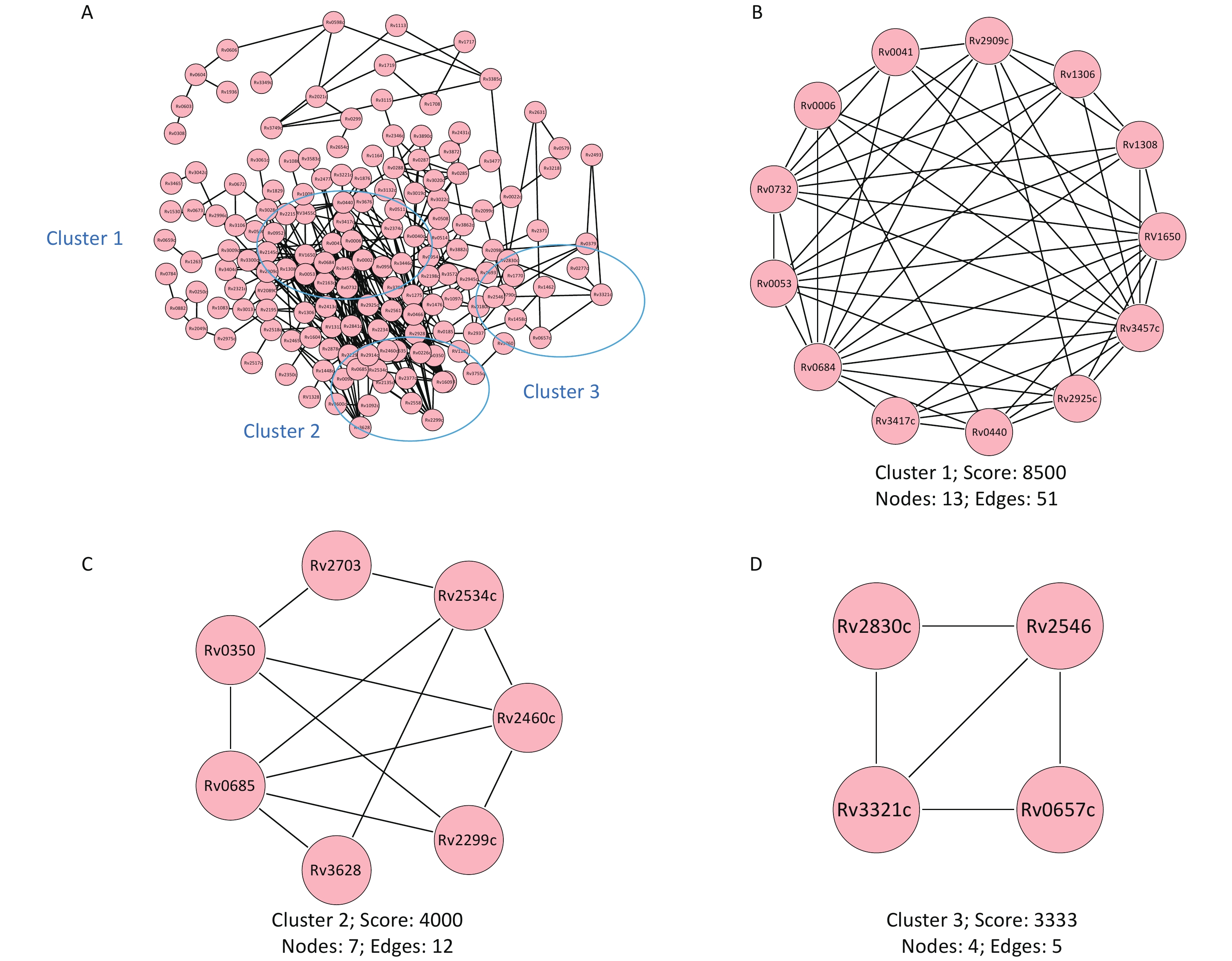
The results from the protein-protein interaction analysis are shown in Figure 6. Of the proteins recognized by the microchip, three main interaction network clusters were found. The cluster with the highest score contained 13 proteins (Rv1308, Rv2925c, Rv2909c, Rv0041, Rv1306, Rv0732, Rv0006, Rv0440, Rv0053, Rv0684, Rv3417c, Rv3457c, Rv1650) and belonged to the cell composition cluster (GO:0044464). The second cluster contained seven proteins (Rv2460c, Rv2299c, Rv0350, Rv2703, Rv2534c, Rv3628, Rv0685) and belonged to the cytoplasmic composition modification (GO:0005737) related cluster. The third cluster contained four proteins (Rv2830c, Rv2546, Rv3321c, Rv0657c) and belonged to the toxin-antitoxin system (kw-1277) related cluster.
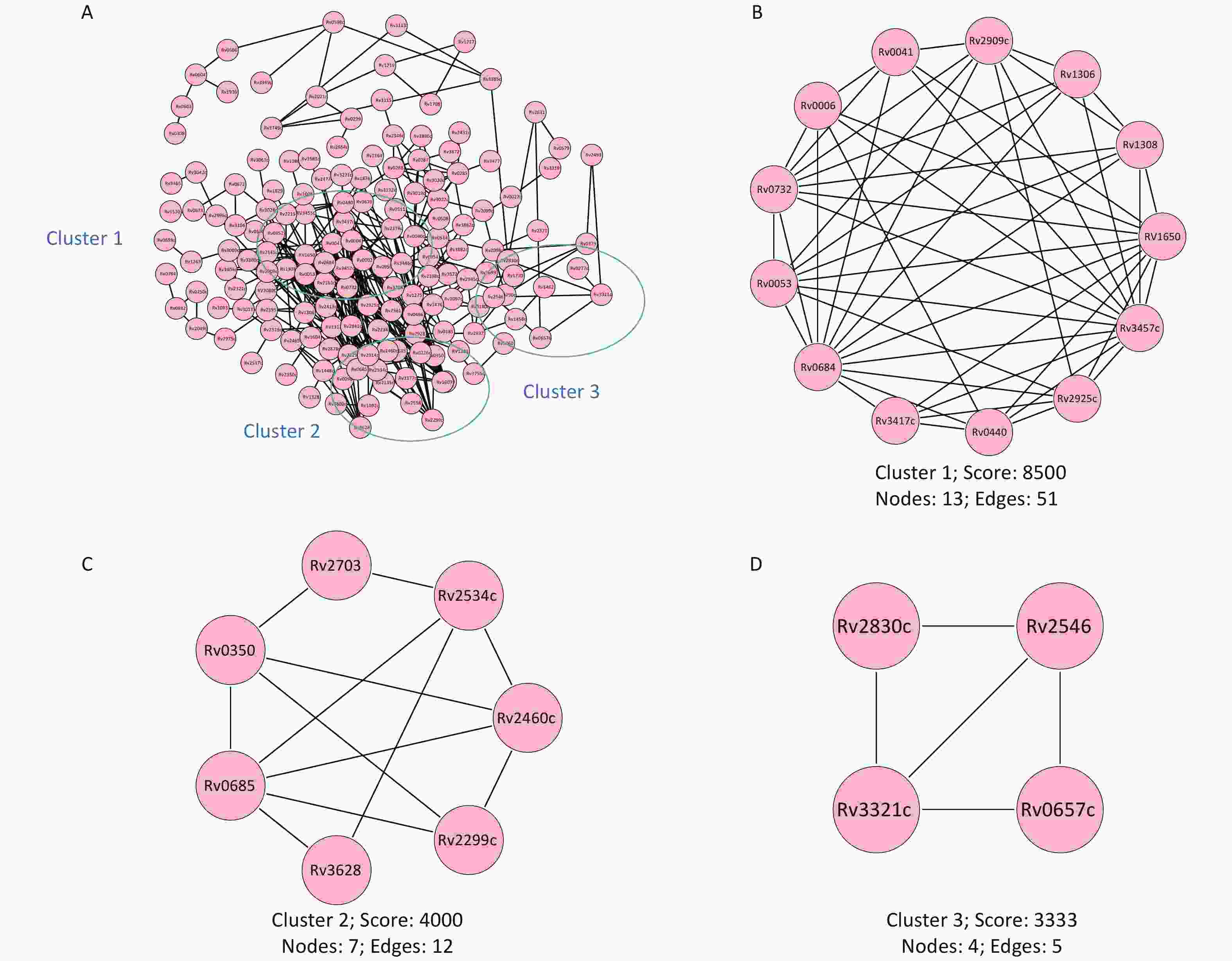
Figure 6. Protein-protein interaction network. The protein interaction network was constructed based on the String database. (A) Map of the interaction network of all screened antigenic proteins. (B) Cell composition (GO:0044464) correlated cluster. (C) Cytoplasmic composition modification (GO:0005737) correlated cluster. (D) Toxin-antitoxin system (kw-1277) correlated cluster. Lines represent specific protein-protein associations. GO, gene ontology.
