

 下载:
下载:

-
Exosomes are nano-sized (30–200 nm) phospholipid bilayer vesicles that are widely distributed in eukaryotic fluids, including plasma, saliva, and urine[1, 2]. They were long regarded as byproducts of membrane biosynthesis and abscission[3, 4], and then, exosome involvement in antigen presentation during immune cell activation in vivo was determined during the 1990s[5]. Thereafter, accumulated evidence has revealed that exosomes carry various biological molecules, including mRNAs, miRNAs, lncRNAs, DNA, lipids, and proteins, and are cell type-specific and have various biological functions[6, 7]. For example, the quantity and quality of exosomes derived from normal and malignant cells considerably differ, and they play important roles in various processes via intercellular communication[8, 9]. Exosomes not only influence the biological characteristics of adjacent cells but can also alter the fate of the recipient cells over a long distance[3, 10]. Chronic exposure to environmental chemicals results in alterations in exosomal content, secretion, and consequent pathological processes of environment-related diseases[11]. However, associations among chemical exposure, alteration of exosomal content, secretion, and disease progression remain largely unclear [11].
MicroRNAs (miRNAs), small short-chain (19-25 nucleotides) and non-coding RNAs, are important epigenetic regulators that promote the degradation of their target mRNA or inhibit their translation via complementary binding to 3’-untranslated region (3’-UTR) [12, 13]. The reported biological effects of miRNA include cell proliferation, differentiation, apoptosis, migration, and carcinogenesis[14, 15]. Abnormal miRNA expression is associated with the pathological processes of various diseases, and miRNA can function as typical oncogenes[16-18]. The most widely studied miRNA is miR-221, which is often upregulated in tumors and related to poor outcomes[19-21]. Nonetheless, the function of exosomal miR-221 in benzene-induced toxicity remains elusive.
Benzene is a common chemical that is also an established environmental carcinogen[22]. However, the mechanism of benzene-induced carcinogenesis remains unclear. We previously showed that exposure to benzene is associated with elevated miR-221 expression in peripheral blood lymphocytes from petrol station attendants[23]. However, the adverse health effects of elevated miR-221 content and whether it can be transferred to other cells and influence their biological characteristics via intercellular communication through exosomes is essentially unknown.
Here, we constructed hydroquinone (HQ, a representative metabolite of benzene with biological activity) -transformed malignant cell line, and investigated whether exosomal miR-221 derived from the transformed cells would influence apoptosis induced by HQ in normal recipient cells.
-
The Human Bronchial Epithelial Cells (16HBE) (American Type Culture Collection, Manassas, VA, USA) and HQ-transformed 16HBE (16HBE-t) cells were cultured in 1640 medium (Gibco, Laboratories, Gaithersburg, MD, USA) containing 10% fetal bovine serum (FBS), 10 U/mL penicillin, and 10 U/mL streptomycin (Hyclone Laboratories Inc., South Logan, UT, USA) in 5% CO2 at 37 ℃. The cells were digested with trypsin (Solarbio Co., Ltd., Beijing, China) when they reached 80% confluence and then seeded and passaged.
-
Soft agar assay was conducted to identify the biological characteristics of malignant 16HBE-t. Briefly, 1 mL per well of normal melting point agar (NMA; Solarbio) (0.6 w/v) in 1640 medium containing 10% FBS was added to 6-well plates and then solidified at 4 ℃ for 30 min. Thereafter, 3 × 103 cells/well in 1 mL of low melting agar (LMA, Solarbio) (0.3 w/v) in 1640 medium was added to the plates and incubated at 37 ℃ with 5% CO2 for 3 wk. Thereafter, we assessed 16HBE-t colony formation.
-
The viability of 16HBE and 16HBE-t cells was analyzed using CCK-8 assays (Dojindo Laboratories Co., Ltd., Kumamoto, Japan) as described by the manufacturer. Briefly, 1 × 103 16HBE or 16HBE-t cells/well in 100 μL complete medium were seeded into 96-well plates and incubated at 37 ℃ with 5% CO2 for 1–6 d. Thereafter, CCK-8 (10 μL/well) was added, and the cells were incubated for another 4 h at 37 ℃. Optical density (OD) was then determined using a spectrophotometer (BioTek Instruments, Winooski, VT, USA) at 450 nm. All groups of cells were assessed in triplicate.
-
A miR-221 inhibitor sequence was designed to inhibit miR-221 signals in 16HBE-t cells and synthesized at a commercial laboratory (RiboBio Co., Ltd., Guangzhou, China). The inhibitor was transfected into 16HBE-t cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA), and then exosomes were isolated.
-
A total of 1 × 106 cells were seeded into 10 cm2 cell culture plates and cultured with 100 mL serum- free conditioned culture medium for 3 d, washed three times with PBS, incubated with exosomes-free serum medium for 24 h, then the supernatant of which was collected. In order to remove dead cells and cell debris, the supernatant was centrifuged separately at 300 ×g for 30 min and at 16,500 ×g for 30 min in 4 ℃. Exosomes were then isolated via 120,000 ×g centrifugation for 1 h at 4 ℃ by using polypropylene centrifuge tubes (Beckman Coulter Instruments, Inc. Brea, CA, USA) on ultracentrifuge (HITACHI, Japan). Exosomal pellets were then resuspended in 100 μL PBS and stored in 4 ℃ for following research.
-
The morphological characteristics of exosomes were determined using a JEM-1200EX transmission electron microscope (TEM, Jeol Ltd., Tokyo, Japan). Briefly, 20 μL of the exosomal pellet supernatant was added to copper grids, dried at room temperature for 10 min, and then negatively stained with phosphotungstic acid (pH 7.0) for 5 min before analysis by transmission electron microscopy (TEM).
-
Exosome uptake was determined by confocal laser scanning microscopy. We agitated a mixture of 1 μL of the lipophilic stain PKH 67 (BestBio Co., Shanghai, China) that stably binds to the lipid region of the exosomal membrane, 25 μL of exosome suspension, and 250 μL of diluent C (BestBio) and then incubated it at 37 ℃ for 5 min. The reaction was terminated by adding 1 mL of PBS, and then, exosomes were incubated with recipient cells for 3 h at 37 ℃. The nuclei of recipient cells were stained with 4,6-diamidino-2-phenylindole (DAPI; BestBio Co.) at 37 ℃ for 15 min, and then the cells were fixed in 4% paraformaldehyde in PBS for 30 min. Exosome uptake was then assayed using an LSM700B laser scanning confocal microscope (Carl Zeiss Microscopy GmbH., Jena, Germany) at room temperature.
-
Total RNA was extracted from 16HBE and 16HBE-t using TRIzol (Invitrogen), and then cDNA was synthesized using Beastar qPCR RT Kits (DBI Bioscience, Ludwigshafen, Germany). Relative levels of miR-221 were also analyzed by qRT-PCR using the following internal control U6 and primer sequences.
miR-221 stem-loop (RT primer): 5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGAAACCCA-3′.
miR-221: Forward: 5′-ACACTCCAGCTGGGAGCTACATTGTC-3′.
Universal Reverse (Reverse primer): 5′-CTCAACTGGTGTCGTGGA-3′.
U6: Forward: 5′-CTCGCTTCGGCAGCACA-3′; Reverse: 5′- AACGCTTCACGAATTTGCGT-3′.
The qRT-PCR cycling conditions were 95 ℃ for 30 s, 40 cycles of 95 ℃ for 10 s, 56 ℃ for 30 s, and 72 ℃ for 30 s. Relative miR-221 expression levels were calculated using the 2-ΔΔCT method.
