

 下载:
下载:

-
Aeromonas spp. are Gram-negative bacteria belonging to the Aeromonadaceae family and are widely distributed in the aquatic environment. Aeromonas has been isolated from dairy products (4%), vegetables (26%–41%), meat (5%–10%), poultry (3%–70%), and seafood (31%–72%)[1]. Aeromonas spp. are opportunistic pathogens that can directly infect or co-infect with other pathogenic bacteria, thus causing sepsis and meningitis in humans and animals[2]. Aeromonas spp. cause acute watery diarrhea, dysentery, and chronic gastrointestinal diseases[3], which pose threats to human health and quality of life. China continues to be one of the countries with the highest mortality rate owing to childhood diarrhea. Indeed, > 10,000 deaths from diarrheal diseases occur annually[4].
The Aeromonas genus currently consists of > 26 known species[5]. Owing to the limitations of the existing phenotypic identification systems and the complexity of the taxonomy of Aeromonas spp., phenotypic identification is deemed reasonably difficult to the species level, especially for Aeromonas spp. related to human diseases[6]. Furthermore, conventional methods, such as matrix-assisted laser desorption/ionization-time of flight mass spectrometry (MALDI-TOF MS), are time-consuming, labor-intensive, and error-prone[7]. For instance, Morinaga et al.[8] reported that isolates originally recognized as A. hydrophila using traditional phenotypic analysis have been re-identified as A. dhakensis based on cpn60 and gyrB sequencing methods. David et al.[9] concluded that the universal target (UT) sequence from the cpn60 gene can be used for phylogenetic and taxonomic studies of the Aeromonas genus. The type I chaperone gene, cpn60[9], and the single-copy protein-coding gene, gyrB[10], are widespread and have been applied in phylogenetic analysis and species identification of Aeromonas[10].
The mechanism underlying the pathogenesis of Aeromonas is multifactorial[11]. Specifically, the pathogenicity is related to virulence factors, including aerolysin (aer), heat-stable cytotoxin (ast), hemolysin (hlyA), lateral flagella (laf), polar flagellum (fla), elastase (ela), lipase (lip), cytotoxic enterotoxin (act), and cytotoxic enterotoxin (alt). The Aeromonas spp. utilize these virulence factors to enable survival within the host, thereby causing extensive cell and tissue destruction, evading the host immune response, and enhancing pathogenic resistance mechanisms, which subsequently help the pathogen to establish an infection[12]. Therefore, further elucidation of the correlation between Aeromonas pathogenicity and the associated virulence factors is warranted.
Owing to the ease-of-culture, low cost, short life cycle, and simple genetic background of Caenorhabditis elegans[13], wild-type C. elegans has been increasingly used as a model to evaluate the virulence of bacteria with pathogenic potential, including Aeromonas spp.[14]. Moreover, Wu et al.[15] reported that the virulence manifested in the cytotoxicity assay is correlated with virulence traits demonstrated in a mouse infection model.
Antibiotics are frequently abused in industries to prevent and control Aeromonas infections, thus resulting in increased antimicrobial resistance[16]. It has been reported that Aeromonas isolates from South Korea were 100% resistant to amoxicillin and nalidixic acid, and 98.5% of the isolates were resistant to ampicillin; all isolates showed multiple antimicrobial resistance phenotypes[17]. Multidrug-resistant (MDR) Aeromonas strains from food products can bypass the acidic conditions of the digestive tract of humans and animals[18]. Hence, antimicrobial resistance of Aeromonas from food items should be strictly monitored to reduce the risk of food-borne Aeromonas infections[19].
In the current study we analyzed the distribution, genetic diversity, antimicrobial resistance, and pathogenicity of Aeromonas isolated from food items to assess the risk of disease in humans and animals. Our study provided the basis for clinical treatment of diseases caused by Aeromonas spp.
-
All isolates were obtained from food products, including fish, shrimp, clams, pigs, chickens, cows, and sheep, collected from six supermarkets in Pudong New District, Shanghai in 2019 (Figure 1). An automatic bacteriologic analyzer (Vitek 2 Compact; BioMèrieux, Marcy I'Etoile, France) was used to identify the Aeromonas isolates[20], which were then cultured at 28 ℃ on brain heart infusion agar plates (Oxoid Ltd., Basingstoke, UK).

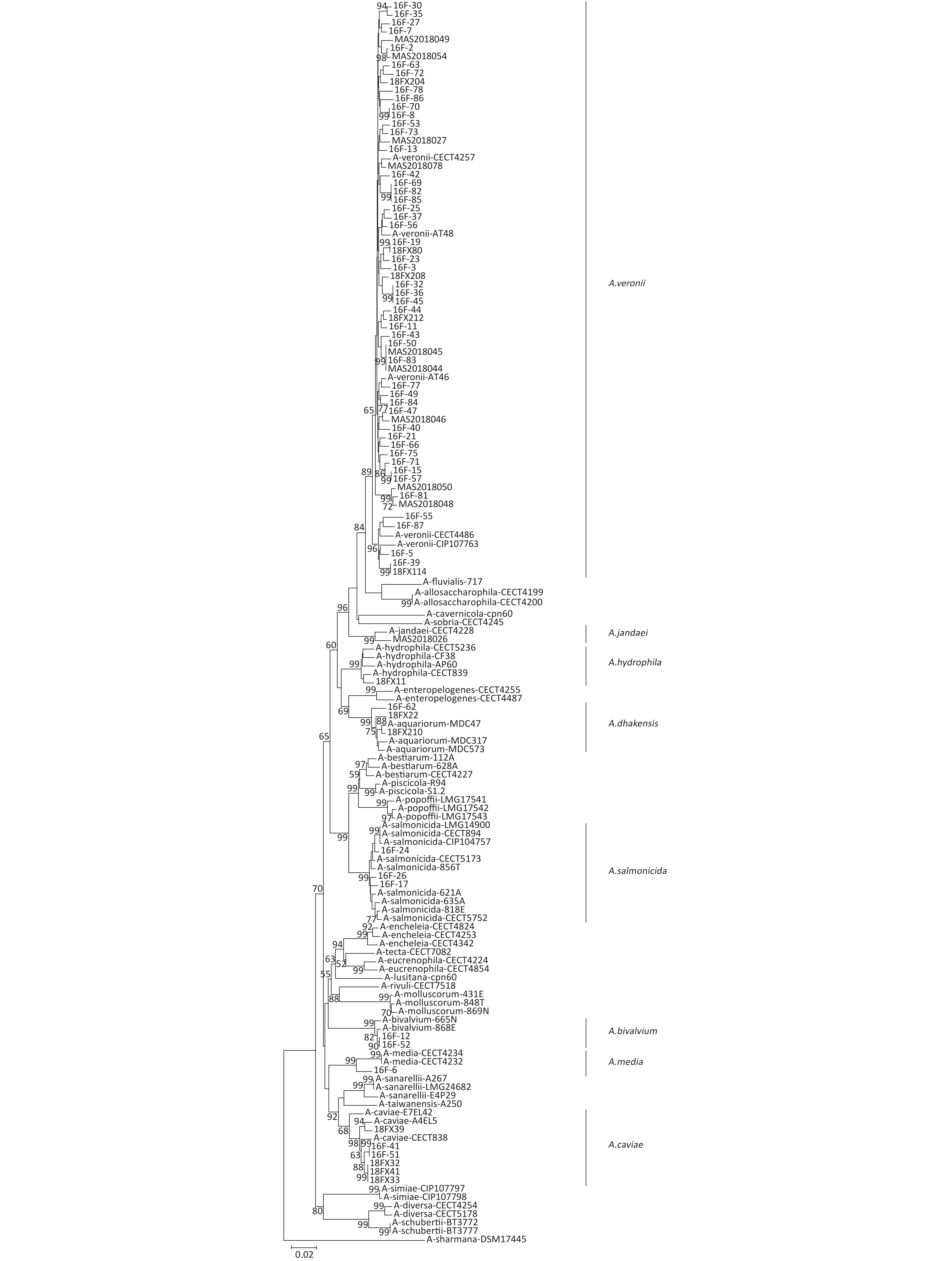
Figure 1. Phylogeny of Aeromonas spp. The concatenated sequences of six housekeeping genes were used to analyze the phylogeny of Aeromonas spp. The information of source, species, virulence genes, and antibiotic resistance are shown. MDR, multidrug-resistance
-
We extracted total chromosomal DNA from the Aeromonas isolates using the QIAamp DNA Mini Kit (Qiagen, Shanghai, China) according to the manufacturer’s instructions. Concatenated gyrB-cpn60 phylogenetic trees were constructed, and we identified the Aeromonas isolates at the species level by comparing two-gene sequences of representative species[21]. The Aeromonas MLST scheme (
http://pubmlst.org/Aeromonas/ ), which relies on the amplification and sequencing of six housekeeping genes (gyrB, groL, gltA, metG, ppsA, and recA), was applied to identify each Aeromonas isolate subtype[22]. PCR was performed using previously described primers and protocols and the sequences of the six loci were compared with the loci hosted on the Aeromonas MLST database and sequence types (STs)[23]. A phylogenetic tree was also constructed using the neighbor-joining method in Clustal-W with bootstrap values calculated using 1,000 replicates. -
The hemolytic ability and extracellular protease activity of the Aeromonas isolates were determined on Luria Bertani (LB) agar plates containing 5% sheep blood (KeMaJia, Shanghai, China) and LB agar plates containing 2% (w/v) skim milk (KeMaJia), respectively. An Aeromonas colony was inoculated on blood and milk agar plates and incubated at 28 ℃[20]. The presence of transparent zones surrounding the Aeromonas colonies were considered positive reactions for both tests.