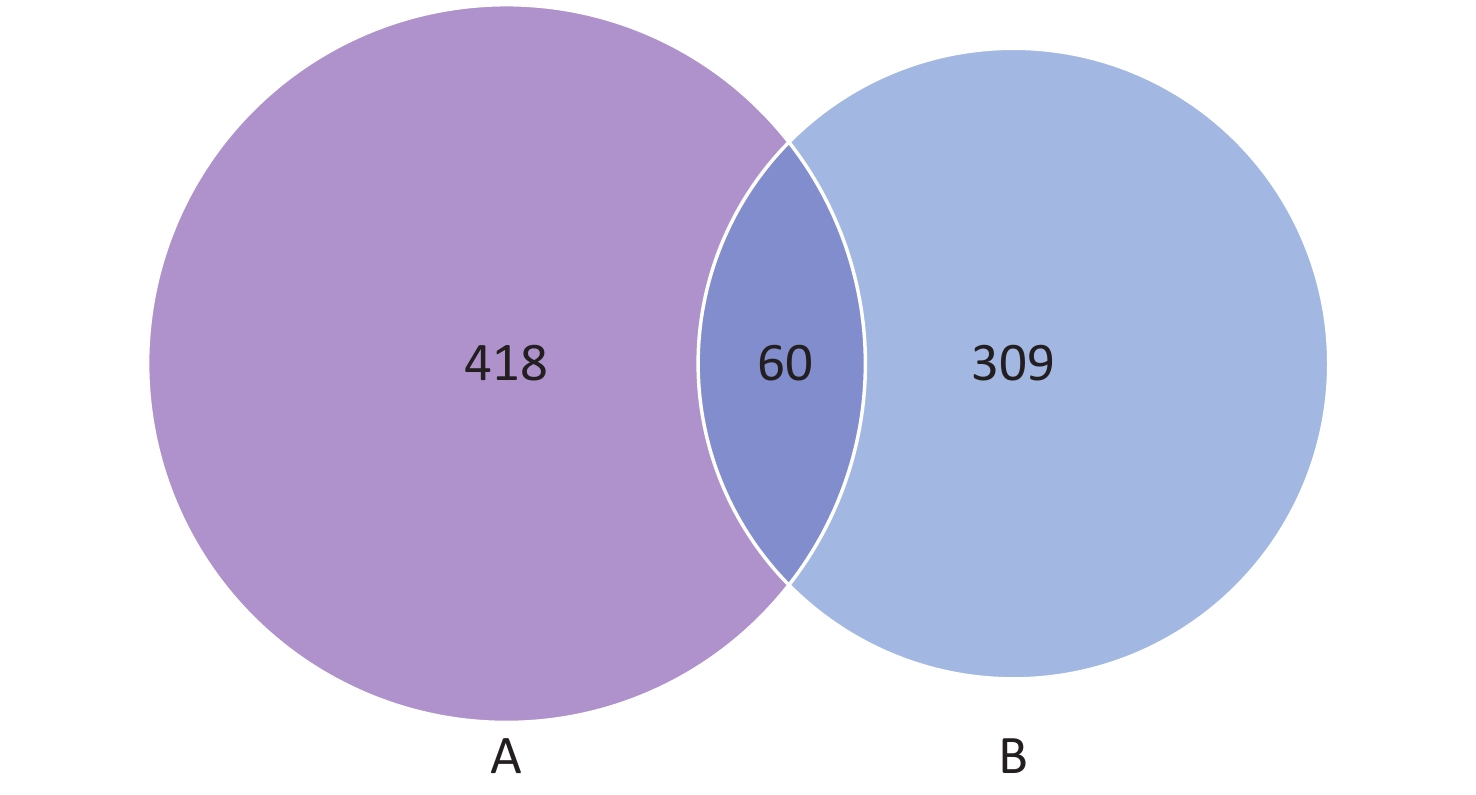
The results from the comparative analysis of antigens in protein microarray and comparative genomic analysis are shown in Figure 7. Of the 478 genes and 369 proteins found by microchip and comparative genomic analysis, 60 shared common antigens were found. In total, 418 exclusive antigens were revealed from protein microarray analysis and 309 from comparative genomic analysis. The names of the proteins in each group are listed in Supplementary Table S2, available in www.besjournal.com.
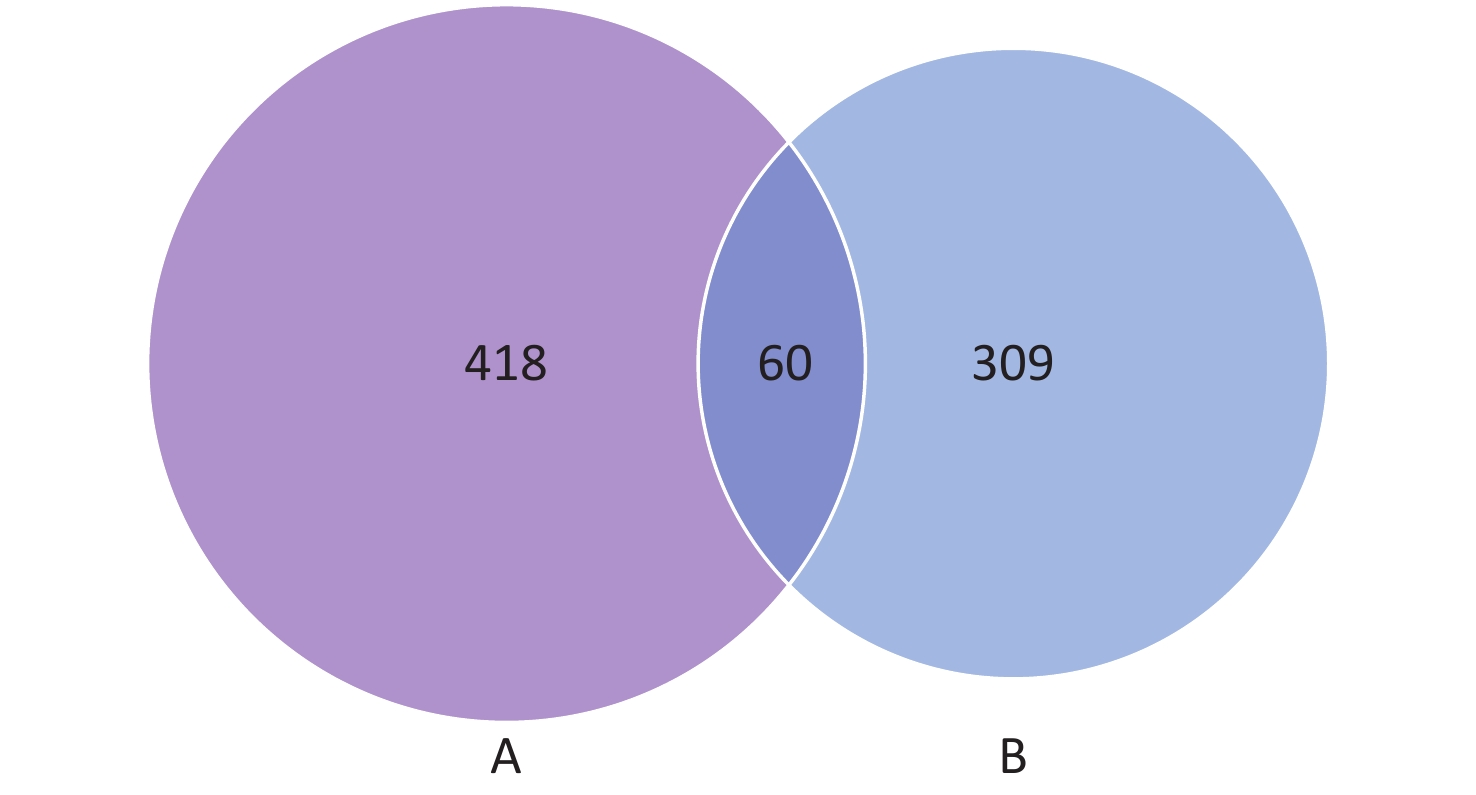
Figure 7. Comparative analysis of antigens in protein microarray and comparative genomic analysis. Venn diagram showing the distribution of common antigens and exclusive antigens between protein microarrays (A) and comparative genomic analysis (B). The numbers shown in the diagram represent the number of proteins in each group.
Table S2. Names of common antigens identified by microchip and comparative genomic analysis
No. Genome BioChip Common 1 Rv0009 MT0066.1 Rv0009 2 Rv0011c MT0066.2 Rv0046c 3 Rv0019c MT0116.1 Rv0053 4 Rv0046c MT0250 Rv0337c 5 Rv0053 MT0270.1 Rv0350 6 Rv0055 MT0383 Rv0384c 7 Rv0126 MT0392 Rv0423c 8 Rv0130 MT0407 Rv0440 9 Rv0137c MT0470 Rv0491 10 Rv0156 MT0487 Rv0551c 11 Rv0157 MT0543 Rv0667 12 Rv0158 MT0555 Rv0672 13 Rv0189c MT0610 Rv0673 14 Rv0211 MT0946 Rv0684 15 Rv0230c MT0968.1 Rv0685 16 Rv0236A MT1029 Rv0732 17 Rv0238 MT1040.1 Rv0861c 18 Rv0242c MT1055 Rv0984 19 Rv0244c MT1083.1 Rv1013 20 Rv0267 MT1172.1 Rv1070c 21 Rv0321 MT1264.1 Rv1080c 22 Rv0337c MT1305.1 Rv1086 23 Rv0350 MT1329 Rv1092c 24 Rv0352 MT1479.1 Rv1257c 25 Rv0357c MT1534 Rv1292 26 Rv0363c MT1555.1 Rv1308 27 Rv0380c MT1775 Rv1458c 28 Rv0384c MT1849.1 Rv1654 29 Rv0391 MT2068 Rv1829 30 Rv0407 MT2113 Rv2115c 31 Rv0411c MT2138.2 Rv2241 32 Rv0423c MT2142 Rv2346c 33 Rv0430 MT2283 Rv2374c 34 Rv0440 MT2291 Rv2457c 35 Rv0458 MT2316 Rv2460c 36 Rv0465c MT2330.1 Rv2465c 37 Rv0467 MT2334.1 Rv2477c 38 Rv0491 MT2361.1 Rv2534c 39 Rv0498 MT2405 Rv2558 40 Rv0500A MT2455 Rv2697c 41 Rv0510 MT2488.1 Rv2711 42 Rv0527 MT2501 Rv2754c 43 Rv0548c MT2502 Rv2788 44 Rv0551c MT2520.1 Rv2795c 45 Rv0566c MT2554.1 Rv2909c 46 Rv0634B MT2625 Rv3009c 47 Rv0636 MT2626 Rv3028c 48 Rv0639 MT2637.1 Rv3118 49 Rv0640 MT2721 Rv3221c 50 Rv0641 MT2779 Rv3248c 51 Rv0642c MT2871 Rv3412 52 Rv0647c MT2958.1 Rv3442c 53 Rv0651 MT3139.1 Rv3457c 54 Rv0652 MT3270.1 Rv3551 55 Rv0655 MT3279 Rv3583c 56 Rv0667 MT3284 Rv3609c 57 Rv0668 MT3289 Rv3628 58 Rv0672 MT3290.2 Rv3676 59 Rv0673 MT3532.1 Rv3710 60 Rv0682 MT3573.12 Rv3791 61 Rv0683 MT3631 62 Rv0684 MT3770 63 Rv0685 MT3858 64 Rv0691A MT3876 65 Rv0693 MT3878 66 Rv0700 MT4026.1 67 Rv0701 Rv0002 68 Rv0702 Rv0006 69 Rv0703 Rv0009 70 Rv0704 Rv0022c 71 Rv0705 Rv0025 72 Rv0707 Rv0028 73 Rv0708 rv0036 74 Rv0709 Rv0040c 75 Rv0714 Rv0041 76 Rv0716 Rv0043c 77 Rv0717 Rv0045c 78 Rv0718 Rv0046c 79 Rv0719 Rv0053 80 Rv0721 Rv0063 81 Rv0723 Rv0076c 82 Rv0732 Rv0089 83 Rv0733 Rv0091 84 Rv0737 Rv0095c 85 Rv0753c Rv0098 86 Rv0803 Rv0100 87 Rv0808 Rv0110 88 Rv0814c Rv0119 89 Rv0815c Rv0145 90 Rv0820 Rv0150c 91 Rv0821c Rv0155 92 Rv0859 Rv0180c 93 Rv0861c Rv0185 94 Rv0889c Rv0187 95 Rv0896 Rv0226c 96 Rv0903c Rv0232 97 Rv0946c Rv0248c 98 Rv0958 Rv0250c 99 Rv0974c Rv0264c 100 Rv0975c Rv0277c 101 Rv0981 Rv0281 102 Rv0984 Rv0285 103 Rv1013 Rv0287 104 Rv1017c Rv0288 105 Rv1019 Rv0290 106 Rv1023 Rv0295c 107 Rv1038c Rv0299 108 Rv1070c Rv0301 109 Rv1074c Rv0308 110 Rv1077 Rv0333 111 Rv1080c Rv0337c 112 Rv1086 Rv0350 113 Rv1092c Rv0369c 114 Rv1095 Rv0379 115 Rv1098c Rv0384c 116 Rv1099c Rv0385 117 Rv1151c Rv0387c 118 Rv1177 Rv0398c 119 Rv1187 Rv0423c 120 Rv1197 Rv0429c 121 Rv1198 Rv0437c 122 Rv1211 Rv0440 123 Rv1213 Rv0446c 124 Rv1240 Rv0459 125 Rv1248c Rv0466 126 Rv1257c Rv0489 127 Rv1262c Rv0491 128 Rv1292 Rv0500 129 Rv1298 Rv0508 130 Rv1305 Rv0511 131 Rv1308 Rv0514 132 Rv1310 Rv0518 133 Rv1311 Rv0521c 134 Rv1315 Rv0546c 135 Rv1321 Rv0551c 136 Rv1331 Rv0561c 137 Rv1380 Rv0571c 138 Rv1381 Rv0577 139 Rv1383 Rv0579 140 Rv1384 Rv0580c 141 Rv1388 Rv0598c 142 Rv1392 Rv0600c 143 Rv1415 Rv0603 144 Rv1423 Rv0604 145 Rv1436 Rv0606 146 Rv1447c Rv0612 147 Rv1458c Rv0635 148 Rv1474c Rv0657c 149 Rv1475c Rv0659c 150 Rv1479 Rv0666 151 Rv1481 Rv0667 152 Rv1483 Rv0672 153 Rv1484 Rv0673 154 Rv1488 Rv0678 155 Rv1493 Rv0684 156 Rv1547 Rv0685 157 Rv1589 Rv0699 158 Rv1601 Rv0730 159 Rv1611 Rv0731c 160 Rv1617 Rv0732 161 Rv1627c Rv0750 162 Rv1630 Rv0764c 163 Rv1633 Rv0766c 164 Rv1638 Rv0769 165 Rv1641 Rv0772 166 Rv1642 Rv0784 167 Rv1643 Rv0790c 168 Rv1654 Rv0793 169 Rv1655 Rv0801 170 Rv1657 Rv0810c 171 Rv1658 Rv0819 172 Rv1659 Rv0828c 173 Rv1729c Rv0837c 174 Rv1730c Rv0848 175 Rv1731 Rv0857 176 Rv1783 Rv0861c 177 Rv1793 Rv0865 178 Rv1821 Rv0869c 179 Rv1827 Rv0882 180 Rv1829 Rv0887c 181 Rv2050 Rv0937c 182 Rv2062c Rv0950c 183 Rv2097c Rv0952 184 Rv2111c Rv0954 185 Rv2112c Rv0956 186 Rv2115c Rv0970 187 Rv2122c Rv0984 188 Rv2134c Rv1008 189 Rv2146c Rv1012 190 Rv2150c RV1013 