-
We previously found that the LD50 of HQ was about 38 μmol/L[24]. Therefore, we induced apoptosis in 16HBE cells using 25 μmol/L HQ. Apoptosis was assessed using Annexin V-fluorescein isothiocyanate (V-FITC)/propidium iodide (PI) Apoptosis Detection Kits (KeyGEN BioTECH Co., Ltd., Nanjing, China). Cells (5 × 105) suspended in 500 μL of binding buffer were mixed with 5 μL each of Annexin V-FITC and PI and incubated for 15 min at room temperature in darkness, and then apoptosis was analyzed using a Guava® easyCyte flow cytometer (Merck KGaA, Darmstadt, Germany).
-
Data were presented as mean ± standard deviation. All data were statistically analyzed using SPSS 19.0 (IBM Corp., Armonk, NY, USA) and GraphPad Prism 7.0.0 software (GraphPad Software, San Diego, CA, USA). Quantitative differences between groups were analyzed using Student t-tests, and apoptosis rates were compared among different groups using chi-square tests. The cutoff for statistical significance was P < 0.05.
-
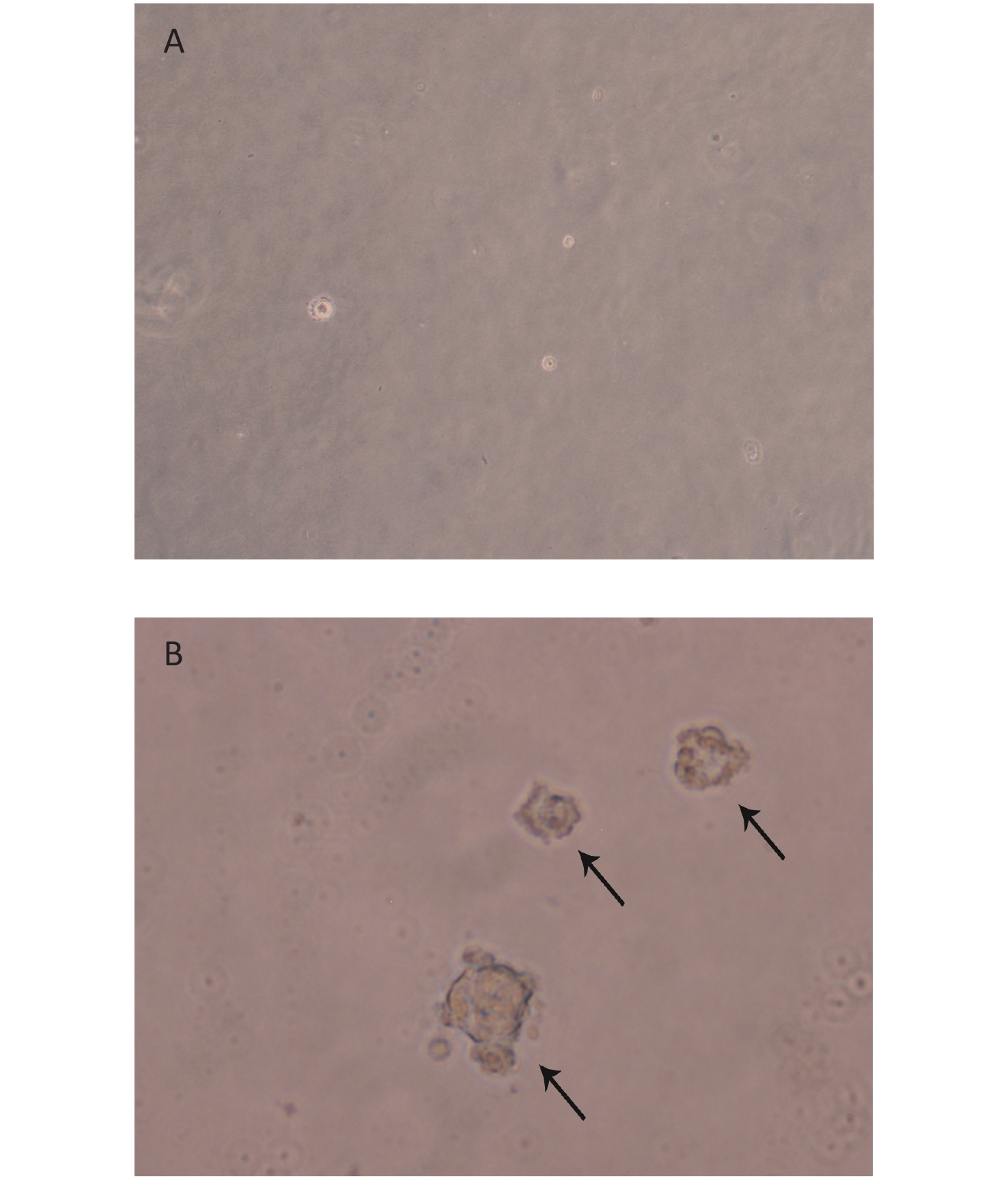
We created 16HBE-t cells as described[24]. The soft agar assay, which is a stringent test for malignant transformation of cells, and is semi-quantitative in nature to evaluate the response to treatment conditions[25], was used in malignant cell characterization. The results showed that 16HBE-t cells formed typical cell colonies in bilayers soft agar with about 6.85% efficiency. The colonies had the obvious morphological characteristic of anchorage-independent growth. While control group of 16HBE cells did not form colonies in bilayers soft agar (Figure 1).

Figure 1. Clone formation of 16HBE-t in bilayer soft agar. (A) Normal 16HBE cells could not form typical clone in bilayer soft agar (×100); (B) 16HBE-t cell clones (×100).
-
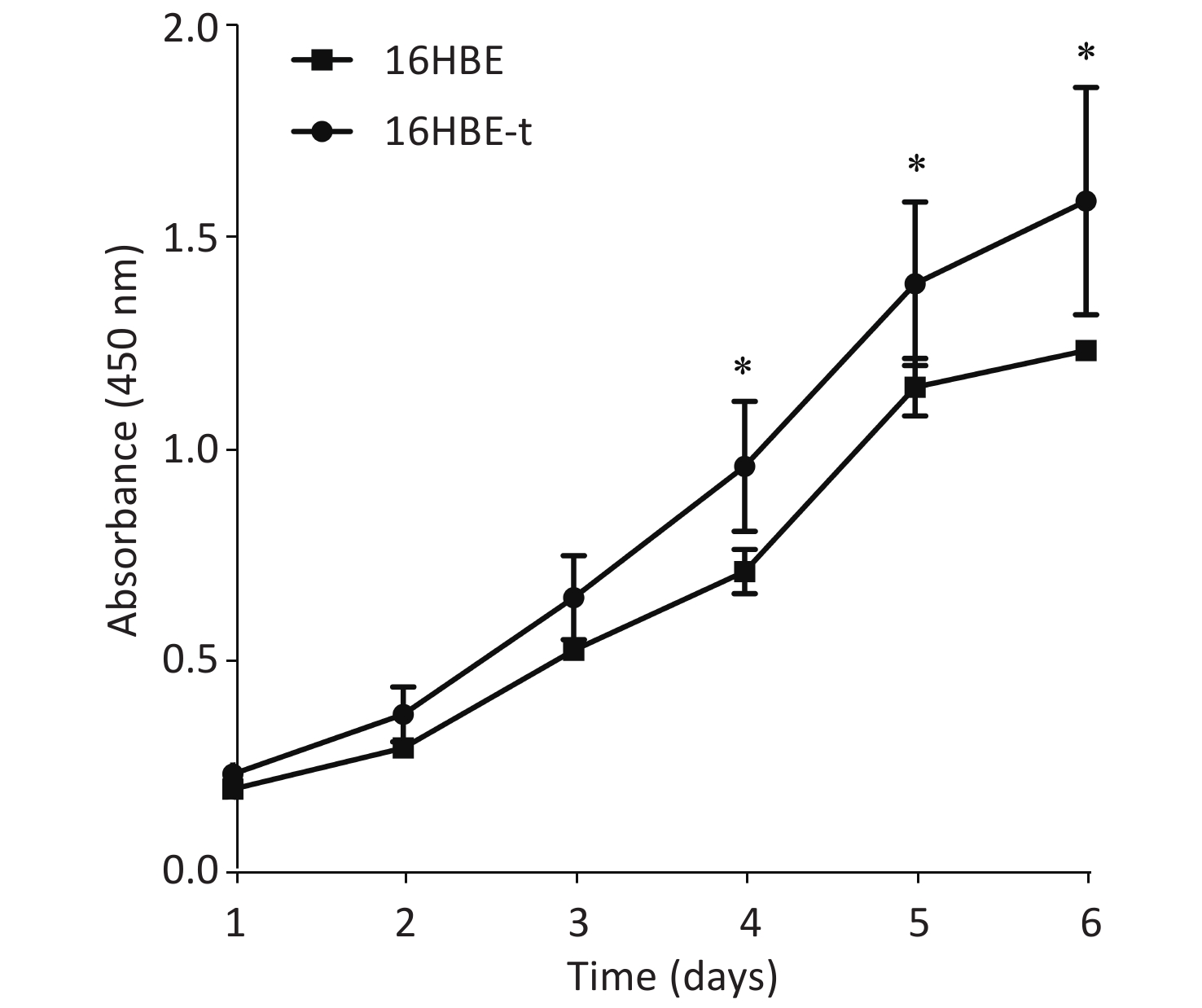
The proliferation of 16HBE and 16HBE-t cells was analyzed using CCK-8. Absorbance significantly differed (P < 0.05) between the two cell lines after 4–6 d in culture. The growth curve characteristics of 16HBE-t cells was typical of malignant cells (Figure 2).