-
Aeromonas isolates were selected for the C. elegans LT assay[15]. A single colony of Aeromonas was incubated in 45 mL of LB broth at 28 ℃ for 24 h. The Aeromonas suspension was adjusted to an OD600 of 3.0, which was prepared for the toxicity assay. Nematode growth medium (NGM) agar plates with Escherichia coli strain OP50 were used to culture C. elegans. The synchronized adult L4 worms were washed in M9 buffer (
http://www.wormbook.org/ ). After centrifugation, the worms were re-suspended in 5 µL of S-medium (http://www.wormbook.org/ ). Approximately 40–50 worms were added to a 48-well plate with 5 µL of fluorodeoxyuridine to prevent reproduction. Finally, 190 µL of the LB broth with Aeromonas isolates was added to each lawn to a final volume of 200 µL. The plate was incubated at 20 ℃ and the survival rate of worms was observed after 1, 2, and 3 days. -
The mouse C2C12 fibroblast cell line obtained from the American Type Culture Collection (Manassas VA, USA) was used for cytotoxicity assays[20]. The cells were cultured in complete medium with Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS; Gibco) at 37 ℃ in a 5% CO2 incubator for 24 h. The cells were then thrice-washed with DMEM and reacted with Aeromonas spp. for 3 h[15]. The Aeromonas cultures were incubated with cells at a 100 multiplicity of infection (MOI). The CytoTox 96® Assay (Promega, Madison, WI, USA) was used to measure cell death following treatment with a cytotoxic drug or compound[19]. The CytoTox 96® Assay kit was used to determine the release of lactate dehydrogenase (LDH) from the cell after interacting with Aeromonas isolates at 37 ℃ for 3 h. A group treated with 10× lysis solution (Promega) was used as the positive control, and an untreated group with DMEM (Gibco) was the negative control[15]. Cytotoxicity was expressed as the released LDH level induced by Aeromonas isolates compared with that induced by 10× lysis solution (defined as 100% cytotoxicity).
-
Virulence genes (aerA, hlyA, act, ast, alt, ascV, aexT, lip, ela, fla, and laf) were screened as potential toxicity factors of Aeromonas[24]. The PCR system (30 μL) contained 15 μL of Taq PCR MasterMix (Takara Bio, Inc., Kyoto, Japan), 1 μL of primer (10 μmol/L), 11.5 μL of ddH2O, and 1.5 μL of DNA template. The cycling conditions were as follows: pre-denaturation at 96 ℃ for 5 min; 34 cycles of denaturation at 96 ℃ for 30 s, annealing at 55–60 ℃ for 30 s, and extension at 72 ℃ for 1 min and the final extension was at 72 ℃ for 5 min. The PCR products were analyzed using sequencing.
-
Antimicrobial susceptibility testing for Aeromonas spp. was performed following the 2010 Clinical and Laboratory Standard Institute (CLSI) guideline M45-A2[6]. The minimal inhibition concentrations (MICs) of 10 antibiotics [amoxicillin/clavulanic acid (AMC), ampicillin/sulbactam (SAM), cefazolin (CZO), ceftazidime (CAZ), ceftriaxone (CRO), aztreonam (ATM), ciprofloxacin (CIP), levofloxacin (LEV), oxazin/sulfamethoxine oxazole (SXT), and colistin (CT)] were measured. The quality control strains for all antimicrobial susceptibility tests were Escherichia coli ATCC 25922[25].
-
We detected 19 resistance genes (tetA, tetB, tetE; blaTEM, blaSHV, blaCTX; armA, aphAI-IAB, aac(6ʹ)-Ib, aac(3ʹ)-IIa; sul1, sul2; mcr-1, mcr-2, mcr-3, mcr-4; qnrA, qnrB, and qnrS) in the Aeromonas isolates. The primer sequences of the antibiotic resistance genes are shown in Table 1. The PCR-positive products were confirmed using sequencing.
Table 1. Primer sequences used to amplify antimicrobial resistance genes
Targeted genes Primers Sequences (5′→3′) Product size (bp) Extended-spectrum beta-lactamase blaTEM blaTEM-F ATAAAATTCTTGAAGACGAAA 1,080 blaTEM-R GACAGTTACCAATGCTTAATC blaSHV blaSHV-F TTATCTCCCTGTTAGCCACC 795 blaSHV-R GATTTGCTGATTTCGCTCGG blaCTX-M blaCTX-M-F CGCTTTGCGATGTGCAG 550 blaCTX-M-R ACCGCGATATCGTTGGT Tetracycline resistance tetA tetA-F GTAATTCTGAGCACTGTCGC 1,000 tetA-R CTGCCTGGACAACATTGCTT tetB tetB-F CTCAGTATTCCAAGCCTTTG 400 tetB-R CTAAGCACTTGTCTCCTGTT tetE tetE-F GTGATGATGGCACTGGTCAT 1,100 tetE-R CTCTGCTGTACATCGCTCTT Plasmids mediate quinolone resistance qnrA qnrA-F AGAGGATTTCTCACGCCAGG 580 qnrA-R TGCCAGGCACAGATCTTGAC qnrB qnrB-F GATCGTGAAAGCCAGAAAGG 496 qnrB-R ACGATGCCTGGTAGTTGTCC qnrS qnrS-F GCAAGTTCATTGAACAGGGT 428 qnrS-R TCTAAACCGTCGAGTTCGGCG Aminoglycoside resistance armA armA-F AGGTTGTTTCCATTTCTGAG 591 armA-R TCTCTTCCATTCCCTTCTCC aphAI-IAB aphAI-IAB-F AAACGTCTTGCTCGA GGC 500 aphAI-IAB-R CAAACCGTTATTCATTCGTGA aac(3)-IIa aac(3)-IIa-F ATGGGCATC ATTCGCACA 749 aac(3)-IIa-R TCTCGGCTTGAACGAATTGT aac(6’)-Ib aac(6’)-Ib-F TTGCGATGCTCTATGAGTGGCTA 482 aac(6’)-Ib-R CTCGAATGCCTGGCGTGTTT Mobile colistin resistance mcr-1 mcr-1-F CGGTCAGTCCGTTTGTTC 309 mcr-2-R CTTGGTCGGTCTGTAGGG mcr-2 mcr-2-F TGTTGCTTGTGCCGATTGGA 567 mcr-2-R CAGCAACCAACAATACCATCT mcr-3 mcr-3-F AGTTTGGTTTCGCCATTTCATTAC 1,084 mcr-3-R ATATCACTGCGTGGACAGTCAGG mcr-4 mcr-4-F TTACAGCCAGAATCATTATCA 488 mcr-4-R ATTGGGATAGTCGCCTTTTT Sulfonamide resistance sul1 sul1-F CGGCGTGGGCTACCTGAACG 433 sul1-R GCCGATCGCGTGAAGTTCCG sul2 sul2-F GCGCTCAAGGCAGATGGCATT 293 sul2-R GCGTTTGATACCGGCACCCGT -
Data analysis was performed using SPSS software (version 23.0 for Windows; IBM Corp, Armonk, NY, USA). The 95% CIs of categorical variables for the distribution of virulence genes and the results of the C. elegans LT assay were calculated using Fisher’s exact tests. The cytotoxicity test data were analyzed using a nonparametric test to reveal the toxicity differences among different Aeromonas isolates.