191 Rv2156c Rv1046c 192 Rv2166c Rv1056 193 Rv2178c Rv1070c 194 Rv2193 Rv1080c 195 Rv2204c Rv1083 196 Rv2218 Rv1086 197 Rv2220 Rv1092c 198 Rv2222c Rv1097c 199 Rv2225 Rv1109c 200 Rv2241 Rv1112 201 Rv2244 Rv1113 202 Rv2245 Rv1124 203 Rv2247 Rv1132 204 Rv2256c Rv1153c 205 Rv2259 Rv1164 206 Rv2346c RV1181 207 Rv2347c Rv1202 208 Rv2362c Rv1208 209 Rv2367c Rv1239c 210 Rv2374c Rv1257c 211 Rv2375 RV1259 212 Rv2402 Rv1263 213 Rv2404c RV1264 214 Rv2406c Rv1275 215 Rv2412 RV1282C 216 Rv2420c Rv1284 217 Rv2421c Rv1292 218 Rv2426c Rv1306 219 Rv2428 Rv1308 220 Rv2441c Rv1309 221 Rv2442c RV1312 222 Rv2448c RV1328 223 Rv2457c Rv1371 224 Rv2460c Rv1373 225 Rv2461c Rv1377c 226 Rv2465c RV1395 227 Rv2466c RV1404 228 Rv2477c Rv1414 229 Rv2502c Rv1427c 230 Rv2511 Rv1428c 231 Rv2534c Rv1448c 232 Rv2539c Rv1451 233 Rv2540c Rv1458c 234 Rv2558 Rv1462 235 Rv2572c Rv1463 236 Rv2583c Rv1476 237 Rv2592c Rv1501 238 Rv2603c Rv1530 239 Rv2605c Rv1531 240 Rv2606c Rv1536 241 Rv2674 Rv1544 242 Rv2676c Rv1584c 243 Rv2692 Rv1604 244 Rv2697c Rv1609 245 Rv2699c RV1650 246 Rv2708c Rv1654 247 Rv2710 Rv1677 248 Rv2711 Rv1685c 249 Rv2713 Rv1692 250 Rv2720 Rv1693 251 Rv2725c Rv1695 252 Rv2733c Rv1708 253 Rv2744c Rv1710 254 Rv2754c Rv1717 255 Rv2764c RV1718 256 Rv2783c Rv1719 257 Rv2788 Rv1724c 258 Rv2795c Rv1742 259 Rv2831 Rv1770 260 Rv2840c Rv1791 261 Rv2861c Rv1806 262 Rv2868c Rv1828 263 Rv2882c Rv1829 264 Rv2890c RV1837c 265 Rv2901c Rv1848 266 Rv2904c Rv1875 267 Rv2909c Rv1876 268 Rv2911 Rv1893 269 Rv2919c Rv1894c 270 Rv2927c Rv1896c 271 Rv2965c Rv1898 272 Rv2975a RV1912C 273 Rv2987c Rv1936 274 Rv3003c Rv1959c 275 Rv3009c Rv1968 276 Rv3011c Rv1988 277 Rv3012c Rv1992c 278 Rv3028c Rv2021c 279 Rv3029c Rv2042c 280 Rv3043c Rv2043c 281 Rv3048c Rv2049c 282 Rv3051c RV2089C 283 Rv3053c Rv2098c 284 Rv3102c Rv2099c 285 Rv3105c Rv2102 286 Rv3117 Rv2104c 287 Rv3118 Rv2107 288 Rv3146 Rv2115c 289 Rv3148 Rv2130c 290 Rv3150 Rv2135c 291 Rv3155 Rv2140c 292 Rv3219 Rv2145c 293 Rv3221c Rv2158c 294 Rv3240c Rv2163c 295 Rv3246c Rv2195 296 Rv3248c Rv2198c 297 Rv3270 Rv2215 298 Rv3280 Rv2226 299 Rv3303c Rv2229c 300 Rv3318 Rv2233 301 Rv3319 Rv2234 302 Rv3339c Rv2239c 303 Rv3340 Rv2241 304 Rv3356c Rv2258c 305 Rv3362c Rv2293c 306 Rv3368c Rv2299c 307 Rv3370c Rv2321c 308 Rv3396c Rv2324 309 Rv3409c Rv2346c 310 Rv3410c Rv2350c 311 Rv3411c Rv2351c 312 Rv3412 Rv2360c 313 Rv3418c Rv2368c 314 Rv3432c Rv2371 315 Rv3436c Rv2374c 316 Rv3442c Rv2376c 317 Rv3443c Rv2377c 318 Rv3457c Rv2413c 319 Rv3458c Rv2431c 320 Rv3459c Rv2436 321 Rv3460c RV2438C 322 Rv3461c Rv2445c 323 Rv3462c Rv2452c 324 Rv3464 Rv2457c 325 Rv3489 Rv2460c 326 Rv3501c Rv2465c 327 Rv3515c RV2467 328 Rv3516 Rv2471 329 Rv3526 Rv2473 330 Rv3534c Rv2477c 331 Rv3535c Rv2493 332 Rv3543c Rv2499c 333 Rv3550 Rv2505c 334 Rv3551 Rv2517c 335 Rv3553 Rv2518c 336 Rv3556c Rv2528c 337 Rv3557c Rv2534c 338 Rv3559c Rv2546 339 Rv3560c Rv2555c 340 Rv3562 Rv2558 341 Rv3567c Rv2561 342 Rv3568c Rv2564 343 Rv3570c Rv2576c 344 Rv3574 Rv2579 345 Rv3583c Rv2595 346 Rv3586 Rv2614A 347 Rv3592 Rv2631 348 Rv3596c Rv2638 349 Rv3597c Rv2641 350 Rv3609c Rv2654c 351 Rv3610c Rv2666 352 Rv3620c Rv2680 353 Rv3628 Rv2684 354 Rv3648c Rv2685 355 Rv3676 Rv2697c 356 Rv3678A Rv2703 357 Rv3692 Rv2711 358 Rv3708c Rv2731 359 Rv3709c Rv2754c 360 Rv3710 Rv2775 361 Rv3715c Rv2788 362 Rv3753c Rv2795c 363 Rv3783 Rv2806 364 Rv3789 Rv2830c 365 Rv3791 Rv2835c 366 Rv3809c Rv2837c 367 Rv3842c Rv2841c 368 Rv3856c Rv2863 369 Rv3859c Rv2878c Rv2885c Rv2889c Rv2903c Rv2907c Rv2909c Rv2914c Rv2925c Rv2928 Rv2937 Rv2944 Rv2945c RV2951C Rv2975c Rv2984 Rv2996c Rv3002c Rv3007c Rv3009c Rv3013 Rv3019c Rv3020c Rv3022c Rv3024c Rv3028c Rv3042c Rv3050c Rv3061c Rv3071 Rv3072c Rv3076 Rv3089 Rv3106 Rv3115 Rv3118 Rv3132c Rv3160c Rv3169 Rv3179 Rv3196 Rv3198c RV3213C Rv3218 Rv3221c Rv3232c Rv3237c Rv3248c Rv3257c Rv3283 RV3284 Rv3285 Rv3300c Rv3309c Rv3311 Rv3315c Rv3321c Rv3322c Rv3341 Rv3349c Rv3385c Rv3395c Rv3404c Rv3406 Rv3412 Rv3416 Rv3417c Rv3437 Rv3442c Rv3446c RV3455C Rv3457c Rv3465 Rv3477 RV3503C Rv3519 Rv3551 Rv3555c Rv3572 Rv3575c Rv3583c Rv3600c Rv3609c Rv3614c Rv3628 Rv3653 Rv3672c Rv3676 Rv3688c Rv3703c Rv3710 Rv3717 Rv3733c Rv3735 Rv3749c Rv3755c Rv3756c Rv3760 Rv3768 Rv3788 Rv3791 Rv3799c Rv3836 Rv3841 Rv3855 Rv3862c Rv3872 Rv3882c Rv3890c Rv3908 Rv3918c -
TB is an ancient infectious respiratory disease that seriously endangers human health. Almost a quarter of the world’s population is estimated to have latent TB infection (LTBI). Numerous potential risk factors may promote the progression of LTBI to active TB once the efficiency of the immune system declines[16]. Therefore, there is an imperative need to develop new improved vaccines that protect against both active TB and LTBI[17]. To date, vaccination continues to be the most effective method to prevent and control TB[18]. However, the protection capacity of traditional BCG vaccines is limited. As a result, new strategies to improve the effectiveness of vaccination are required.