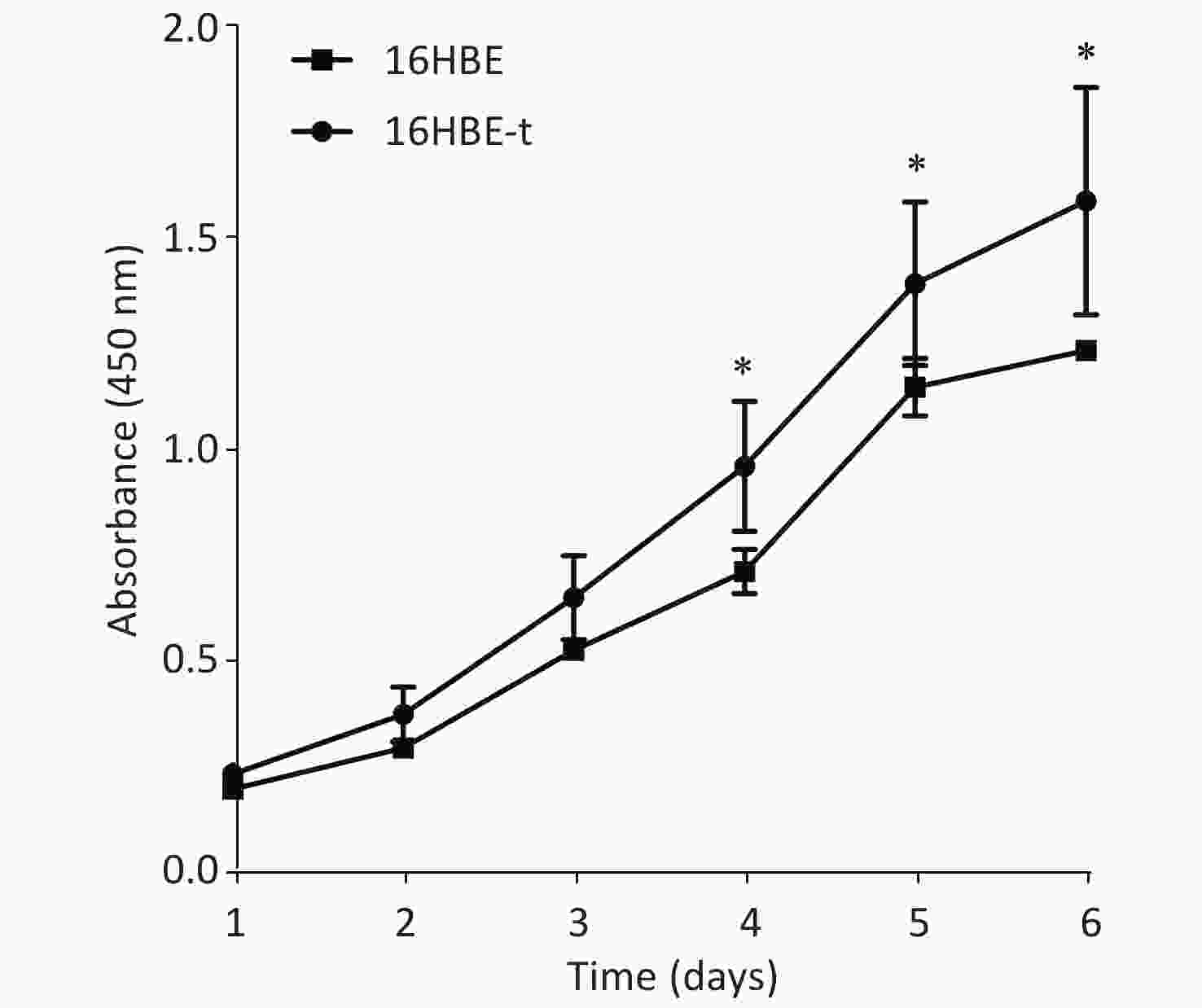
Figure 2. Growth curves of 16HBE and 16HBE-t cells (Three parallel samples are set for each group of cells) (*shows P < 0.05, i.e., there was significant difference in absorbance between 16HBE and 16HBE-t cells).
-
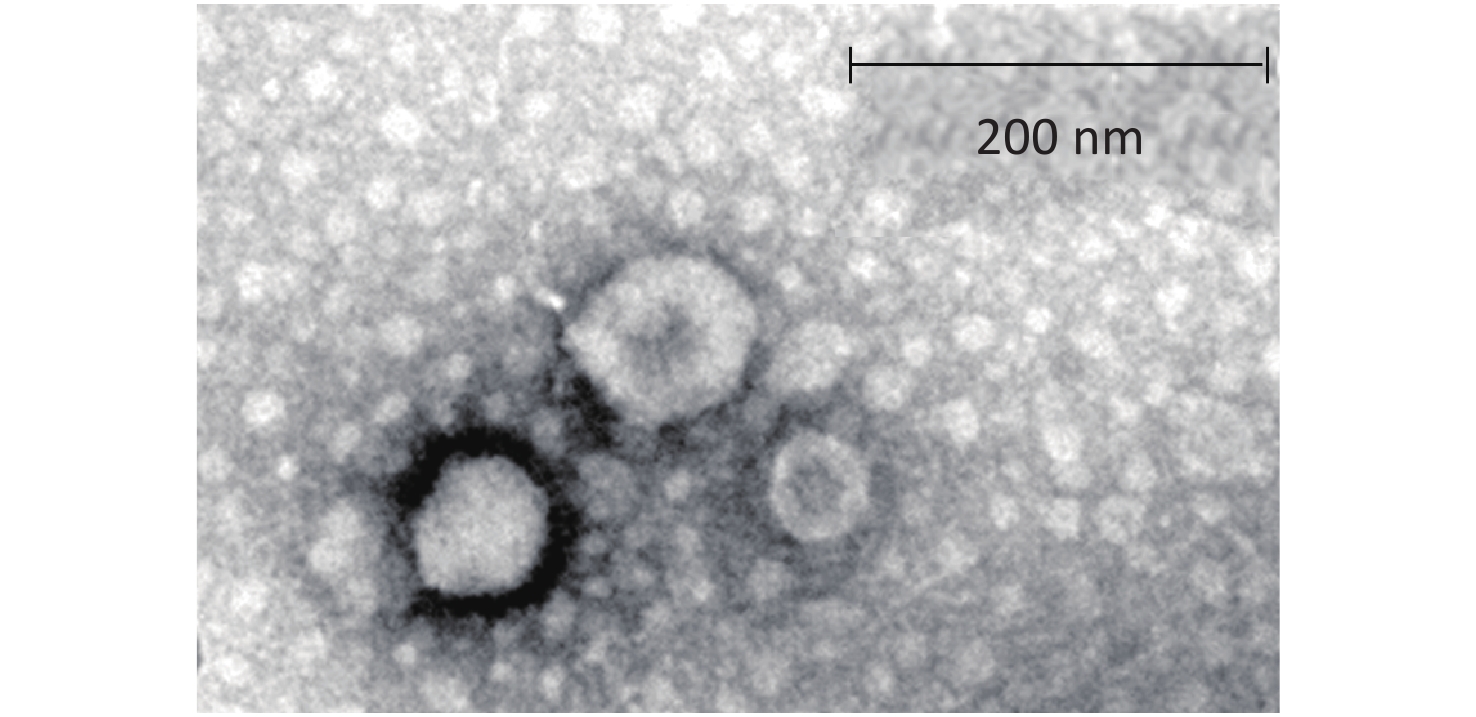
The TEM findings revealed exosomes with morphological characteristic of cup-shaped and typical double-layer membrane structure. As compared with the 200 nm scale bar, all the exosomes were within 60–120 nm in size, indicating that exosomes were isolated successfully (Figure 3).