-
The 79 isolates of Aeromonas were categorized into 73 STs, 65 of which were novel (ST889-ST961 and ST991-ST1023), revealing a high degree of genetic diversity among the Aeromonas strains (Figure 1). There was no dominant ST observed in any of the Aeromonas isolates.
-
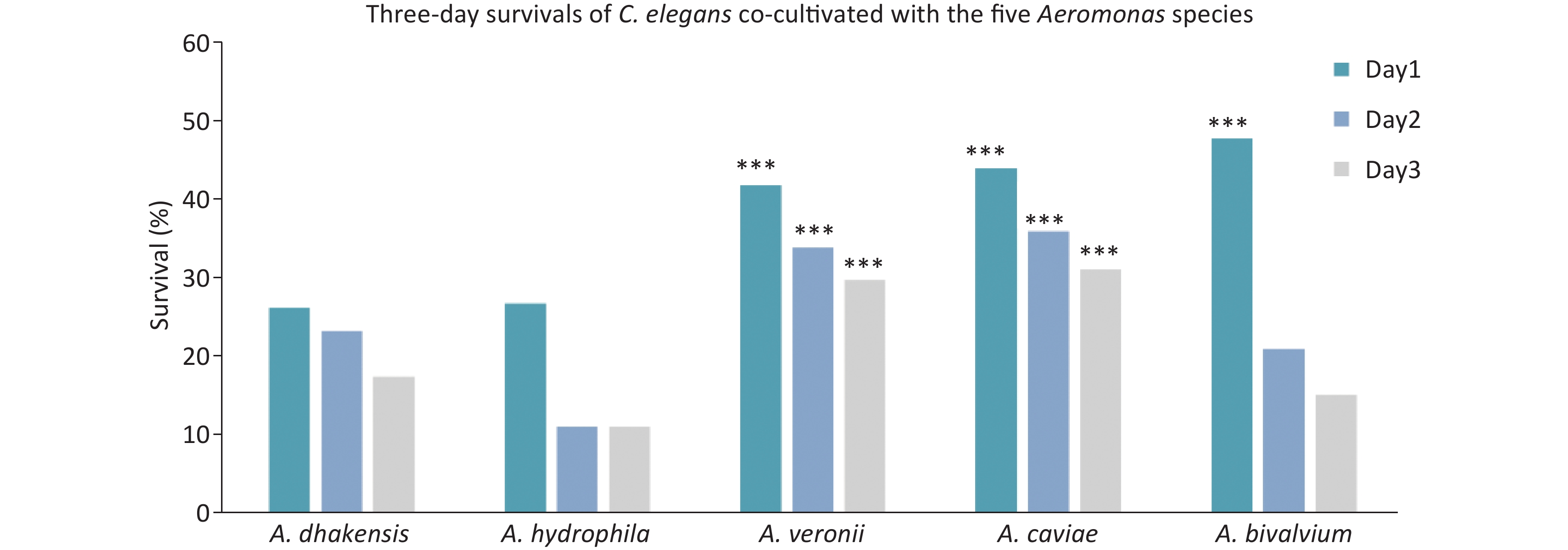
We identified all the Aeromonas isolates based on gyrB-cpn60 sequencing, which were distributed as follows: A. veronii (78.5%), A. caviae (7.6%), A. dhakensis (3.8%), A. salmonicida (3.8%), A. bivalvium (2.5%), A. hydrophila (1.3%), A. jandaei (1.3%), and A. media (1.3%), Figure 2. In addition, the Aeromonas spp. were isolated from a variety of food products, of which the four most frequent foodstuffs were fish, pork, chicken, and spiral shell (Figure 1).
Figure 2. Subtyping of Aeromonas isolates. A neighbor-joining phylogenetic tree was constructed using the concatenated sequences of gyrB and cpn60 to reveal the relationships between 79 Aeromonas isolates from food products.
-
From the 79 Aeromonas isolates, 36 (45.6%) exhibited β-hemolytic activity and 64 (81%) exhibited proteolytic activity (Table 2). A. caviae lacked hemolytic activity; the positive rate of proteolytic activity was 16.7%. A. dhakensis (66.7% and 100%, respectively) and A. hydrophila (100% and 100%, respectively) had more hemolytic and exoprotease activities than A. veronii (50.0% and 85.5%, respectively), A. caviae (0 and 16.7%, respectively) and A. bivalvium (0 and 100%, respectively). Moreover, the positive rate of hemolytic and proteolytic activities in A. salmonicida strains were 33.3% and 66.7%, respectively. A. jandaei showed no signs of β-hemolysis.
Table 2. Prevalence of hemolytic and proteolytic activities in Aeromonas isolates
Assay A. veronii,
n (%)A. caviae,
n (%)A. dhakensis,
n (%)A. hydrophila,
n (%)A. salmonicida,
n (%)A. bivalvium,
n (%)A. jandaei,
n (%)A. media,
n (%)Hemolysis 31 (50.0) 0 2 (66.7) 1 (100) 1 (33.3) 0 0 1 (100) Proteolysis 53 (85.5) 1 (16.7) 3 (100) 1 (100) 2 (66.7) 2 (100) 1 (100) 1 (100) Total 62 6 3 1 3 2 1 1 -
The pathogenicity of Aeromonas isolates was assessed using the C. elegans LT test, and the results are shown in Figure 3. The 1, 2, and 3-day survival rates of C. elegans fed with A. dhakensis, A. hydrophila, A. veronii, A. caviae, and A. bivalvium were as follows: 26.1%, 23.2%, and 17.4%; 26.7%, 11.1%, and 11.1%; 41.7%, 33.8%, and 29.6%; 43.8%, 35.8%, and 31.0%; and 47.7%, 20.9%, and 15.1%, respectively. Thus, A. dhakensis and A. hydrophila were more pathogenic than other Aeromonas spp. Overall, the C. elegans survival rates when cultured with A. dhakensis and A. hydrophila were significantly lower than A. veronii, A. caviae, and A. bivalvium (P < 0.05).