Bacterial proteins are important potential candidates for the development of new anti-TB vaccines because they can interact with host cells as key cell antigens[19]. Several M. tuberculosis secretory proteins have been used in newly designed anti-TB vaccines, and some of them showed excellent immune responses and protective efficacy against M. tuberculosis infection[20-22]. For example, Ag85, EAST-6, CFP10, and Rv1886c are excellent antigens; one or more of these proteins have been used in subunit and viral-vector vaccines, some of which have successfully entered phases I–III clinical trials and showed better protection ability than BCG[23-26].
A recent study showed that mycobacterial extracts from the rapidly growing NTM M. vaccae used in vaccines had promising protective effects against TB[12]. Several vaccines based on M. vaccae antigens, such as Vaccae™, DAR-901, and SRL172, have entered different phases of clinical trials[27]. SRL172, an inactivated, whole-cell vaccine prepared from M. vaccae, was safe, well-tolerated, and immunogenic in humans[13,28]. A randomized controlled phase III trial in Tanzania demonstrated that boosting with SRL172 could protect against culture-confirmed TB in HIV-infected adults who had received BCG at birth[29]. The excellent immune effect of M. vaccae indicates that NTMs could also serve as a good source for designing new effective anti-TB vaccines. In the present study, we evaluated the immunogenicity and protective efficacy of M. intracellulare whole bacterial protein extracts in BALB/c mice to explore the possibility of using M. intracellulare as a source of immune effective antigens for developing anti-TB vaccines. Our results showed that M. intracellulare bacterial proteins promote high-level production of IFN-γ, IL-2, IL-6, and IL-12 in mice, while IL-4 remained at a very low level, thus proving that immunization with M. intracellulare bacterial proteins predominantly elicited Th1-type cytokine production in BALB/c mice. Although the immune mechanism of vaccine protection from TB remains unclear, high expression levels of IFN-γ have been associated with enhanced protection against mycobacterial infection[30, 31], and studies have shown that IFN-γ responses against multiple mycobacterial antigens could predict protection against TB[32, 33]. IL-2 is also an important indicator in most clinical trials for evaluating TB-vaccine efficacy; high IL-2 levels indicate high concentrations of vaccine-induced activated CD4 and/or CD8 cells. These cells are necessary, although not sufficient, for protective immunity against M. tuberculosis in both animal models and humans[34,35]. IL-6 is a well-known inflammatory marker cytokine. Although several studies have confirmed that the level of IL-6 could be used to monitor the progress of infection and infer the risk of progression to active TB, its role in vaccine-mediated immune protection against TB is still unclear[36]. A previous study showed that IL-6 plays a major role in priming but not in late Th1 response to a TB vaccine, thus regulating the phenotype of the immune response[37]. The high concentration of IL-6 detected in this study was possibly induced by the antigenic proteins in M. intracellulare, which are also critical for immunity[38]. IL-12 is a key cytokine that mediates the immune response of Th1 cells. Its main immunomodulatory effect is to induce differentiation of early T helper cells into Th1 cells and promote the development and proliferation of Th1 cells. High levels of IL-12 indicate cellular immunity rather than humoral immunity[39]. A recent study reported that IL-12 production could also inhibit intracellular mycobacterial growth by enhancing autophagy[40]. Another cytokine we detected is IL-4, which is a central cytokine produced by Th2 cells and promotes B cell proliferation[41]. We found that IL-4 was at a very low concentration in both groups, and there was no significant difference between the two groups.
Data from previous studies suggested that both Th1 and Th2 immune responses play important roles in host protection against M. tuberculosis infection, and Th1 is essential against this intracellular pathogen. Current attempts to generate a vaccine against TB are generally based on the assumption that it must drive a Th1 response[42]. Our results showed that M. intracellulare proteins could induce a Th1/Th2 balance shift toward Th1 in BALB/c mice, which favors the control of M. tuberculosis in vivo.
While the role of cell-mediated immunity in the protective immune response against TB has been well established, the role of B cells in this process is not clearly understood[43]. Emerging evidence suggests that B cell-dominated humoral immunity can modulate the host immune response to various intracellular pathogens, including M. tuberculosis, by regulating the level of granulomatous reaction, cytokine production, and T cell response[44,45]. Therefore, we also monitored antibody production in our animal model. The levels of the four antibodies (IgG, IgG1, IgM, and IgG2a) were measured in a time- and dose-dependent manner; results showed an increase in antibody titer levels. After three immunizations, they all reached a significantly higher quantity than that in the negative control group mice, suggesting that M. intracellulare bacterial proteins potentially improve the humoral immune response in mice. The results also indicated that PolyI:C and DDA function well as subcutaneous adjuvants to promote the production of antibodies without eliciting immune responses of their own.
Macrophages are the first line of defense of the host immune system against M. tuberculosis infection[46,47]. During its long-term interaction with macrophages, M. tuberculosis develops many effective strategies to avoid elimination, while surviving and proliferating inside macrophages[48]. Although a series of complex interactions between the host and pathogen ultimately determine the outcome of infection, the mechanisms of macrophage-bacillus interactions are complicated and still under investigation. The improved clearance ability of macrophages is essential for the host to fight TB. Our results showed that BMDMs from M. intracellulare-immunized mice had a significant ability to inhibit the growth of M. tuberculosis compared to those from control mice in vitro, indicating that M. intracellulare bacterial proteins can effectively change the host immune system environment to impair M. tuberculosis survival. One explanation for this result may be that bacterial protein immunization upregulates the expression of specific genes in BMDMs and thus helps to impede M. tuberculosis growth. Therefore, more specific experiments, including transcriptome analysis or other methods, are required to reveal the relevant mechanisms. Our results also suggest the possibility of using M. intracellulare antigenic proteins for the development of TB vaccines.
To gain insights into the antigens expressed among the M. intracellulare bacterial proteins that changed the immune properties of mice to prevent M. tuberculosis infection, a protein microarray coated with 4,219 proteins of M. tuberculosis was used to identify the antigen repertoire by cross-reacting with IgG and IgM in the sera of mice immunized with whole M. intracellulare proteins. A total of 478 M. tuberculosis proteins were recognized by the serum antibodies IgG and IgM. Through GO enrichment analysis and KEGG pathway analysis of the 478 proteins as well as the comparative genomic analysis between M. intracellulare and M. tuberculosis, we primarily obtained an insight into the main biological processes and pathways involved in shared antigens between M. tuberculosis and M. intracellulare. In the protein-protein interaction analysis, we selected specific antigens that formed three particular clusters: cell composition, cytoplasmic composition modification, and toxin-antitoxin system-related cluster. The strong interactions between these proteins suggest that they may work together to regulate the host immune profile. These protein combinations may therefore be potential targets for the development of anti-TB vaccines. In addition, we compared the antigens detected by combinational genomics and proteomics; among 478 genes and 369 proteins identified in protein microarray and comparative genomic analysis, 60 common antigens were found. These antigens may possibly be potential cross-reactive antigens that may be used for the subsequent development of cross-immune vaccines for both M. intracellulare and M. tuberculosis in the future.