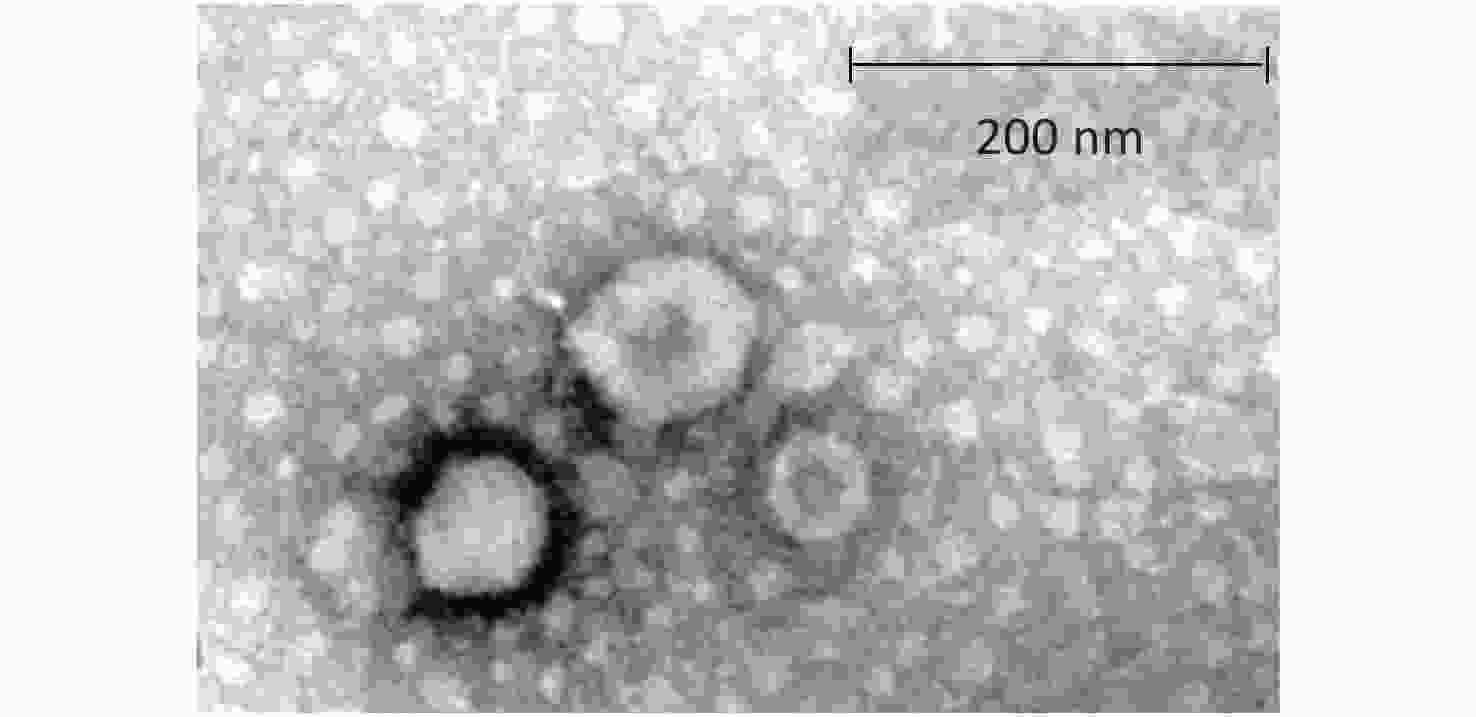
Figure 3. Morphological characteristic of exosomes under transmission electron microscope.
-
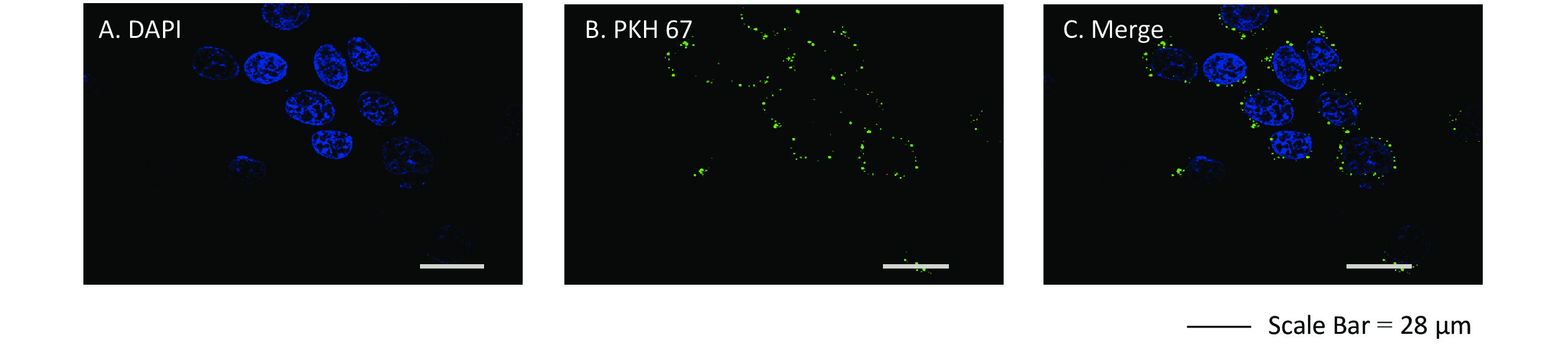
We assessed whether exosomes derived from 16HBE-t cells could be ingested by recipient 16HBE cells. The nuclei of 16HBE cells were stained blue using DAPI, and exosomes were stained in green with PKH 67. Laser scanning confocal microscope showed that recipient 16HBE cells ingested exosomes after 3 h of co-culture (Figure 4).