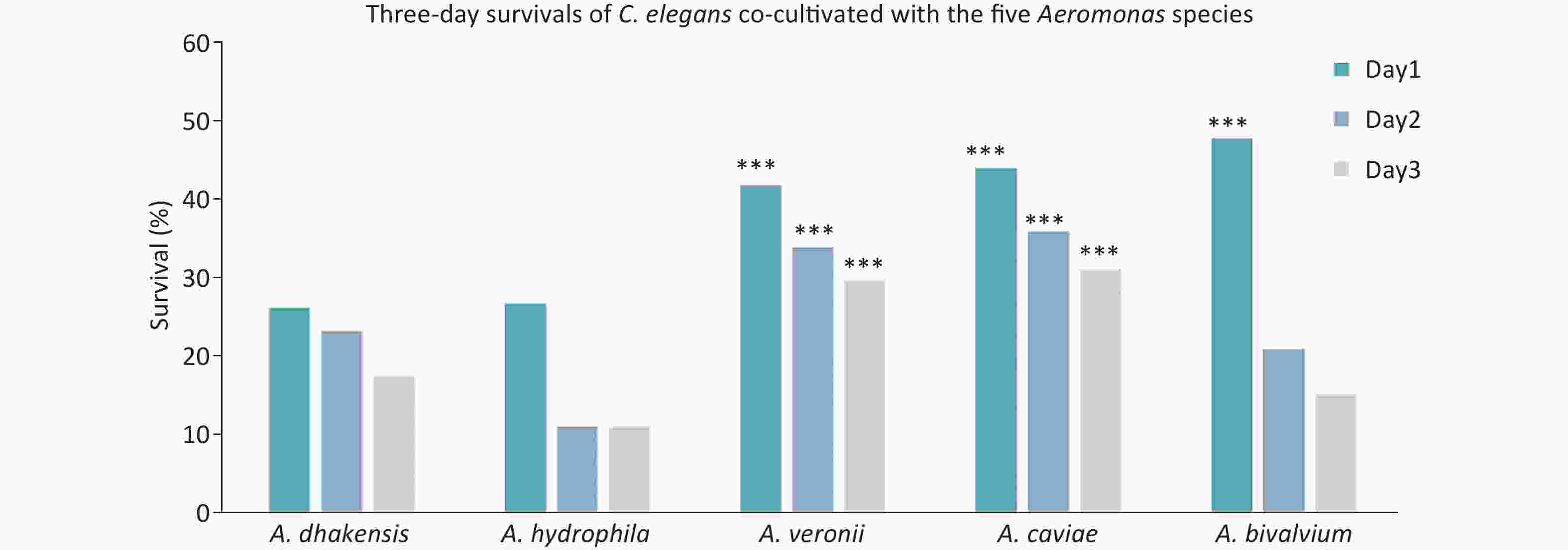
Figure 3. C. elegans liquid toxicity assay. Three-day survival of C. elegans co-cultivated with the following five Aeromonas species: A. dhakensis; A. hydrophila; A. veronii; A. caviae; and A. bivalvium in the liquid-toxic assay. ***P < 0.05, as compared with A. dhakensis
The pathogenicity of Aeromonas isolates was assessed using cytotoxicity tests. Compared with LDH release of the 10× lysis solution, the volume of LDH release induced by A. dhakensis, A. hydrophila, A. veronii, A. caviae, and A. bivalvium were 75.9%, 74.5%, 46.4%, 11.9%, and 5.2%, respectively. Our results indicated that A. dhakensis and A. hydrophila were more virulent than A. veronii, A. caviae, and A. bivalvium, which was in agreement with the results of the C. elegans LT assay. The LDH release level varied between Aeromonas spp. (P < 0.05).
-
The distribution of the 11 virulence genes in the Aeromonas strains is summarized in detail (Table 3). In particular, act and fla were the two most frequent virulence genes among all the isolates, accounting for 73.4% and 69.6%, respectively. The ast and aerA genes were detected in < 10% of all isolates. The detection rates of lip, ela, act, alt, aerA, and hlyA were significantly different among the eight species of Aeromonas [P < 0.05 (Fisher’s exact test)]. Apart from the three genes (ast, aexT, and ascV), all other virulence genes were detected in A. dhakensis. Only act, fla, laf, aexT, and ascV were more prevalent in A. veronii than other Aeromonas spp.
Table 3. Distribution of putative virulence factors in all Aeromonas isolates
Isolate ast, n (%) lip, n (%) ela, n (%) act, n (%) alt, n (%) aerA, n (%) hlyA, n (%) fla, n (%) laf, n (%) ascV, n (%) aexT, n (%) n = 2 n = 18 n = 21 n = 58 n = 11 n = 6 n = 8 n = 55 n = 11 n = 24 n = 29 A. veronii 1 (1.6) 4 (6.5) 8 (13) 53 (85.5) 1 (1.6) 2 (3.2) 2 (3.2) 45 (72.6) 7 (11.3) 22 (35.5) 26 (41.9) A. caviae 0 6 (100) 5 (83.3) 0 2 (33.3) 0 0 4 (66.7) 3 (50) 0 1 (16.7) A. dhakensis 0 3 (100) 2 (66.7) 2 (66.7) 3 (100) 2 (66.7) 3 (100) 2 (66.7) 1 (33.3) 0 0 A. hydrophila 1 (100) 1 (100) 1 (100) 0 0 0 1 (100) 0 0 1 (100) 0 A. salmonicida 0 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 0 1 (33.3) 1 (33.3) A. bivalvium 0 2 (100) 2 (100) 0 2 (100) 0 0 2 (100) 0 0 0 A. jandaei 0 0 0 1 (100) 1 (100) 0 0 0 0 0 1 (100) A. media 0 0 1 (100) 0 0 0 0 0 0 0 0 P value 0.105 < 0.001 < 0.001 < 0.001 < 0.001 0.003 < 0.001 0.310 0.215 0.155 0.486 In addition to three genes (fla, ascV, and aexT), the prevalence of other genes differed significantly among the five Aeromonas spp. [P < 0.05 (Fisher’s exact test); Table 4]. Only hlyA was more frequent in A. dhakensis and A. hydrophila than A. veronii, A. caviae, and A. bivalvium. The genes, act and aerA, were abundant in A. dhakensis and A.veronii, but neither was detected in the other three Aeromonas spp. The gene, alt, was most commonly identified in A. dhakensis, A. hydrophila, and A. bivalvium. Moreover, the genes, lip and ela, were less prevalent in A.veronii than A.dhakensis, A. hydrophila, A. caviae, and A. bivalvium.
Table 4. Distribution of virulence factors among isolates of A. dhakensis, A. hydrophila, A. veronii A. caviae, and A. bivalvium
Gene A. dhakensis, n (%) A. hydrophila, n (%) A. veronii, n (%) A. caviae, n (%) A. bivalvium, n (%) P value n = 3 n = 1 n = 62 n = 6 n = 2 ast 0 1 (100) 1 (1.6) 0 0 0.047 lip 3 (100) 1 (100) 4 (6.5) 6 (100) 2 (100) < 0.001 ela 2 (66.7) 1 (100) 8 (13) 5 (83.3) 2 (100) < 0.001 act 2 (66.7) 0 53 (85.5) 0 0 < 0.001 alt 3 (100) 1 (100) 1 (1.6) 2 (33.3) 2 (100) < 0.001 aerA 2 (66.7) 0 2 (3.2) 0 0 0.027 hlyA 3 (100) 1 (100) 2 (3.2) 0 0 < 0.001 fla 2 (66.7) 0 45 (72.6) 4 (66.7) 2 (100) 0.603 laf 1 (33.3) 1 (100) 7 (11.3) 3 (50) 0 0.020 ascV 0 1 (100) 22 (35.5) 0 0 0.121 aexT 0 0 26 (41.9) 1 (16.7) 0 0.360 -
Resistance to CZO was demonstrated in 74 (93.7%) Aeromonas isolates. The antibiotic resistance rates of A. veronii to AMC, CRO, CIP, LEV, trimethoprim-sulfamethoxazole, and CT were 1.6%, 3.2%, 4.8%, 1.6%, 9.7%, and 1.6%, respectively. A. veronii was susceptible to SAM, CAZ, and ATM. A. dhakensis showed 100% resistance to CT and CZO, and 33.3% to AMC and CRO. A. caviae was shown to be 100% resistant to CZO and 66.7% against CAZ, CRO, and ATM. In addition to CZO, A. bivalvium was susceptible to other antibiotics. The antimicrobial resistance rates of Aeromonas isolates against 10 common antibiotics are presented in (Table 5).