In summary, the present study demonstrates the possibility of whole bacterial protein extracts of M. intracellulare to serve as antigens against TB vaccines and reveals a number of proteins with cross-reactivity between M. intracellulare and M. tuberculosis. One limitation of the present study is that we still cannot screen for more specific antigens with clear immunogenicity to provide candidates for TB vaccines that can effectively protect against M. tuberculosis infection. Further studies are required to cautiously choose immunodominant antigens, which are crucial for developing novel vaccines for the control of TB.
-
The authors declare that they have no competing interests.
-
FENG Wen Hai, LIU Hai Can, and LI Gui Lian designed this study, revised the manuscript, and edited the manuscript. XIAO Shi Qi prepared the first draft of the manuscript. XIAO Shi Qi, XU Da, DUAN Hong Yang, FAN Xue Ting, LI Xin Yao, LI Na, LI Ma Chao, and ZHAO Li Lan performed experiments. ZHAO Xiu Qin cultured the strains. ZHANG Wen and HAN Na performed genome sequencing of the M. intracellulare strain. WAN Kang Lin provided analysis support. All authors provided suggestions on the preparation of the manuscript and read and approved the final draft of the manuscript.
-
The authors thank all the staff working at the Laboratory Animal Center, the National Institute for Communicable Disease Control and Prevention, Chinese Center for Disease Control and Prevention for their support of this research.
doi: 10.3967/bes2021.073
Immunogenicity of Whole Mycobacterium intracellulare Proteins and Fingding on the Cross-Reactive Proteins between M. intracellulare and M. tuberculosis
-
Abstract:
Objectives To evaluate the immunogenicity of Mycobacterium intracellulare proteins and determine the cross-reactive proteins between M. intracellulare and M. tuberculosis. Methods Protein extracts from M. intracellulare were used to immunize BALB/c mice. The antigens were evaluated using cellular and humoral immunoassays. The common genes between M. intracellular and M. tuberculosis were identified using genome-wide comparative analysis, and cross-reactive proteins were screened using immunoproteome microarrays. Results Immunization with M. intracellulare proteins induced significantly higher levels of the cytokines interferon-γ (IFN-γ), interleukin-2 (IL-2), interleukin-12 (IL-12), interleukin-6 (IL-6) and immunoglobulins IgG, IgG1, IgM, and IgG2a in mouse serum. Bone marrow-derived macrophages isolated from mice immunized with M. intracellulare antigens displayed significantly lower bacillary loads than those isolated from mice immunized with adjuvants. Whole-genome sequence analysis revealed 396 common genes between M. intracellulare and M. tuberculosis. Microchip hybridization with M. tuberculosis proteins revealed the presence of 478 proteins in the serum of mice immunized with M. intracellulare protein extracts. Sixty common antigens were found using both microchip and genomic comparative analyses. Conclusion This is the advanced study to investigate the immunogenicity of M. intracellulare proteins and the cross-reactive proteins between M. intracellulare and M. tuberculosis. The results revealed the presence of a number of cross-reactive proteins between M. intracellulare and M. tuberculosis. Therefore, this study provides a new way of identifying immunogenic proteins for use in tuberculosis vaccines against both M. intracellulare and M. tuberculosis in future. -
Figure 1. Cellular immune responses in immunized BALB/c mice. Lymphocytes isolated from BALB/c mice immunized with M. intracellulare protein extracts or adjuvants only were stimulated with M. intracellulare protein extracts, and the concentration of five different cytokines (A) IFN-γ, (B) IL-2, (C) IL-12, (D) IL-4, and (E) IL-6 were determined after 48 h. Capped line with asterisk * indicates significant difference between the two immunization methods, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Columns indicate the mean from at least three samples, and error bars denote the standard deviation (SD)
Figure 2. Antibody level monitoring in mice that had received three interval immunizations with M. intracellulare protein extracts or adjuvants. The change trends of titer levels of IgG, IgG1, IgM, and IgG2a are shown in (A), (B), (C), and (D), respectively. Dotted lines show the antibody titer levels in BALB/c mice immunized with M. intracellulare bacterial proteins + PolyI:C and DDA as an adjuvant; lines with squares show the antibody titer levels in BALB/c mice immunized with PolyI:C and DDA. The antibody titer levels were monitored at day 0, day 10 (10 days after the first immunization), day 20 (10 days after the second immunization), and day 30 (10 days after the third immunization). Points indicate the mean of at least three different serum samples in a group, and error bars denote standard deviation (SD).
Figure 3. Immunization with M. intracellulare protein extracts enhanced the ability of bone marrow-derived macrophages to inhibit M. tuberculosis growth ex vivo. (A) BMDMs were infected with H37Rv at MOI = 3. (B) BMDMs were infected with H37Rv at MOI = 5. Colony counts were obtained for 0, 3, and 5 days of BMDM infection were compared with those obtained for H37RV. Points indicate the mean from triplicate cultures, and error bars denote standard deviation (SD). P-values were determined with two-tailed Student's t test.
Figure 4. Top 15 GO terms identified based on the proteins found in the serum of M. intracellulare immunized mice. The results include three main categories: red bars represent biological processes, green bars represent molecular functions, and blue bars represent cellular components. The x-axis shows the percentage of the specific GO term in each category. GO, gene ontology.
Figure 5. Kyoto encyclopedia of genes and genomes (KEGG) pathway analysis for screened antigens. Blue bars represent each category with the name labeled in the left; x-axis shows the number of antigens in each specific KEGG term in each category.
Figure 6. Protein-protein interaction network. The protein interaction network was constructed based on the String database. (A) Map of the interaction network of all screened antigenic proteins. (B) Cell composition (GO:0044464) correlated cluster. (C) Cytoplasmic composition modification (GO:0005737) correlated cluster. (D) Toxin-antitoxin system (kw-1277) correlated cluster. Lines represent specific protein-protein associations. GO, gene ontology.
Figure 7. Comparative analysis of antigens in protein microarray and comparative genomic analysis. Venn diagram showing the distribution of common antigens and exclusive antigens between protein microarrays (A) and comparative genomic analysis (B). The numbers shown in the diagram represent the number of proteins in each group.
Table 1. Annotation information for the two genomic sequences
Characters NC_000962* NC_016946** Genome size (Mb) 4.41 5.4 GC (%) 65.60 68.10 No. of proteins 3,906 5,000 No. of rRNA 3 3 No. of tRNA 45 46 No. of other RNA 22 3 No. of genes 4,008 5,104 No. of pseudogenes 30 52 Note. *https://www.ncbi.nlm.nih.gov/genome/166?genome_assembly_id=159857. **https://www.ncbi.nlm.nih.gov/genome/1703?genome_assembly_id=171542.  下载: 导出CSV
下载: 导出CSV
Table 2. Functional classification of the 369 common genes found in the genomes of M. tuberculosis and M. intracellulare
No. Function Number 1 Virulence, detoxification, adaptation 10 2 Lipid metabolism 35 3 Information pathways 73 4 Cell wall and cell processes 39 5 Intermediary metabolism and respiration 143 6 Regulatory proteins 24 7 Conserved hypotheticals 41
下载: 导出CSV