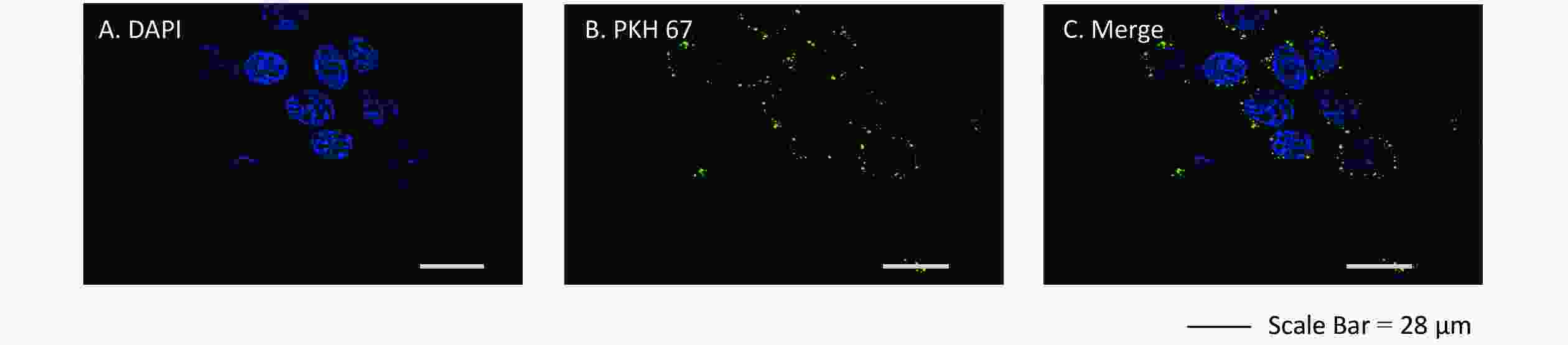
Figure 4. Confocal microscopy images of exosomes ingested by recipient 16HBE cells. (A) Nuclei stained with DAPI in 16HBE cells; (B) Exosomes stained with PKH 67; (C) Exosomes ingested by 16HBE cells.
-
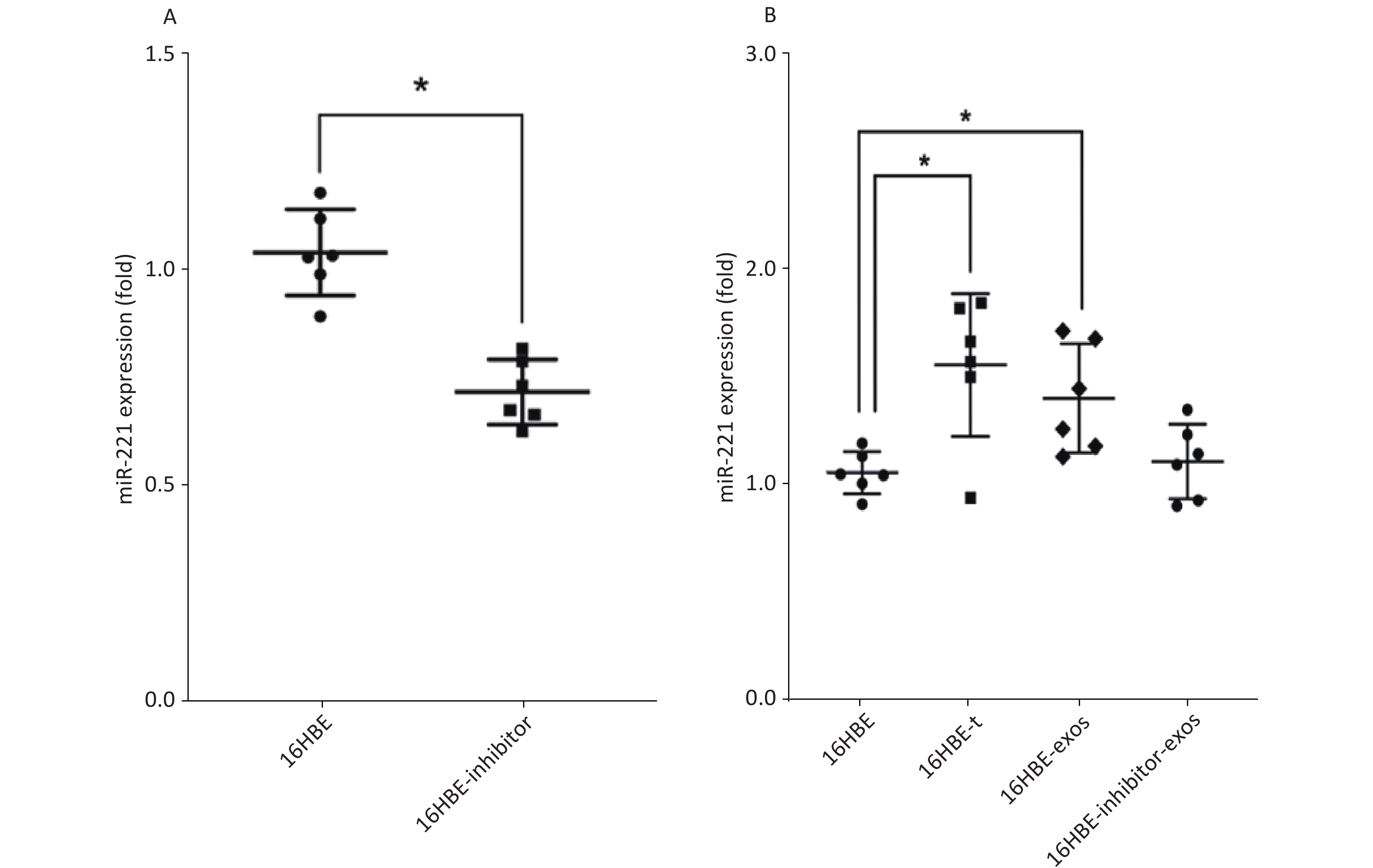
We detected the level of miR-221 using qRT-PCR. The data showed that miR-221 inhibitor could effectively inhibit the expression of miR-221 (P < 0.05; Figure 5A). Levels of miR-221 were significantly increased in 16HBE-t cells, compared with 16HBE cells (P < 0.05). After ingesting exosomes derived from 16HBE-t cells, miR-221 levels were significantly elevated in 16HBE-exos cells, compared with normal 16HBE cells. Levels of miR-221 in recipient cells did not significantly change after exosomal miR-221 in 16HBE-t cells was blocked using the inhibitor (Figure 5B).