Table 5. Prevalence of resistance to different antibiotics
Drug Resistance isolates, n (%) A. veronii A. caviae A. dhakensis A. hydrophila A. salmonicida A. bivalvium A. jandaei A. media n = 62 n = 6 n = 3 n = 1 n = 3 n = 2 n = 1 n = 1 Penicillins Ampicillin/sulbactam 0 0 0 0 0 0 0 1 (100) Amoxicillin/clavulanic acid 1 (1.6) 0 1 (33.3) 0 1 (33.3) 0 1 (100) 0 Cephems Cefazolin 58 (93.5) 6 (100) 3 (100) 1 (100) 3 (100) 1 (50) 1 (100) 1 (100) Ceftazidime 0 4 (66.7) 0 0 0 0 0 0 Ceftriaxone 2 (3.2) 4 (66.7) 1 (33.3) 0 0 0 1 (100) 0 Monobactams Aztreonam 0 4 (66.7) 0 0 0 0 0 0 Quinolones Ciprofloxacin 3 (4.8) 0 0 1 (100) 0 0 0 0 Levofloxacin 1 (1.6) 0 0 0 0 0 0 0 Folate pathway inhibitors Trimethoprim-sulfamethoxazole 6 (9.7) 0 0 1 (100) 0 0 0 0 Polymyxins Colistin 1 (1.6) 0 3 (100) 1 (100) 0 0 1 (100) 0 In this study, 9 (11.4%) Aeromonas isolates were multidrug-resistant, and the high MDR rates of A. dhakensis, A. hydrophila, A. veronii, and A. caviae were 33.3%, 100%, 3.2%, and 66.7%, respectively, which is an enormous challenge for treatment options.
-
The tetracycline resistance gene, tetE, was found in A. veronii (35.5%), A. caviae (33.3%), A. salmonicida (66.7%), and A. media (100%). The aminoglycoside resistance gene, aac(6ʹ)-Ib, was detected in A. veronii (16.1%), A. caviae (50.0%), A. bivalvium (50.0%), A. dhakensis (33.3%), A. media (100%), and A. hydrophila (100%). Moreover, the CT resistance gene, mcr-3, accounted for 16.1% of A. veronii and 33.3% of A. dhakensis. The PMQR gene, qnrS, was present in 3.2% of A. veronii and 100% of A. hydrophila. In addition, blaTEM (1.6%), tetA (1.6%), and mcr-1(1.6%) were present in A. veronii. No drug resistance gene was observed in A. jandaei. The following genes were not detected in any Aeromonas isolates: the ESBL genes, blaSHV and blaCTX-M; the tetracycline resistance gene, tetB; the PMQR genes, qnrA and qnrB; the aminoglycoside resistance genes, aphAI-IAB, aac (3ʹ)-IIa, and armA; the CT resistance genes, mcr-2 and mcr-4; and the sulphonamide resistance genes, sul1 and sul2 (Figure 1).
-
Common clinical manifestations of Aeromonas infections include gastrointestinal infections, diarrhea, bacteremia, and localized soft tissue infections[11]. The clinical Aeromonas isolates (96.5%) that have been identified to date belong to one of the following species: A. caviae (29.9%); A. dhakensis (26.3%); A. veronii (24.8%); and A. hydrophila (15.5%)[26]. Escarpulli et al.[27] detected 82 species of Aeromonas in 250 frozen edible fish in Mexico City, Mexico and emphasized the risks associated with the consumption of the Aeromonas isolated from frozen edible fish. In our study, 29 (36.7%) strains of Aeromonas were isolated from fish samples. Consumption of food infected with Aeromonas poses a potential threat to human health. Accordingly, we analyzed Aeromonas isolates obtained from food samples from Shanghai Agricultural Market and evaluated the species distribution, antibiotic resistance, and pathogenicity.
Among the 73 STs identified from the 79 Aeromonas isolates, 65 were novel, which indicated that the isolates obtained from food products had high genetic diversity. Lau et al.[28] also found 36 STs in 47 isolates in Malaysia, 34 of which were novel STs, which suggested that the isolates obtained from food products often had high genetic diversity. The phylogenetic relationship of 79 Aeromonas isolates was evaluated using the housekeeping gene, gyrB-cpn60[28]. A. veronii was the most dominant Aeromonas species, comprising 78.5% of all isolates, and was isolated from meat (43.5%) and fish (33.9%). A. caviae (7.6%) was the second most prevalent Aeromonas species and was distributed in various foods, such as fish and calms. Elala et al.[29] isolated A. caviae from chickens, suggesting that this species could survive in both aquatic and terrestrial environments. The third most abundant species was A. dhakensis, which has been reported[5] to have a higher virulence potential and is a notable human pathogen. In our study, A. dhakensis was isolated from fish and shrimp samples at a 2:1 ratio. Similarly, Wu et al.[15] reported bacteria isolated from fish and clinical isolates in Tainan to be Aeromonas spp.; the most prevalent species were A. caviae, A. dhakensis, and A. veronii.
In this study, we analyzed the virulence of Aeromonas isolates using β-hemolysis and proteolytic, C. elegans LT, and cytotoxicity assays. The positivity rates of A. dhakensis and A. hydrophila based on the β-hemolysis and proteolytic assays were higher than A. veronii, A. caviae, and A. bivalvium, which was confirmed by the survival rate of C. elegans fed with Aeromonas isolates, as well as the LDH release from mouse fibroblasts C2C12 infected with Aeromonas spp. Wu et al.[15] reported that the pathogenicity of A. veronii and A. caviae were less virulent than A. dhakensis and A. hydrophila, which was consistent with our findings. The virulence of A. dhakensis against C. elegans was shown to be more evident than that of A. hydrophila[20], in contrast to the pathogenicity of A. dhakensis, which differed little from A. hydrophila in our investigation.
Cody et al.[30] reported that the virulence phenotype of Aeromonas was the result of a combined effect of its virulence factors. We detected several virulence genes from the Aeromonas spp. In all Aeromonas isolates, act was the most prevalent virulence gene, followed by fla, aexT, and ascV. The cytolytic enterotoxin gene, act, was the most frequently detected among all isolates, and the result corresponded with the data of act (63%) and alt (57%), which were higher than ast (6%) in Aeromonas from water and fish sourced in India[31]. Furthermore, act was mainly distributed in A. veronii (85.5%), and fla was observed in most Aeromonas spp. except A. jandaei, A. hydrophila, and A. media. The aexT gene was present in all A. jandaei, 41.9% of A. veronii, 33.3% of A. salmonicida, and 16.7% of A. caviae. Another study[15] suggested that compared to other Aeromonas pathogens, the lack of aerA or hlyA may be associated with low virulence of A. caviae and A. bivalvium. These results represented the distribution of virulence genes in food products and indicated the potential pathogenicity of Aeromonas isolates, which could help enhance food quality and hygiene in this region, and aid in preventing and controlling the outbreak of Aeromonas-related diseases.
The results of this study indicated that Aeromonas isolates displayed 93.7% resistance to the first-generation cephalosporin (CZO) and < 11% resistance to third-generation cephalosporins (10.1% for CRO and 5% for CAZ). Several studies[32, 33] reported that the majority of isolates had 100% susceptibility to third-generation cephalosporins (CAZ and cefotaxime), while there was a different degree of resistance to first-generation cephalosporins, ranging from 79.3% to 91.8%. Generally, our findings were in agreement with other studies[32, 33]. In addition, A. bivalvium had resistance to CZO (50%), and this finding corroborated the finding that all isolates from clinical and environmental samples were 80% resistant to CZO[33], which indicates that resistance to first-generation cephalosporins could be prevalent[34]. We found that isolates of A. dhakensis exhibited resistance to CZO (100%), CT (100%), CRO (33.3%), and AMC (33.3%), confirming multiple drug resistance of this species, which is in agreement with previous findings[6]. This study demonstrated that A. veronii had various types of drug resistance, with 100% susceptibility to SAM, CAZ, and ATM. Absolute (100%) sensitivity to CIP was reported in a previous study in which several A. veronii strains were isolated from clinical patients[33]; however, this differed from the CIP resistance rate of 5% determined in the current study. Moreover, we found that the resistance rate of Aeromonas spp. to CT was 7.6%, which was approximately the same as that of Aeromonas spp. isolated from Australian fish. CT is the last line of antibiotic treatment used for clinically severe infections caused by multidrug-resistant gram-negative bacteria[33]. In the present study, nine strains of Aeromonas showed a multidrug resistance rate of 11.4%, which was lower than that reported in a previous study[6].