S1. Names, functions, and pathways of 478 proteins identified in the sera of M. intracellulare-immunized mice by protein microarrays
ID KEGG pathway No. of genes Genes mapped mtu01100 Metabolic
pathways30 mtu:Rv0091 mtn; 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase mtu:Rv0500 proC; pyrroline-5-carboxylate reductase mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0769 oxidoreductase mtu:Rv0848 cysK2; cysteine synthase CysK mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv0956 purN; phosphoribosylglycinamide formyltransferase PurN mtu:Rv1092c coaA; pantothenate kinase mtu:Rv1164 narI; nitrate reductase subunit gamma mtu:Rv1257c oxidoreductase mtu:Rv1263 amiB2; amidase AmiB mtu:Rv1306 atpF; ATP synthase subunit B mtu:Rv1308 atpA; ATP synthase subunit alpha mtu:Rv1448c tal; transaldolase mtu:Rv1530 adh; alcohol dehydrogenase mtu:Rv1604 impA; inositol-monophosphatase ImpA mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2195 qcrA; ubiquinol-cytochrome C reductase rieske iron-sulfur subunit mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2350c plcB; membrane-associated phospholipase B mtu:Rv2377c hypothetical protein mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3009c gatB; aspartyl/glutamyl-tRNA(Asn/Gln) amidotransferase subunit B mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu:Rv3106 fprA; NADPH-ferredoxin reductase FprA mtu:Rv3465 rmlC; dTDP-4-dehydrorhamnose 3,5-epimerase mtu:Rv3600c type III pantothenate kinase mtu:Rv3703c etgB; iron(II)-dependent oxidoreductase EgtB mtu01110 Biosynthesis of
secondary
metabolites19 mtu:Rv0500 proC; pyrroline-5-carboxylate reductase mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0769 oxidoreductase mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv0956 purN; phosphoribosylglycinamide formyltransferase PurN mtu:Rv1086 (2Z,6E)-farnesyl diphosphate synthase mtu:Rv1257c oxidoreductase mtu:Rv1448c tal; transaldolase mtu:Rv1530 adh; alcohol dehydrogenase mtu:Rv1604 impA; inositol-monophosphatase ImpA mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2350c plcB; membrane-associated phospholipase B mtu:Rv2377c hypothetical protein mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu:Rv3465 rmlC; dTDP-4-dehydrorhamnose 3,5-epimerase mtu01120 Microbial
metabolism in
diverse
environments12 mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv1164 narI; nitrate reductase subunit gamma mtu:Rv1257c oxidoreductase mtu:Rv1263 amiB2; amidase AmiB mtu:Rv1448c tal; transaldolase mtu:Rv1530 adh; alcohol dehydrogenase mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu01230 Biosynthesis of
amino acids7 mtu:Rv0091 mtn; 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase mtu:Rv0500 proC; pyrroline-5-carboxylate reductase mtu:Rv1448c tal; transaldolase mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu01200 Carbon
metabolism6 mtu:Rv0952 sucD; succinyl-CoA ligase subunit alpha mtu:Rv1448c tal; transaldolase mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu:Rv2465c rpiB; ribose-5-phosphate isomerase B mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu:Rv3042c serB2; phosphoserine phosphatase SerB mtu01240 Biosynthesis of
cofactors4 mtu:Rv0511 hemD; uroporphyrin-III C-methyltransferase mtu:Rv0769 oxidoreductase mtu:Rv1092c coaA; pantothenate kinase mtu:Rv3600c type III pantothenate kinase mtu02024 Quorum sensing 4 mtu:Rv0732 secY; preprotein translocase SecY mtu:Rv1609 trpE; anthranilate synthase component I mtu:Rv2350c plcB; membrane-associated phospholipase B mtu:Rv3676 crp; cAMP receptor protein mtu00190 Oxidative
phosphorylation4 mtu:Rv1306 atpF; ATP synthase subunit B mtu:Rv1308 atpA; ATP synthase subunit alpha mtu:Rv2195 qcrA; ubiquinol-cytochrome C reductase rieske iron-sulfur subunit mtu:Rv3628 ppa; inorganic pyrophosphatase mtu02020 Two-component
system4 mtu:Rv1164 narI; nitrate reductase subunit gamma mtu:Rv2234 ptpA; protein-tyrosine-phosphatase mtu:Rv3132c devS; two component sensor histidine kinase DevS mtu:Rv3676 crp; cAMP receptor protein mtu00270 Cysteine and
methionine
metabolism3 mtu:Rv0091 mtn; 5'-methylthioadenosine/S-adenosylhomocysteine nucleosidase mtu:Rv0848 cysK2; cysteine synthase CysK mtu:Rv2996c serA1; D-3-phosphoglycerate dehydrogenase mtu00380 Tryptophan
metabolism3 mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv1263 amiB2; amidase AmiB mtu:Rv2215 dlaT; pyruvate dehydrogenase E2 component dihydrolipoamide acyltransferase mtu03018 RNA
degradation3 mtu:Rv0350 dnaK; chaperone protein DnaK mtu:Rv0440 groEL2; molecular chaperone GroEL mtu:Rv3417c groEL1; chaperonin GroEL mtu05152 Tuberculosis 3 mtu:Rv0350 dnaK; chaperone protein DnaK mtu:Rv0440 groEL2; molecular chaperone GroEL mtu:Rv3417c groEL1; chaperonin GroEL mtu03430 Mismatch repair 2 mtu:Rv0002 dnaN; DNA polymerase III subunit beta mtu:Rv2413c hypothetical protein mtu01212 Fatty acid
metabolism2 mtu:Rv0673 echA4; enoyl-CoA hydratase EchA4 mtu:Rv0769 oxidoreductase
下载: 导出CSV
S2. Names of common antigens identified by microchip and comparative genomic analysis
No. Genome BioChip Common 1 Rv0009 MT0066.1 Rv0009 2 Rv0011c MT0066.2 Rv0046c 3 Rv0019c MT0116.1 Rv0053 4 Rv0046c MT0250 Rv0337c 5 Rv0053 MT0270.1 Rv0350 6 Rv0055 MT0383 Rv0384c 7 Rv0126 MT0392 Rv0423c 8 Rv0130 MT0407 Rv0440 9 Rv0137c MT0470 Rv0491 10 Rv0156 MT0487 Rv0551c 11 Rv0157 MT0543 Rv0667 12 Rv0158 MT0555 Rv0672 13 Rv0189c MT0610 Rv0673 14 Rv0211 MT0946 Rv0684 15 Rv0230c MT0968.1 Rv0685 16 Rv0236A MT1029 Rv0732 17 Rv0238 MT1040.1 Rv0861c 18 Rv0242c MT1055 Rv0984 19 Rv0244c MT1083.1 Rv1013 20 Rv0267 MT1172.1 Rv1070c 21 Rv0321 MT1264.1 Rv1080c 22 Rv0337c MT1305.1 Rv1086 23 Rv0350 MT1329 Rv1092c 24 Rv0352 MT1479.1 Rv1257c 25 Rv0357c MT1534 Rv1292 26 Rv0363c MT1555.1 Rv1308 27 Rv0380c MT1775 Rv1458c 28 Rv0384c MT1849.1 Rv1654 29 Rv0391 MT2068 Rv1829 30 Rv0407 MT2113 Rv2115c 31 Rv0411c MT2138.2 Rv2241 32 Rv0423c MT2142 Rv2346c 33 Rv0430 MT2283 Rv2374c 34 Rv0440 MT2291 Rv2457c 35 Rv0458 MT2316 Rv2460c 36 Rv0465c MT2330.1 Rv2465c 37 Rv0467 MT2334.1 Rv2477c 38 Rv0491 MT2361.1 Rv2534c 39 Rv0498 MT2405 Rv2558 40 Rv0500A MT2455 Rv2697c 41 Rv0510 MT2488.1 Rv2711 42 Rv0527 MT2501 Rv2754c 43 Rv0548c MT2502 Rv2788 44 Rv0551c MT2520.1 Rv2795c 45 Rv0566c MT2554.1 Rv2909c 46 Rv0634B MT2625 Rv3009c 47 Rv0636 MT2626 Rv3028c 48 Rv0639 MT2637.1 Rv3118 49 Rv0640 MT2721 Rv3221c 50 Rv0641 MT2779 Rv3248c 51 Rv0642c MT2871 Rv3412 52 Rv0647c MT2958.1 Rv3442c 53 Rv0651 MT3139.1 Rv3457c 54 Rv0652 MT3270.1 Rv3551 55 Rv0655 MT3279 Rv3583c 56 Rv0667 MT3284 Rv3609c 57 Rv0668 MT3289 Rv3628 58 Rv0672 MT3290.2 Rv3676 59 Rv0673 MT3532.1 Rv3710 60 Rv0682 MT3573.12 Rv3791 61 Rv0683 MT3631 62 Rv0684 MT3770 63 Rv0685 MT3858 64 Rv0691A MT3876 65 Rv0693 MT3878 66 Rv0700 MT4026.1 67 Rv0701 Rv0002 68 Rv0702 Rv0006 69 Rv0703 Rv0009 70 Rv0704 Rv0022c 71 Rv0705 Rv0025 72 Rv0707 Rv0028 73 Rv0708 rv0036 74 Rv0709 Rv0040c 75 Rv0714 Rv0041 76 Rv0716 Rv0043c 77 Rv0717 Rv0045c 78 Rv0718 Rv0046c 79 Rv0719 Rv0053 80 Rv0721 Rv0063 81 Rv0723 Rv0076c 82 Rv0732 Rv0089 83 Rv0733 Rv0091 84 Rv0737 Rv0095c 85 Rv0753c Rv0098 86 Rv0803 Rv0100 87 Rv0808 Rv0110 88 Rv0814c Rv0119 89 Rv0815c Rv0145 90 Rv0820 Rv0150c 91 Rv0821c Rv0155 92 Rv0859 Rv0180c 93 Rv0861c Rv0185 94 Rv0889c Rv0187 95 Rv0896 Rv0226c 96 Rv0903c Rv0232 97 Rv0946c Rv0248c 98 Rv0958 Rv0250c 99 Rv0974c Rv0264c 100 Rv0975c Rv0277c 101 Rv0981 Rv0281 102 Rv0984 Rv0285 103 Rv1013 Rv0287 104 Rv1017c Rv0288 105 Rv1019 Rv0290 106 Rv1023 Rv0295c 107 Rv1038c Rv0299 108 Rv1070c Rv0301 109 Rv1074c Rv0308 110 Rv1077 Rv0333 111 Rv1080c Rv0337c 112 Rv1086 Rv0350 113 Rv1092c Rv0369c 114 Rv1095 Rv0379 115 Rv1098c Rv0384c 116 Rv1099c Rv0385 117 Rv1151c Rv0387c 118 Rv1177 Rv0398c 119 Rv1187 Rv0423c 120 Rv1197 Rv0429c 121 Rv1198 Rv0437c 122 Rv1211 Rv0440 123 Rv1213 Rv0446c 124 