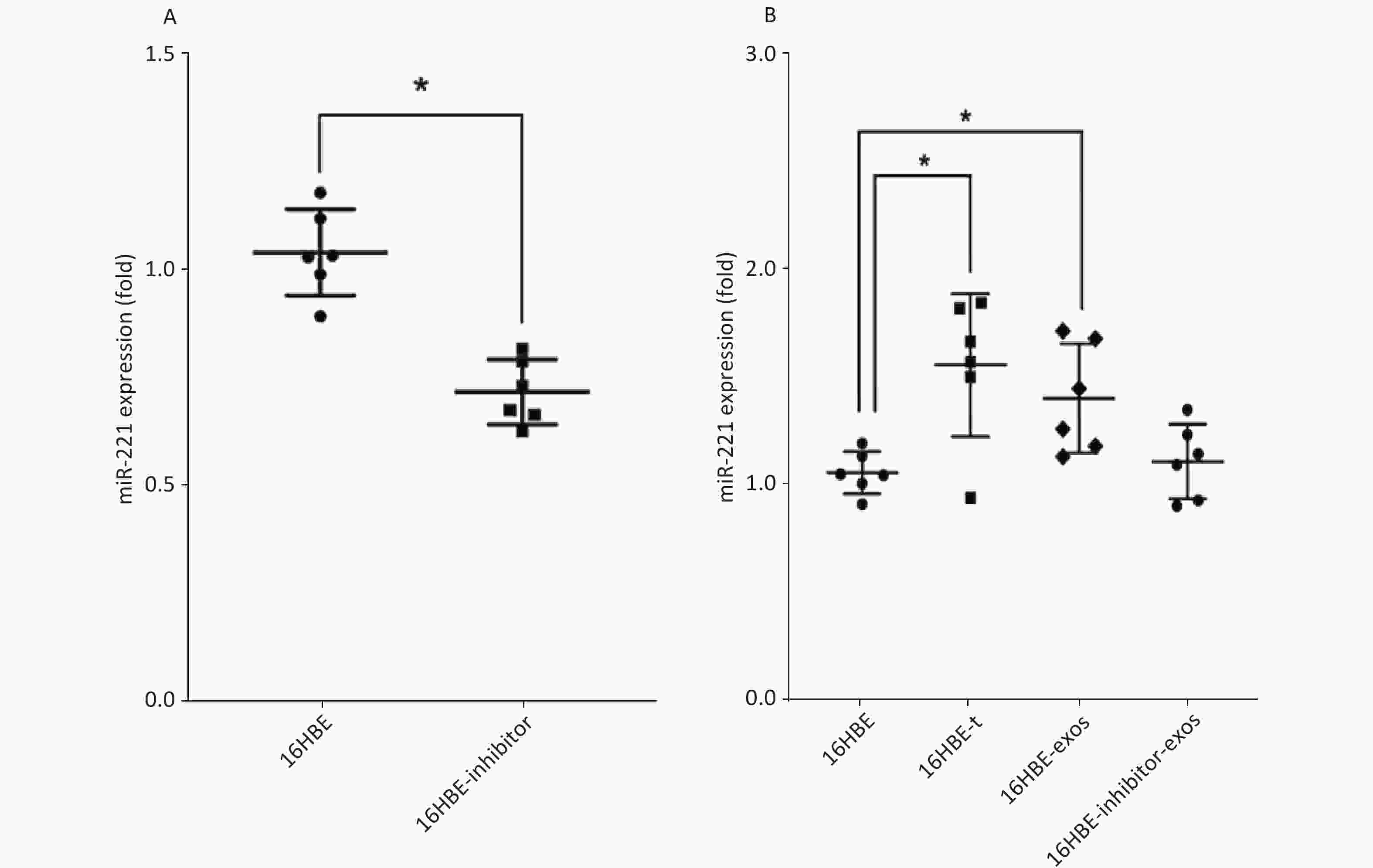
Figure 5. Result of qRT-PCR analysis of miR-221: (A) The efficiency of miR-221 inhibitor on miR-221 expression; (B) miR-221 expression level in different groups of cells (There are six parallel samples for each group of cells; *P < 0.05).
-
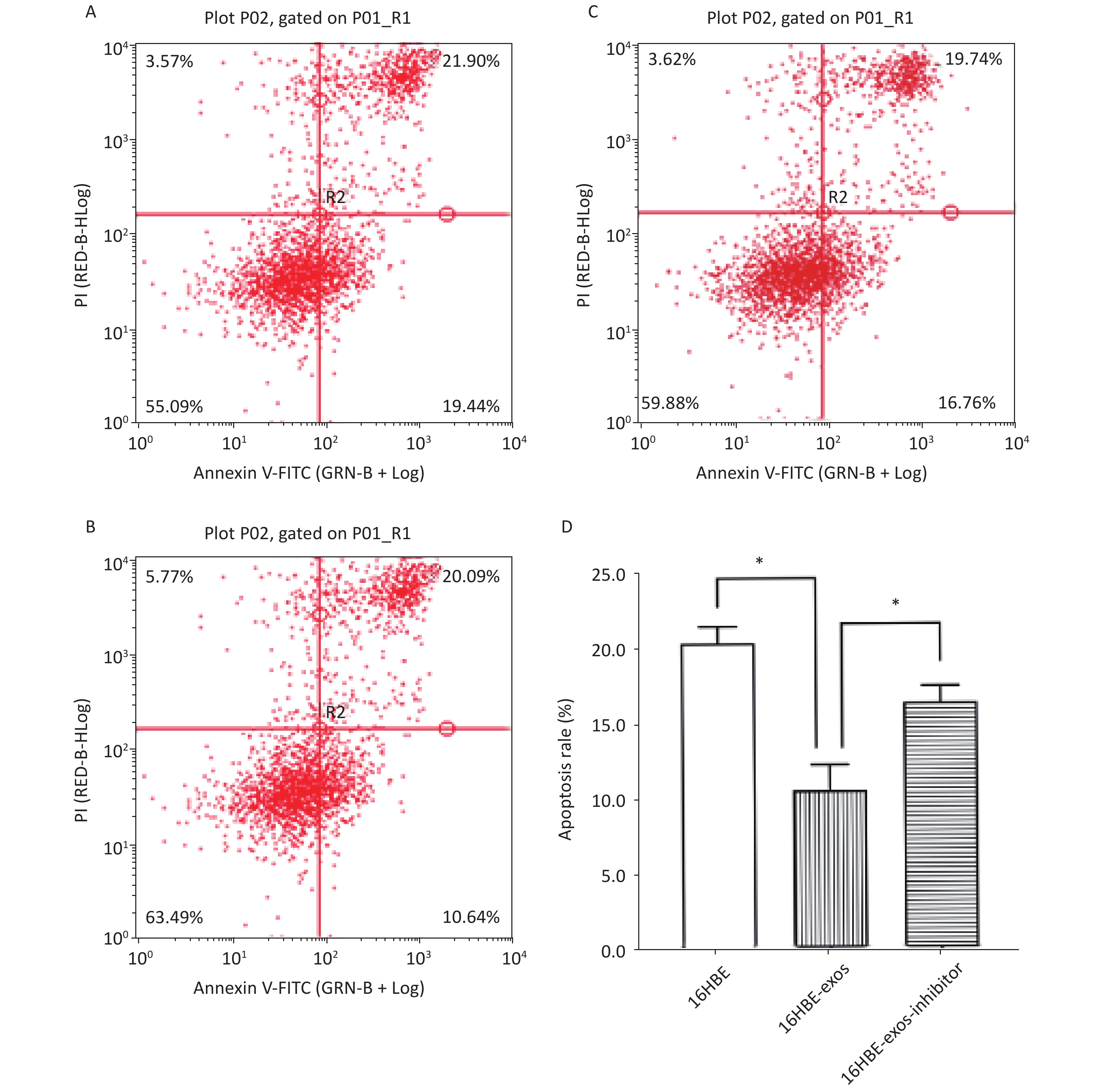
The effects of exosomal miR-221 on apoptosis induced by HQ were analyzed using flow cytometry. We found that HQ (25 μmol/L) induced obvious apoptosis in 16HBE cells. The apoptosis of 16HBE cells with ingested exosomes derived from 16HBE-t cells (16HBE-exos cells) decreased compared with normal 16HBE cells (P < 0.05). Notably, the apoptosis rate of 16HBE cells with ingested exosomes derived from 16HBE-t cells incubated with the miR-221 inhibitor did not significantly change (Figure 6).