Consistent with the findings of a study conducted by Dahanayake et al.[35], tetE was detected in 27 strains (34.2%) and tetA was detected in 1 strain (1.3%); However, tetB was not detected in the present study. Moreover, the tetE detection rates varied among A. veronii, A. caviae, A. media, and A. salmonicida, ranging from 33.3% to 100%. Recently, aminoglycosides have been extensively used for treating severe infections caused by Aeromonas, and Dahanayake et al.[35] reported the presence of aac(6ʹ)-Ib in all Aeromonas spp. isolated from cockles in Korea. In the present study, 21.5% of the isolates of Aeromonas spp. harbored aac(6ʹ)-Ib, except for A. salmonicida and A. jandaei.
Our study indicated that approximately 13.9% of Aeromonas spp. harbored mcr-3 (16.1% of A. veronii and 33.3% of A. dhakensis). These data support the discovery of mcr-3 in A. jandaei isolated from fish sold in Beijing, China[36]. Studies have reported the presence of qnrS not only in Enterobacteriaceae, but also in Aeromonas, suggesting that this gene is distributed across different bacterial species[37]. We found that Aeromonas strains collected from food products had a resistance rate of 8.9% to SXT, and the same strains exhibited a detection rate of 3.8% for qnrS. This gene was detected in 3.2% and 100% of A. veronii and A. hydrophila isolates, respectively. Previous studies have reported different rates of detection of qnrS in Lake Lugano (Switzerland)[37] and China[6]. Moreover, the rate of detection of blaTEM in A. veronii was 1.6%, confirming the presence of blaTEM in A. veronii isolated from German water sources[38]. Nonetheless, these findings should be further assessed in future studies.
-
Aeromonas was an important zoonotic pathogen that is mainly distributed in fish and other aquatic products. Studies have demonstrated that these isolates had a high degree of genetic diversity; the 79 isolates in the current study were shown to produce 73 independent STs. The isolates were divided into eight species by gyrB-cpn60 sequencing. The distribution, taxonomy, drug resistance, and pathogenicity varied among different Aeromonas spp. The percentage of Aeromonas isolates shown to be MDR was 11.4%. Food-derived Aeromonas isolates (93.7%) were resistant to CZO. Aeromonas carried some virulence and antibiotic resistance genes, which posed a threat and challenge to food safety and clinical treatment monitoring. A few isolates, such as A. dhakensis and A. hydrophila, had high pathogenicity and could cause extensive infections and posed risks to human and animal health.
-
The authors have declared that no competing interests exist.
doi: 10.3967/bes2022.108
Genetic Diversity, Antibiotic Resistance, and Pathogenicity of Aeromonas Species from Food Products in Shanghai, China
-
Abstract:
Objective Aeromonas has recently been recognized as an emerging human pathogen. Aeromonas-associated diarrhea is a phenomenon occurring worldwide. This study was designed to determine the prevalence, genetic diversity, antibiotic resistance, and pathogenicity of Aeromonas strains isolated from food products in Shanghai. Methods Aeromonas isolates (n = 79) collected from food samples were analyzed using concatenated gyrB-cpn60 sequencing. The antibiotic resistance of these isolates was determined using antimicrobial susceptibility testing. Pathogenicity was assessed using β-hemolytic, extracellular protease, virulence gene detection, C. elegans liquid toxicity (LT), and cytotoxicity assays. Results Eight different species were identified among the 79 isolates. The most prevalent Aeromonas species were A. veronii [62 (78.5%)], A. caviae [6 (7.6%)], A. dhakensis [3 (3.8%)], and A. salmonicida [3 (3.8%)]. The Aeromonas isolates were divided into 73 sequence types (STs), of which 65 were novel. The isolates were hemolytic (45.6%) and protease-positive (81.0%). The most prevalent virulence genes were act (73.4%), fla (69.6%), aexT (36.7%), and ascV (30.4%). The results of C. elegans LT and cytotoxicity assays revealed that A. dhakensis and A. hydrophila were more virulent than A. veronii, A. caviae, and A. bivalvium. Antibiotic resistance genes [tetE, blaTEM, tetA, qnrS, aac(6)-Ib, mcr-1, and mcr-3] were detected in the isolates. The multidrug-resistance rate of the Aeromonas isolates was 11.4%, and 93.7% of the Aeromonas isolates were resistant to cefazolin. Conclusion The taxonomy, antibiotic resistance, and pathogenicity of different Aeromonas species varied. The Aeromonas isolates A. dhakensis and A. hydrophila were highly pathogenic, indicating that food-derived Aeromonas isolates are potential risks for public health and food safety. The monitoring of food quality and safety will result in better prevention and treatment strategies to control diarrhea illnesses in China. -
Key words:
- Aeromonas /
- Genetic diversity /
- Antibiotic resistance /
- Virulence gene /
- Cytotoxicity assay /
- Multidrug-resistance
注释: -
Figure 1. Phylogeny of Aeromonas spp. The concatenated sequences of six housekeeping genes were used to analyze the phylogeny of Aeromonas spp. The information of source, species, virulence genes, and antibiotic resistance are shown. MDR, multidrug-resistance
Figure 2. Subtyping of Aeromonas isolates. A neighbor-joining phylogenetic tree was constructed using the concatenated sequences of gyrB and cpn60 to reveal the relationships between 79 Aeromonas isolates from food products.
Figure 3. C. elegans liquid toxicity assay. Three-day survival of C. elegans co-cultivated with the following five Aeromonas species: A. dhakensis; A. hydrophila; A. veronii; A. caviae; and A. bivalvium in the liquid-toxic assay. ***P < 0.05, as compared with A. dhakensis
Table 1. Primer sequences used to amplify antimicrobial resistance genes