Rv1240 Rv0459 125 Rv1248c Rv0466 126 Rv1257c Rv0489 127 Rv1262c Rv0491 128 Rv1292 Rv0500 129 Rv1298 Rv0508 130 Rv1305 Rv0511 131 Rv1308 Rv0514 132 Rv1310 Rv0518 133 Rv1311 Rv0521c 134 Rv1315 Rv0546c 135 Rv1321 Rv0551c 136 Rv1331 Rv0561c 137 Rv1380 Rv0571c 138 Rv1381 Rv0577 139 Rv1383 Rv0579 140 Rv1384 Rv0580c 141 Rv1388 Rv0598c 142 Rv1392 Rv0600c 143 Rv1415 Rv0603 144 Rv1423 Rv0604 145 Rv1436 Rv0606 146 Rv1447c Rv0612 147 Rv1458c Rv0635 148 Rv1474c Rv0657c 149 Rv1475c Rv0659c 150 Rv1479 Rv0666 151 Rv1481 Rv0667 152 Rv1483 Rv0672 153 Rv1484 Rv0673 154 Rv1488 Rv0678 155 Rv1493 Rv0684 156 Rv1547 Rv0685 157 Rv1589 Rv0699 158 Rv1601 Rv0730 159 Rv1611 Rv0731c 160 Rv1617 Rv0732 161 Rv1627c Rv0750 162 Rv1630 Rv0764c 163 Rv1633 Rv0766c 164 Rv1638 Rv0769 165 Rv1641 Rv0772 166 Rv1642 Rv0784 167 Rv1643 Rv0790c 168 Rv1654 Rv0793 169 Rv1655 Rv0801 170 Rv1657 Rv0810c 171 Rv1658 Rv0819 172 Rv1659 Rv0828c 173 Rv1729c Rv0837c 174 Rv1730c Rv0848 175 Rv1731 Rv0857 176 Rv1783 Rv0861c 177 Rv1793 Rv0865 178 Rv1821 Rv0869c 179 Rv1827 Rv0882 180 Rv1829 Rv0887c 181 Rv2050 Rv0937c 182 Rv2062c Rv0950c 183 Rv2097c Rv0952 184 Rv2111c Rv0954 185 Rv2112c Rv0956 186 Rv2115c Rv0970 187 Rv2122c Rv0984 188 Rv2134c Rv1008 189 Rv2146c Rv1012 190 Rv2150c RV1013 191 Rv2156c Rv1046c 192 Rv2166c Rv1056 193 Rv2178c Rv1070c 194 Rv2193 Rv1080c 195 Rv2204c Rv1083 196 Rv2218 Rv1086 197 Rv2220 Rv1092c 198 Rv2222c Rv1097c 199 Rv2225 Rv1109c 200 Rv2241 Rv1112 201 Rv2244 Rv1113 202 Rv2245 Rv1124 203 Rv2247 Rv1132 204 Rv2256c Rv1153c 205 Rv2259 Rv1164 206 Rv2346c RV1181 207 Rv2347c Rv1202 208 Rv2362c Rv1208 209 Rv2367c Rv1239c 210 Rv2374c Rv1257c 211 Rv2375 RV1259 212 Rv2402 Rv1263 213 Rv2404c RV1264 214 Rv2406c Rv1275 215 Rv2412 RV1282C 216 Rv2420c Rv1284 217 Rv2421c Rv1292 218 Rv2426c Rv1306 219 Rv2428 Rv1308 220 Rv2441c Rv1309 221 Rv2442c RV1312 222 Rv2448c RV1328 223 Rv2457c Rv1371 224 Rv2460c Rv1373 225 Rv2461c Rv1377c 226 Rv2465c RV1395 227 Rv2466c RV1404 228 Rv2477c Rv1414 229 Rv2502c Rv1427c 230 Rv2511 Rv1428c 231 Rv2534c Rv1448c 232 Rv2539c Rv1451 233 Rv2540c Rv1458c 234 Rv2558 Rv1462 235 Rv2572c Rv1463 236 Rv2583c Rv1476 237 Rv2592c Rv1501 238 Rv2603c Rv1530 239 Rv2605c Rv1531 240 Rv2606c Rv1536 241 Rv2674 Rv1544 242 Rv2676c Rv1584c 243 Rv2692 Rv1604 244 Rv2697c Rv1609 245 Rv2699c RV1650 246 Rv2708c Rv1654 247 Rv2710 Rv1677 248 Rv2711 Rv1685c 249 Rv2713 Rv1692 250 Rv2720 Rv1693 251 Rv2725c Rv1695 252 Rv2733c Rv1708 253 Rv2744c Rv1710 254 Rv2754c Rv1717 255 Rv2764c RV1718 256 Rv2783c Rv1719 257 Rv2788 Rv1724c 258 Rv2795c Rv1742 259 Rv2831 Rv1770 260 Rv2840c Rv1791 261 Rv2861c Rv1806 262 Rv2868c Rv1828 263 Rv2882c Rv1829 264 Rv2890c RV1837c 265 Rv2901c Rv1848 266 Rv2904c Rv1875 267 Rv2909c Rv1876 268 Rv2911 Rv1893 269 Rv2919c Rv1894c 270 Rv2927c Rv1896c 271 Rv2965c Rv1898 272 Rv2975a RV1912C 273 Rv2987c Rv1936 274 Rv3003c Rv1959c 275 Rv3009c Rv1968 276 Rv3011c Rv1988 277 Rv3012c Rv1992c 278 Rv3028c Rv2021c 279 Rv3029c Rv2042c 280 Rv3043c Rv2043c 281 Rv3048c Rv2049c 282 Rv3051c RV2089C 283 Rv3053c Rv2098c 284 Rv3102c Rv2099c 285 Rv3105c Rv2102 286 Rv3117 Rv2104c 287 Rv3118 Rv2107 288 Rv3146 Rv2115c 289 Rv3148 Rv2130c 290 Rv3150 Rv2135c 291 Rv3155 Rv2140c 292 Rv3219 Rv2145c 293 Rv3221c Rv2158c 294 Rv3240c Rv2163c 295 Rv3246c Rv2195 296 Rv3248c Rv2198c 297 Rv3270 Rv2215 298 Rv3280 Rv2226 299 Rv3303c Rv2229c 300 Rv3318 Rv2233 301 Rv3319 Rv2234 302 Rv3339c Rv2239c 303 Rv3340 Rv2241 304 Rv3356c Rv2258c 305 Rv3362c Rv2293c 306 Rv3368c Rv2299c 307 Rv3370c Rv2321c 308 Rv3396c Rv2324 309 Rv3409c Rv2346c 310 Rv3410c Rv2350c 311 Rv3411c Rv2351c 312 Rv3412 Rv2360c 313 Rv3418c Rv2368c 314 Rv3432c Rv2371 315 Rv3436c Rv2374c 316 Rv3442c Rv2376c 317 Rv3443c Rv2377c 318 Rv3457c Rv2413c 319 Rv3458c Rv2431c 320 Rv3459c Rv2436 321 Rv3460c RV2438C 322 Rv3461c Rv2445c 323 Rv3462c Rv2452c 324 Rv3464 Rv2457c 325 Rv3489 Rv2460c 326 Rv3501c Rv2465c 327 Rv3515c RV2467 328 Rv3516 Rv2471 329 Rv3526 Rv2473 330 Rv3534c Rv2477c 331 Rv3535c Rv2493 332 Rv3543c Rv2499c 333 Rv3550 Rv2505c 334 Rv3551 Rv2517c 335 Rv3553 Rv2518c 336 Rv3556c Rv2528c 337 Rv3557c Rv2534c 338 Rv3559c Rv2546 339 Rv3560c Rv2555c 340 Rv3562 Rv2558 341 Rv3567c Rv2561 342 Rv3568c Rv2564 343 Rv3570c Rv2576c 344 Rv3574 Rv2579 345 Rv3583c Rv2595 346 Rv3586 Rv2614A 347 Rv3592 Rv2631 348 Rv3596c Rv2638 349 Rv3597c Rv2641 350 Rv3609c Rv2654c 351 Rv3610c Rv2666 352 Rv3620c Rv2680 353 Rv3628 Rv2684 354 Rv3648c Rv2685 355 Rv3676 Rv2697c 356 Rv3678A Rv2703 357 Rv3692 Rv2711 358 Rv3708c Rv2731 359 Rv3709c Rv2754c 360 Rv3710 Rv2775 361 Rv3715c Rv2788 362 Rv3753c Rv2795c 363 Rv3783 Rv2806 364 Rv3789 Rv2830c 365 Rv3791 Rv2835c 366 Rv3809c Rv2837c 367 Rv3842c Rv2841c 368 Rv3856c Rv2863 369 Rv3859c Rv2878c Rv2885c Rv2889c Rv2903c Rv2907c Rv2909c Rv2914c Rv2925c Rv2928 Rv2937 Rv2944 Rv2945c RV2951C Rv2975c Rv2984 Rv2996c Rv3002c Rv3007c Rv3009c Rv3013 Rv3019c Rv3020c Rv3022c Rv3024c Rv3028c Rv3042c Rv3050c Rv3061c Rv3071 Rv3072c Rv3076 Rv3089 Rv3106 Rv3115 Rv3118 Rv3132c Rv3160c Rv3169 Rv3179 Rv3196 Rv3198c RV3213C Rv3218 Rv3221c Rv3232c Rv3237c Rv3248c Rv3257c Rv3283 RV3284 Rv3285 Rv3300c Rv3309c Rv3311 Rv3315c Rv3321c Rv3322c Rv3341 Rv3349c Rv3385c Rv3395c Rv3404c Rv3406 Rv3412 Rv3416 Rv3417c Rv3437 Rv3442c Rv3446c RV3455C Rv3457c Rv3465 Rv3477 RV3503C Rv3519 Rv3551 Rv3555c Rv3572 Rv3575c Rv3583c Rv3600c Rv3609c Rv3614c Rv3628 Rv3653 Rv3672c Rv3676 Rv3688c Rv3703c Rv3710 Rv3717 Rv3733c Rv3735 Rv3749c Rv3755c Rv3756c Rv3760 Rv3768 Rv3788 Rv3791 Rv3799c Rv3836 Rv3841 Rv3855 Rv3862c Rv3872 Rv3882c Rv3890c Rv3908 Rv3918c
下载: 导出CSV
-
[1] WHO. Global tuberculosis report 2019. Geneva: World Health Organization, 2019. [2] De Rycker M, Baragaña B, Duce SL, et al. Challenges and recent progress in drug discovery for tropical diseases. Nature, 2018; 559, 498−506. doi: 10.1038/s41586-018-0327-4 [3] Abdallah AM, Behr MA. Evolution and strain variation in BCG. In: Gagneux S. Strain Variation in the Mycobacterium tuberculosis Complex: Its Role in Biology, Epidemiology and Control. Advances in Experimental Medicine and Biology. Springer, 2017, 155-69. [4] Stallinger H, Kolleritsch H. BCG-vaccination and its complications. How great are the benefits of vaccination today? Klin Padiatr, 1990; 202, 308−14. doi: 10.1055/s-2007-1025537 [5] Chen SS, Xu YJ, Xiao SQ, et al. Analysis on human T cell epitopes polymorphisms of five specific antigens of Mycobacterium tuberculosis in 13 areas of China. Chin J Epidemiol, 2016; 37, 553−7. (In Chinese [6] Ginsberg AM. Designing tuberculosis vaccine efficacy trials-lessons from recent studies. Expert Rev Vaccines, 2019; 18, 423−32. doi: 10.1080/14760584.2019.1593143 [7] Jiang Y, Liu HC, Qiu Y, et al. Polymorphisms of FtsK/SpoIIIE protein in Mycobacterium tuberculosis complex strains may affect both protein function and host immune reaction. Int J Clin Exp Med, 2014; 7, 5385−93. [8] Jiang Y, Liu HC, Dou XF, et al. Polymorphisms of human T cell epitopes of Mycobacterium tuberculosis indicate divergence of host immune pressure on different categories of proteins. Life Sci, 2018; 209, 388−94. doi: 10.1016/j.lfs.2018.08.040 [9] Xiao TY, Liu HC, Li XQ, et al. Immunological evaluation of a novel Mycobacterium tuberculosis antigen Rv0674. Biomed Environ Sci, 2019; 32, 427−37. [10] Wang XZ, Chen SS, Xu YJ, et al. Identification and evaluation of the novel immunodominant antigen Rv2351c from Mycobacterium tuberculosis. Emerg Microbes Infect, 2017; 6, e48. [11] Jiang Y, Liu HC, Wang XZ, et al. Genetic diversity of immune-related antigens in region of difference 2 of Mycobacterium tuberculosis strains. Tuberculosis (Edinb), 2017; 104, 1−7. doi: 10.1016/j.tube.2016.05.002 [12] Bourinbaiar AS, Batbold U, Efremenko Y, et al. Phase III, placebo-controlled, randomized, double-blind trial of