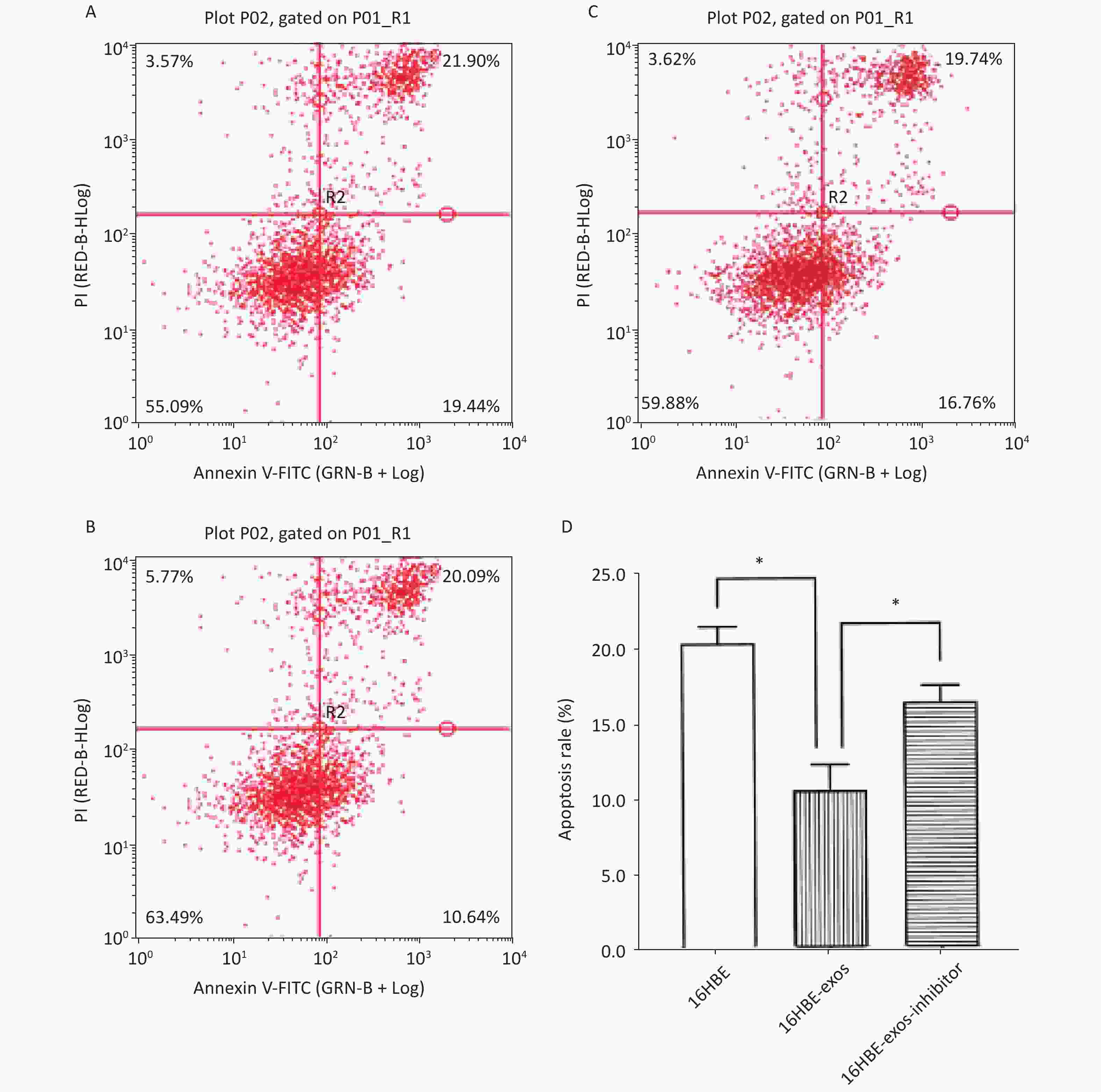
Figure 6. Inhibition effects of exosomal miR-221 on apoptosis induced by HQ in recipient cells. (A) Apoptosis induced by HQ in normal 16HBE cells; (B) Apoptosis induced by HQ in 16HBE-exos; (C) Apoptosis induced by HQ in 16HBE-exos-inhibitor; (D) Apoptosis rate of different groups of cells (* shows P < 0.05).
-
The mechanism of benzene-induced carcinogenesis remains unknown. The toxic effects of benzene on humans body is mainly mediated by its bioactive metabolites, among which HQ is the major representative and has frequently been applied in toxicological investigations of benzene[26-28]. Exosomes are associated with the occurrence and progression of various cancers[12, 13, 29]. This study demonstrated that HQ caused the malignant transformation of normal 16HBE cells. Significantly more miR-221 was expressed in 16HBE-t cells than in normal 16HBE cells, and miR-221 levels significantly enhanced in 16HBE cells after ingesting exosomes derived from 16HBE-t cells. Therefore, intercellular communication mediated by exosomes is involved in the toxic process of benzene. Furthermore, when 16HBE cells ingested the exosmes derived from 16HBE-t cells inhibited HQ-induced cell apoptosis. Apoptosis rates did not significantly change when exosomal miR-221 was inhibited, indicating that exosomal miR-221 reprogrammed the phenotype of the recipient cells, which might be a key event in the carcinogenesis of benzene. However, the tangible mechanism remains to be defined. For example, did exogenous miR-221 from exosomes itself increase the level of miR-221 in recipient cells? Or exogenous miR-221 from exosomes induced increase transcription of endogenous miR-221 in the recipient normal 16HBE cells? Or did both exist? Were there other pathways?
Apoptosis is programmed cell death that usually occurs via several mechanisms in response to internal and external stimuli such as exposure to environmental contaminants and radiation[30, 31]. The main role of apoptosis is to maintain the quantity of normal cells and control that of dysfunctional cells[32, 33]. Apoptosis involves obvious changes in morphological characteristics, including cell shrinkage, cytoplasm condensation, chromatin pyknosis, and nuclear fragmentation, it is considered to be strictly regulated by complex signaling networks [34]. Abnormal apoptosis often triggers pathological processes of various diseases, including tumors[35-37]. However, the tangible mechanisms of apoptosis remain unclear. The present study found that exosomal miR-221 derived from 16HBE-t cells inhibited apoptosis induced by HQ in normal recipient cells, indicating that it plays an important role in the progress of toxicity of benzene. However, the specific signaling pathways involved require further investigation.
Phosphatase and tensin homolog on chromosome 10 (PTEN), a typical tumor suppressor gene, can negatively regulate the PI3K/Akt signaling pathway and act as a molecular switch of the PTEN/PI3K/Akt axis that is involved in cell proliferation, apoptosis, migration, and invasion and plays an important role in carcinogenesis induced by exogenous chemicals[38-41]. Furthermore, PTEN is a direct target for the epigenetic regulation of miR-221[20, 42, 43], and the inactivation and dysfunction of PTEN in somatic cells are common in a variety of cancer patients, such as colon cancer and endometrial cancer, etcs.[31, 44]. The present study revealed that exosomes secreted by 16HBE-t cells inhibited HQ-induced apoptosis in recipient cells via miR-221 transfer. Therefore, we speculate that these biological phenomena mediated by exosomes are associated with the PTEN/PI3K/Akt pathway. Specifically, a novel exosomal-miR-221/PTEN/PI3K/Akt signaling pathway might be involved in the progress of the toxicity of benzene. We plan to follow up this speculation in our next study.
-
Taken together, the present investigation suggests that exosomal miR-221 secreted by 16HBE-t cells can be ingested by normal cells, thus results in inhibition of the apoptosis induced by HQ, which may be an important event in the toxic process of benzene and may be a consequence of human exposure to benzene at levels found in the environment, such as newly decorated houses, gas stations, shoe factories, paint factories and furniture factories, etc.
-
The authors have no conflicts of interest to report.