Targeted genes Primers Sequences (5′→3′) Product size (bp) Extended-spectrum beta-lactamase blaTEM blaTEM-F ATAAAATTCTTGAAGACGAAA 1,080 blaTEM-R GACAGTTACCAATGCTTAATC blaSHV blaSHV-F TTATCTCCCTGTTAGCCACC 795 blaSHV-R GATTTGCTGATTTCGCTCGG blaCTX-M blaCTX-M-F CGCTTTGCGATGTGCAG 550 blaCTX-M-R ACCGCGATATCGTTGGT Tetracycline resistance tetA tetA-F GTAATTCTGAGCACTGTCGC 1,000 tetA-R CTGCCTGGACAACATTGCTT tetB tetB-F CTCAGTATTCCAAGCCTTTG 400 tetB-R CTAAGCACTTGTCTCCTGTT tetE tetE-F GTGATGATGGCACTGGTCAT 1,100 tetE-R CTCTGCTGTACATCGCTCTT Plasmids mediate quinolone resistance qnrA qnrA-F AGAGGATTTCTCACGCCAGG 580 qnrA-R TGCCAGGCACAGATCTTGAC qnrB qnrB-F GATCGTGAAAGCCAGAAAGG 496 qnrB-R ACGATGCCTGGTAGTTGTCC qnrS qnrS-F GCAAGTTCATTGAACAGGGT 428 qnrS-R TCTAAACCGTCGAGTTCGGCG Aminoglycoside resistance armA armA-F AGGTTGTTTCCATTTCTGAG 591 armA-R TCTCTTCCATTCCCTTCTCC aphAI-IAB aphAI-IAB-F AAACGTCTTGCTCGA GGC 500 aphAI-IAB-R CAAACCGTTATTCATTCGTGA aac(3)-IIa aac(3)-IIa-F ATGGGCATC ATTCGCACA 749 aac(3)-IIa-R TCTCGGCTTGAACGAATTGT aac(6’)-Ib aac(6’)-Ib-F TTGCGATGCTCTATGAGTGGCTA 482 aac(6’)-Ib-R CTCGAATGCCTGGCGTGTTT Mobile colistin resistance mcr-1 mcr-1-F CGGTCAGTCCGTTTGTTC 309 mcr-2-R CTTGGTCGGTCTGTAGGG mcr-2 mcr-2-F TGTTGCTTGTGCCGATTGGA 567 mcr-2-R CAGCAACCAACAATACCATCT mcr-3 mcr-3-F AGTTTGGTTTCGCCATTTCATTAC 1,084 mcr-3-R ATATCACTGCGTGGACAGTCAGG mcr-4 mcr-4-F TTACAGCCAGAATCATTATCA 488 mcr-4-R ATTGGGATAGTCGCCTTTTT Sulfonamide resistance sul1 sul1-F CGGCGTGGGCTACCTGAACG 433 sul1-R GCCGATCGCGTGAAGTTCCG sul2 sul2-F GCGCTCAAGGCAGATGGCATT 293 sul2-R GCGTTTGATACCGGCACCCGT  下载: 导出CSV
下载: 导出CSV
Table 2. Prevalence of hemolytic and proteolytic activities in Aeromonas isolates
Assay A. veronii,
n (%)A. caviae,
n (%)A. dhakensis,
n (%)A. hydrophila,
n (%)A. salmonicida,
n (%)A. bivalvium,
n (%)A. jandaei,
n (%)A. media,
n (%)Hemolysis 31 (50.0) 0 2 (66.7) 1 (100) 1 (33.3) 0 0 1 (100) Proteolysis 53 (85.5) 1 (16.7) 3 (100) 1 (100) 2 (66.7) 2 (100) 1 (100) 1 (100) Total 62 6 3 1 3 2 1 1
下载: 导出CSV
Table 3. Distribution of putative virulence factors in all Aeromonas isolates
Isolate ast, n (%) lip, n (%) ela, n (%) act, n (%) alt, n (%) aerA, n (%) hlyA, n (%) fla, n (%) laf, n (%) ascV, n (%) aexT, n (%) n = 2 n = 18 n = 21 n = 58 n = 11 n = 6 n = 8 n = 55 n = 11 n = 24 n = 29 A. veronii 1 (1.6) 4 (6.5) 8 (13) 53 (85.5) 1 (1.6) 2 (3.2) 2 (3.2) 45 (72.6) 7 (11.3) 22 (35.5) 26 (41.9) A. caviae 0 6 (100) 5 (83.3) 0 2 (33.3) 0 0 4 (66.7) 3 (50) 0 1 (16.7) A. dhakensis 0 3 (100) 2 (66.7) 2 (66.7) 3 (100) 2 (66.7) 3 (100) 2 (66.7) 1 (33.3) 0 0 A. hydrophila 1 (100) 1 (100) 1 (100) 0 0 0 1 (100) 0 0 1 (100) 0 A. salmonicida 0 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 2 (66.7) 0 1 (33.3) 1 (33.3) A. bivalvium 0 2 (100) 2 (100) 0 2 (100) 0 0 2 (100) 0 0 0 A. jandaei 0 0 0 1 (100) 1 (100) 0 0 0 0 0 1 (100) A. media 0 0 1 (100) 0 0 0 0 0 0 0 0 P value 0.105 < 0.001 < 0.001 < 0.001 < 0.001 0.003 < 0.001 0.310 0.215 0.155 0.486
下载: 导出CSV
Table 4. Distribution of virulence factors among isolates of A. dhakensis, A. hydrophila, A. veronii A. caviae, and A. bivalvium
Gene A. dhakensis, n (%) A. hydrophila, n (%) A. veronii, n (%) A. caviae, n (%) A. bivalvium, n (%) P value n = 3 n = 1 n = 62 n = 6 n = 2 ast 0 1 (100) 1 (1.6) 0 0 0.047 lip 3 (100) 1 (100) 4 (6.5) 6 (100) 2 (100) < 0.001 ela 2 (66.7) 1 (100) 8 (13) 5 (83.3) 2 (100) < 0.001 act 2 (66.7) 0 53 (85.5) 0 0 < 0.001 alt 3 (100) 1 (100) 1 (1.6) 2 (33.3) 2 (100) < 0.001 aerA 2 (66.7) 0 2 (3.2) 0 0 0.027 hlyA 3 (100) 1 (100) 2 (3.2) 0 0 < 0.001 fla 2 (66.7) 0 45 (72.6) 4 (66.7) 2 (100) 0.603 laf 1 (33.3) 1 (100) 7 (11.3) 3 (50) 0 0.020 ascV 0 1 (100) 22 (35.5) 0 0 0.121 aexT 0 0 26 (41.9) 1 (16.7) 0 0.360
下载: 导出CSV
Table 5. Prevalence of resistance to different antibiotics
Drug Resistance isolates, n (%) A. veronii A. caviae A. dhakensis A. hydrophila A. salmonicida A. bivalvium A. jandaei A. media n = 62 n = 6 n = 3 n = 1 n = 3 n = 2 n = 1 n = 1 Penicillins Ampicillin/sulbactam 0 0 0 0 0 0 0 1 (100) Amoxicillin/clavulanic acid 1 (1.6) 0 1 (33.3) 0 1 (33.3) 0 1 (100) 0 Cephems Cefazolin 58 (93.5) 6 (100) 3 (100) 1 (100) 3 (100) 1 (50) 1 (100) 1 (100) Ceftazidime 0 4 (66.7) 0 0 0 0 0 0 Ceftriaxone 2 (3.2) 4 (66.7) 1 (33.3) 0 0 0 1 (100) 0 Monobactams Aztreonam 0 4 (66.7) 0 0 0 0 0 0 Quinolones Ciprofloxacin 3 (4.8) 0 0 1 (100) 0 0 0 0 Levofloxacin 1 (1.6) 0 0 0 0 0 0 0 Folate pathway inhibitors Trimethoprim-sulfamethoxazole 6 (9.7) 0 0 1 (100) 0 0 0 0 Polymyxins Colistin 1 (1.6) 0 3 (100) 1 (100) 0 0 1 (100) 0
下载: 导出CSV
-
[1] Stratev D, Odeyemi OA. Antimicrobial resistance of Aeromonas hydrophila isolated from different food sources: A mini-review. J Infect Public Health, 2016; 9, 535−44. doi: 10.1016/j.jiph.2015.10.006 [2] Li ZQ, Wang YQ, Li XY, et al. The characteristics of antibiotic resistance and phenotypes in 29 outer-membrane protein mutant strains in Aeromonas hydrophila. Environ Microbiol, 2019; 21, 4614−28. doi: 10.1111/1462-2920.14761 [3] Janda JM, Abbott SL. The genus Aeromonas: Taxonomy, pathogenicity, and infection. Clin Microbiol Rev, 2010; 23, 35−73. doi: 10.1128/CMR.00039-09 [4] Hao RZ, Li P, Wang Y, et al. Diversity of pathogens responsible for acute diarrheal disease in China. Clin Infect Dis, 2013; 57, 1788−90. doi: 10.1093/cid/cit572 [5] Chen PL, Wu CJ, Chen CS, et al. A comparative study of clinical Aeromonas dhakensis and Aeromonas hydrophila isolates in southern Taiwan: A. dhakensis is more predominant and virulent. Clin Microbiol Infect, 2014; 20, O428−34. doi: 10.1111/1469-0691.12456 [6] Meng S, Du XL, Wang YL, et al. Comparative study of the genetic diversity, antimicrobial resistance, and pathogenicity of Aeromonas isolates from clinical patients and healthy individuals. Biomed Environ Sci, 2021; 34, 454−64. [7] Subbaram K, Gatasheh MK, Al Azzam KM, et al. Molecular identification of diarrheal Aeromonas using immuno magnetic polymerase chain reaction (IM-PCR) technique: a comparative study with conventional culture method. Afr Health Sci, 2019; 19, 2036−42. doi: 10.4314/ahs.v19i2.27 [8] Morinaga Y, Yanagihara K, Eugenin FLL, et al. Identification error of Aeromonas aquariorum: a causative agent of septicemia. Diagn Microbiol Infect Dis, 2013; 76, 106−9. doi: 10.1016/j.diagmicrobio.2013.01.019 [9] Miñana-Galbis D, Urbizu-Serrano A, Farfán M, et al. Phylogenetic analysis and identification of Aeromonas species based on sequencing of the cpn60 universal target. Int J Syst Evol Microbiol, 2009; 59, 1976−83. doi: 10.1099/ijs.0.005413-0 [10] Yano Y, Hamano K, Tsutsui I, et al. Occurrence, molecular characterization, and antimicrobial susceptibility of Aeromonas spp. in marine species of shrimps cultured at inland low salinity