tableted, therapeutic TB vaccine (V7) containing heat-killed M. vaccae administered daily for one month. J Clin Tuberc Other Mycobact Dis, 2020; 18, 100141. doi: 10.1016/j.jctube.2019.100141 [13] Waddell RD, Chintu C, Lein AD, et al. Safety and immunogenicity of a five-dose series of inactivated Mycobacterium vaccae vaccination for the prevention of HIV-associated tuberculosis. Clin Infect Dis, 2000; 30, S309−15. doi: 10.1086/313880 [14] Feng ZH, Bai XY, Wang T, et al. Differential responses by human macrophages to infection with Mycobacterium tuberculosis and non-tuberculous mycobacteria. Front Microbiol, 2020; 11, 116. [15] Camus JC, Pryor MJ, Médigue C, et al. Re-annotation of the genome sequence of Mycobacterium tuberculosis H37Rv. Microbiology (Reading), 2002; 148, 2967−73. doi: 10.1099/00221287-148-10-2967 [16] Gao L, Li XW, Liu JM, et al. Incidence of active tuberculosis in individuals with latent tuberculosis infection in rural China: follow-up results of a population-based, multicentre, prospective cohort study. Lancet Infect Dis, 2017; 17, 1053−61. doi: 10.1016/S1473-3099(17)30402-4 [17] Salgame P, Geadas C, Collins L, et al. Latent tuberculosis infection-revisiting and revising concepts. Tuberculosis (Edinb), 2015; 95, 373−84. [18] Simmons JD, Stein CM, Seshadri C, et al. Immunological mechanisms of human resistance to persistent Mycobacterium tuberculosis infection. Nat Rev Immunol, 2018; 18, 575−89. doi: 10.1038/s41577-018-0025-3 [19] Liu CH, Liu HY, Ge BX. Innate immunity in tuberculosis: host defense vs pathogen evasion. Cell Mol Immunol, 2017; 14, 963−75. doi: 10.1038/cmi.2017.88 [20] Liu WB, Li JJ, Niu HX, et al. Immunogenicity and protective efficacy of multistage vaccine candidates (Mtb8.4-HspX and HspX-Mtb8.4) against Mycobacterium tuberculosis infection in mice. Int Immunopharmacol, 2017; 53, 83−9. doi: 10.1016/j.intimp.2017.10.015 [21] Yuan XF, Teng XD, Jing YK, et al. A live attenuated BCG vaccine overexpressing multistage antigens Ag85B and HspX provides superior protection against Mycobacterium tuberculosis infection. Appl Microbiol Biotechnol, 2015; 99, 10587−95. doi: 10.1007/s00253-015-6962-x [22] Yuan W, Dong N, Zhang LF, et al. Immunogenicity and protective efficacy of a tuberculosis DNA vaccine expressing a fusion protein of Ag85B-Esat6-HspX in mice. Vaccine, 2012; 30, 2490−7. doi: 10.1016/j.vaccine.2011.06.029 [23] Aagaard C, Hoang T, Dietrich J, et al. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat Med, 2011; 17, 189−94. doi: 10.1038/nm.2285 [24] Bertholet S, Ireton GC, Ordway DJ, et al. A defined tuberculosis vaccine candidate boosts BCG and protects against multidrug-resistant Mycobacterium tuberculosis. Sci Transl Med, 2010; 2, 53ra74. [25] Karbalaei Zadeh Babaki M, Soleimanpour S, Rezaee SA. Antigen 85 complex as a powerful Mycobacterium tuberculosis immunogene: biology, immune-pathogenicity, applications in diagnosis, and vaccine design. Microb Pathog, 2017; 112, 20−9. doi: 10.1016/j.micpath.2017.08.040 [26] Panigada M, Sturniolo T, Besozzi G, et al. Identification of a promiscuous T-cell epitope in Mycobacterium tuberculosis Mce proteins. Infect Immun, 2002; 70, 79−85. doi: 10.1128/IAI.70.1.79-85.2002 [27] Lahey T, Laddy D, Hill K, et al. Immunogenicity and protective efficacy of the DAR-901 booster vaccine in a murine model of tuberculosis. PLoS One, 2016; 11, e0168521. doi: 10.1371/journal.pone.0168521 [28] Vuola JM, Ristola MA, Cole B, et al. Immunogenicity of an inactivated mycobacterial vaccine for the prevention of HIV-associated tuberculosis: a randomized, controlled trial. AIDS, 2003; 17, 2351−5. doi: 10.1097/00002030-200311070-00010 [29] von Reyn CF, Mtei L, Arbeit RD, et al. Prevention of tuberculosis in Bacille Calmette-Guérin-primed, HIV-infected adults boosted with an inactivated whole-cell mycobacterial vaccine. AIDS, 2010; 24, 675−85. doi: 10.1097/QAD.0b013e3283350f1b [30] Jouanguy E, Altare F, Lamhamedi S, et al. Interferon-γ-receptor deficiency in an infant with fatal bacille Calmette-Guérin infection. N Engl J Med, 1996; 335, 1956−61. doi: 10.1056/NEJM199612263352604 [31] Newport MJ, Huxley CM, Huston S, et al. A mutation in the interferon-γ-receptor gene and susceptibility to mycobacterial infection. N Engl J Med, 1996; 335, 1941−9. doi: 10.1056/NEJM199612263352602 [32] Lahey T, Sheth S, Matee M, et al. Interferon γ responses to mycobacterial antigens protect against subsequent HIV-associated tuberculosis. J Infect Dis, 2010; 202, 1265−72. doi: 10.1086/656332 [33] Lahey T, Mitchell BK, Arbeit RD, et al. Polyantigenic interferon-γ responses are associated with protection from TB among HIV-infected adults with childhood BCG immunization. PLoS One, 2011; 6, e22074. doi: 10.1371/journal.pone.0022074 [34] Jasenosky LD, Scriba TJ, Hanekom WA, et al. T cells and adaptive immunity to Mycobacterium tuberculosis in humans. Immunol Rev, 2015; 264, 74−87. doi: 10.1111/imr.12274 [35] Lewinsohn DA, Lewinsohn DM, Scriba TJ. Polyfunctional CD4+ T cells as targets for tuberculosis vaccination. Front Immunol, 2017; 8, 1262. doi: 10.3389/fimmu.2017.01262 [36] Dalvi SM, Ramraje NN, Patil VW, et al. Study of IL-6 and vitamin D3 in patients of pulmonary tuberculosis. Indian J Tuberc, 2019; 66, 337−45. doi: 10.1016/j.ijtb.2018.05.018 [37] Leal IS, Flórido M, Andersen P, et al. Interleukin-6 regulates the phenotype of the immune response to a tuberculosis subunit vaccine. Immunology, 2001; 103, 375−81. doi: 10.1046/j.1365-2567.2001.01244.x [38] Jung BG, Wang XS, Yi N, et al. Early secreted antigenic target of 6-kDa of Mycobacterium tuberculosis stimulates IL-6 production by macrophages through activation of STAT3. Sci Rep, 2017; 7, 40984. doi: 10.1038/srep40984 [39] Martínez-Barricarte R, Markle JG, Ma CS, et al. Human IFN-γ immunity to mycobacteria is governed by both IL-12 and IL-23. Sci Immunol, 2018; 3, eaau6759. doi: 10.1126/sciimmunol.aau6759 [40] Mawatwal S, Behura A, Mishra A, et al. Calcimycin induced IL-12 production inhibits intracellular mycobacterial growth by enhancing autophagy. Cytokine, 2018; 111, 1−2. doi: 10.1016/j.cyto.2018.07.033 [41] Li QF, Zhang H, Yu L, et al. Down-regulation of Notch signaling pathway reverses the Th1/Th2 imbalance in tuberculosis patients. Int Immunopharmacol, 2018; 54, 24−32. doi: 10.1016/j.intimp.2017.10.026 [42] Rook GAW, Hernandez-Pando R, Dheda K, et al. IL-4 in tuberculosis: implications for vaccine design. Trends Immunol, 2004; 25, 483−8. doi: 10.1016/j.it.2004.06.005 [43] Abebe F. Synergy between Th1 and Th2 responses during Mycobacterium tuberculosis infection: a review of current understanding. Int Rev Immunol, 2019; 38, 172−9. doi: 10.1080/08830185.2019.1632842 [44] Chan J, Mehta S, Bharrhan S, et al. The role of B cells and humoral immunity in Mycobacterium tuberculosis infection. Semin Immunol, 2014; 26, 588−600. doi: 10.1016/j.smim.2014.10.005 [45] Khademi F, Yousefi A, Derakhshan M, et al. Enhancing immunogenicity of novel multistage subunit vaccine of Mycobacterium tuberculosis using PLGA: DDA hybrid nanoparticles and MPLA: subcutaneous administration. Iran J Basic Med Sci, 2019; 22, 893−900. [46] Upadhyay S, Mittal E, Philips JA. Tuberculosis and the art of macrophage manipulation. Pathog Dis, 2018; 76, fty037. [47] de Martino M, Lodi L, Galli L, et al. Immune response to Mycobacterium tuberculosis: a narrative review. Front Pediatr, 2019; 7, 350. doi: 10.3389/fped.2019.00350 [48] Hmama Z, Peña-Díaz S, Joseph S, et al. Immunoevasion and immunosuppression of the macrophage by Mycobacterium tuberculosis. Immunol Rev, 2015; 264, 220−32. doi: 10.1111/imr.12268 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 730
- HTML全文浏览量: 316
- PDF下载量: 75
- 被引次数: 0

 Quick Links
Quick Links