ponds. Food Microbiol, 2015; 47, 21−7. doi: 10.1016/j.fm.2014.11.003 [11] Senderovich Y, Ken-Dror S, Vainblat I, et al. A molecular study on the prevalence and virulence potential of Aeromonas spp. recovered from patients suffering from diarrhea in Israel. PLoS One, 2012; 7, e30070. doi: 10.1371/journal.pone.0030070 [12] Batra P, Mathur P, Misra MC. Aeromonas spp. : An emerging nosocomial pathogen. J Lab Physicians, 2016; 8, 1−4. doi: 10.4103/0974-2727.176234 [13] Tan MW, Rahme LG, Sternberg JA, et al. Pseudomonas aeruginosa killing of Caenorhabditis elegans used to identify P. aeruginosa virulence factors. Proc Natl Acad Sci U S A, 1999; 96, 2408−13. doi: 10.1073/pnas.96.5.2408 [14] Gravato-Nobre MJ, Hodgkin J. Caenorhabditis elegans as a model for innate immunity to pathogens. Cell Microbiol, 2005; 7, 741−51. doi: 10.1111/j.1462-5822.2005.00523.x [15] Wu CJ, Ko WC, Lee NY, et al. Aeromonas isolates from fish and patients in Tainan City, Taiwan: Genotypic and phenotypic characteristics. Appl Environ Microbiol, 2019; 85, e01360−19. [16] Wang XM, Wang Y, Zhou Y, et al. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg Microbes Infect, 2018; 7, 122. [17] Hossain S, De Silva BCJ, Wimalasena SHMP, et al. Distribution of antimicrobial resistance genes and Class 1 integron gene cassette arrays in motile Aeromonas spp. Isolated from Goldfish (Carassius auratus). Microb Drug Resist, 2018; 24, 1217−25. doi: 10.1089/mdr.2017.0388 [18] Figueira V, Vaz-Moreira I, Silva M, et al. Diversity and antibiotic resistance of Aeromonas spp. in drinking and waste water treatment plants. Water Res, 2011; 45, 5599−611. doi: 10.1016/j.watres.2011.08.021 [19] Zhou YY, Yu L, Nan Z, et al. Taxonomy, virulence genes and antimicrobial resistance of Aeromonas isolated from extra-intestinal and intestinal infections. BMC Infect Dis, 2019; 19, 158. doi: 10.1186/s12879-019-3766-0 [20] Chen PL, Wu CJ, Tsai PJ, et al. Virulence diversity among bacteremic Aeromonas isolates: ex vivo, animal, and clinical evidences. PLoS One, 2014; 9, e111213. doi: 10.1371/journal.pone.0111213 [21] Hossain S, Wickramanayake MVKS, Dahanayake PS, et al. Species identification, virulence markers and antimicrobial resistance profiles of Aeromonas sp. isolated from marketed hard-shelled mussel (Mytilus coruscus) in Korea. Lett Appl Microbiol, 2020; 70, 221−9. doi: 10.1111/lam.13266 [22] Meng S, Wang YL, Liu CG, et al. Genetic diversity, antimicrobial resistance, and virulence genes of Aeromonas isolates from clinical patients, tap water systems, and food. Biomed Environ Sci, 2020; 33, 385−95. [23] Martinez-Murcia AJ, Monera A, Saavedra MJ, et al. Multilocus phylogenetic analysis of the genus Aeromonas. Syst Appl Microbiol, 2011; 34, 189−99. doi: 10.1016/j.syapm.2010.11.014 [24] Hossain S, De Silva BCJ, Dahanayake PS, et al. Phylogenetic characteristics, virulence properties and antibiogram profile of motile Aeromonas spp. isolated from ornamental guppy (Poecilia reticulata). Arch Microbiol, 2020; 202, 501−9. doi: 10.1007/s00203-019-01762-5 [25] Pu W, Guo G, Yang N, et al. Three species of Aeromonas (A. dhakensis, A. hydrophila and A. jandaei) isolated from freshwater crocodiles (Crocodylus siamensis) with pneumonia and septicemia. Lett Appl Microbiol, 2019; 68, 212−8. doi: 10.1111/lam.13112 [26] Fernández-Bravo A, Figueras MJ. An update on the genus Aeromonas: Taxonomy, epidemiology, and pathogenicity. Microorganisms, 2020; 8, 129. doi: 10.3390/microorganisms8010129 [27] Castro-Escarpulli G, Figueras MJ, Aguilera-Arreola G, et al. Characterisation of Aeromonas spp. isolated from frozen fish intended for human consumption in Mexico. Int J Food Microbiol, 2003; 84, 41−9. doi: 10.1016/S0168-1605(02)00393-8 [28] Lau TTV, Tan JAMA, Puthucheary SD, et al. Genetic relatedness and novel sequence types of clinical Aeromonas dhakensis from Malaysia. Braz J Microbiol, 2020; 51, 909−18. doi: 10.1007/s42770-020-00239-8 [29] Abu-Elala N, Abdelsalam M, Marouf S, et al. Comparative analysis of virulence genes, antibiotic resistance and gyrB-based phylogeny of motile Aeromonas species isolates from Nile tilapia and domestic fowl. Lett Appl Microbiol, 2015; 61, 429−36. doi: 10.1111/lam.12484 [30] Rasmussen-Ivey CR, Figueras MJ, McGarey D, et al. Virulence factors of Aeromonas hydrophila: In the wake of reclassification. Front Microbiol, 2016; 7, 1337. [31] Rather MA, Willayat MM, Wani SA, et al. Enterotoxin gene profile and molecular epidemiology of Aeromonas species from fish and diverse water sources. J Appl Microbiol, 2019; 127, 921−31. doi: 10.1111/jam.14351 [32] Lamy B, Kodjo A, Laurent F. Prospective nationwide study of Aeromonas infections in France. J Clin Microbiol, 2009; 47, 1234−7. doi: 10.1128/JCM.00155-09 [33] Aravena-Román M, Inglis TJJ, Henderson B, et al. Antimicrobial susceptibilities of Aeromonas strains isolated from clinical and environmental sources to 26 antimicrobial agents. Antimicrob Agents Chemother, 2012; 56, 1110−2. doi: 10.1128/AAC.05387-11 [34] Ramadan H, Ibrahim N, Samir M, et al. Aeromonas hydrophila from marketed mullet (Mugil cephalus) in Egypt: PCR characterization of β-lactam resistance and virulence genes. J Appl Microbiol, 2018; 124, 1629−37. doi: 10.1111/jam.13734 [35] Dahanayake PS, Hossain S, Wickramanayake MVKS, et al. Prevalence of virulence and antimicrobial resistance genes in Aeromonas species isolated from marketed cockles (Tegillarca granosa) in Korea. Lett Appl Microbiol, 2020; 71, 94−101. doi: 10.1111/lam.13261 [36] Liu DJ, Song HW, Ke YB, et al. Co-existence of two novel phosphoethanolamine transferase gene variants in Aeromonas jandaei from retail fish. Int J Antimicrob Agents, 2020; 55, 105856. doi: 10.1016/j.ijantimicag.2019.11.013 [37] Picão RC, Poirel L, Demarta A, et al. Plasmid-mediated quinolone resistance in Aeromonas allosaccharophila recovered from a Swiss lake. J Antimicrob Chemother, 2008; 62, 948−50. doi: 10.1093/jac/dkn341 [38] Piotrowska M, Popowska M. The prevalence of antibiotic resistance genes among Aeromonas species in aquatic environments. Ann Microbiol, 2014; 64, 921−34. doi: 10.1007/s13213-014-0911-2 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 799
- HTML全文浏览量: 355
- PDF下载量: 119
- 被引次数: 0

 Quick Links
Quick Links