-
XIAN Hong Yi, CHEN Ying, ZHANG Jia Ying performed experimental study and paper writing; TANG Mei Lin, LIAN Zhen Wei, JIANG Ran, HU Zu Qing, LI Yang Feng performed experimental study, provided important suggestions and revised the paper; HU Da Lin is the corresponding author, who responsible for design, organizing, implementation and supervision of this project.
doi: 10.3967/bes2021.072
Exosomes Derived from Hydroquinone-transformed Human Bronchial Epithelial Cells Inhibited Recipient Cell Apoptosis by transferring miR-221
-
Abstract:
Objective Although benzene is a confirmed environmental carcinogen, the mechanism of its carcinogenicity remains largely unclear. The suggested oncogene, miR-221, is elevated and plays important roles in various tumors, but its role in benzene-induced carcinogenesis remains unknown. Methods In the present study, we constructed hydroquinone (HQ, a representative metabolite of benzene with biological activity)-transformed malignant cell line (16HBE-t) and analyzed the level of miR-221 in it with qRT-PCR. Exosomes from 16HBE-t cells incubated with or without an miR-221 inhibitor were isolated by ultracentrifugation, characterized by transmission electron microscopy and laser scanning confocal microscope, and then transfected into 16HBE cells. The effects of exosomal miR-221 on apoptosis induced by HQ in recipient cells were determined using flow cytometry. Results The amount of miR-221 in 16HBE-t was significantly increased compared with controls. When recipient cells ingested exosomes derived from 16HBE-t, miR-221 was increased, and apoptosis induced by HQ was inhibited. Blocking miR-221 in 16HBE-t using an inhibitor did not significantly alter miR-221 or apoptosis in recipient cells. Conclusion Exosomal miR-221 secreted by 16HBE-t inhibits apoptosis induced by HQ in normal recipient cells. -
Key words:
- Benzene /
- Toxic mechanism /
- Apoptosis /
- Exosomes /
- miR-221 /
- Intercellular communications
注释: -
Figure 1. Clone formation of 16HBE-t in bilayer soft agar. (A) Normal 16HBE cells could not form typical clone in bilayer soft agar (×100); (B) 16HBE-t cell clones (×100).
Figure 2. Growth curves of 16HBE and 16HBE-t cells (Three parallel samples are set for each group of cells) (*shows P < 0.05, i.e., there was significant difference in absorbance between 16HBE and 16HBE-t cells).
Figure 3. Morphological characteristic of exosomes under transmission electron microscope.
Figure 4. Confocal microscopy images of exosomes ingested by recipient 16HBE cells. (A) Nuclei stained with DAPI in 16HBE cells; (B) Exosomes stained with PKH 67; (C) Exosomes ingested by 16HBE cells.
Figure 5. Result of qRT-PCR analysis of miR-221: (A) The efficiency of miR-221 inhibitor on miR-221 expression; (B) miR-221 expression level in different groups of cells (There are six parallel samples for each group of cells; *P < 0.05).
Figure 6. Inhibition effects of exosomal miR-221 on apoptosis induced by HQ in recipient cells. (A) Apoptosis induced by HQ in normal 16HBE cells; (B) Apoptosis induced by HQ in 16HBE-exos; (C) Apoptosis induced by HQ in 16HBE-exos-inhibitor; (D) Apoptosis rate of different groups of cells (* shows P < 0.05).
-
[1] Tang YT, Huang YY, Zheng L, et al. Comparison of isolation methods of exosomes and exosomal RNA from cell culture medium and serum. Int J Mol Med, 2017; 40, 834−44. doi: 10.3892/ijmm.2017.3080 [2] Wu MX, Ouyang YS, Wang ZY, et al. Isolation of exosomes from whole blood by integrating acoustics and microfluidics. Proc Natl Acad Sci USA, 2017; 114, 10584−9. doi: 10.1073/pnas.1709210114 [3] Kalluri R. The biology and function of exosomes in cancer. J Clin Invest, 2016; 126, 1208−15. doi: 10.1172/JCI81135 [4] Hessvik NP, Llorente A. Current knowledge on exosome biogenesis and release. Cell Mol Life Sci, 2018; 75, 193−208. doi: 10.1007/s00018-017-2595-9 [5] Raposo G, Nijman HW, Stoorvogel W, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med, 1996; 183, 1161−72. doi: 10.1084/jem.183.3.1161 [6] Street JM, Koritzinsky EH, Glispie DM, et al. Urine exosomes: an emerging trove of biomarkers. Adv Clin Chem, 2017; 78, 103−22. [7] Samanta S, Rajasingh S, Drosos N, et al. Exosomes: new molecular targets of diseases. Acta Pharmacol Sin, 2018; 39, 501−13. doi: 10.1038/aps.2017.162 [8] Ngalame NNO, Luz AL, Makia N, et al. Arsenic alters exosome quantity and cargo to mediate stem cell recruitment into a cancer stem cell-like phenotype. Toxicol Sci, 2018; 165, 40−9. doi: 10.1093/toxsci/kfy176 [9] Poe AJ, Knowlton AA. Exosomes and cardiovascular cell-cell communication. Essays Biochem, 2018; 62, 193−204. doi: 10.1042/EBC20170081 [10] Huber HJ, Holvoet P. Exosomes: emerging roles in communication between blood cells and vascular tissues during atherosclerosis. Curr Opin Lipidol, 2015; 26, 412−9. doi: 10.1097/MOL.0000000000000214 [11] Harischandra DS, Ghaisas S, Rokad D, et al. Exosomes in toxicology: relevance to chemical exposure and pathogenesis of environmentally linked diseases. Toxicol Sci, 2017; 158, 3−13. doi: 10.1093/toxsci/kfx074 [12] Moutinho C, Esteller M. MicroRNAs and epigenetics. Adv Cancer Res, 2017; 135, 189−220. [13] Vaiman D. Genes, epigenetics and miRNA regulation in the placenta. Placenta, 2017; 52, 127−33. doi: 10.1016/j.placenta.2016.12.026 [14] Kim VN. MicroRNA biogenesis: coordinated cropping and dicing. Nat Rev Mol Cell Biol, 2005; 6, 376−85. [15] Xie LM, Zhang ZW, Tan ZQ, et al. MicroRNA-124 inhibits proliferation and induces apoptosis by directly repressing EZH2 in gastric cancer. Mol Cell Biochem, 2014; 392, 153−9. doi: 10.1007/s11010-014-2028-0 [16] Li CL, Deng L, Zhi QM, et al. MicroRNA-183 functions as an oncogene by regulating PDCD4 in gastric cancer. Anticancer Agents Med Chem, 2016; 16, 447−55. doi: 10.2174/1871520615666150914114237 [17] Zhang XY, Chen YF, Zhao P, et al. MicroRNA-19a functions as an oncogene by regulating PTEN/AKT/pAKT pathway in myeloma. Leuk Lymphoma, 2017; 58, 932−40. doi: 10.1080/10428194.2016.1213827 [18] Paliouras AR, Monteverde T, Garofalo M. Oncogene-induced regulation of microRNA expression: implications for cancer initiation, progression and therapy. Cancer Lett, 2018; 421, 152−160. doi: 10.1016/j.canlet.2018.02.029 [19] Garofalo M, Quintavalle C, Romano G, et al. miR221/222 in cancer: their role in tumor progression and response to therapy. Curr Mol Med, 2012; 12, 27−33. doi: 10.2174/156652412798376170 [20] Li BL, Lu Y, Yu LH, et al. miR-221/222 promote cancer stem-like cell properties and tumor growth of breast cancer via targeting PTEN and sustained Akt/NF-κB/COX-2 activation. Chem Biol Interact, 2017; 277, 33−42. doi: 10.1016/j.cbi.2017.08.014 [21] Liu SW, Wang YD, Li WG, et al. miR-221-5p acts as an oncogene and predicts worse survival in patients of renal cell cancer. Biomed Pharmacother, 2019; 119, 109406. doi: 10.1016/j.biopha.2019.109406 [22] Loomis D, Guyton KZ, Grosse Y, et al. Carcinogenicity of benzene. Lancet Oncol, 2017; 18, 1574−5. doi: 10.1016/S1470-2045(17)30832-X [23] Hu DL, Peng XW, Liu YG, et al. Overexpression of miR-221 in peripheral blood lymphocytes in petrol station attendants: a population based cross-sectional study in southern China. Chemosphere, 2016; 149, 8−13. doi: 10.1016/j.chemosphere.2016.01.083 [24] Jiang R, Huang HY, Lian ZW, et al. Exosomal miR-221 derived from hydroquinone-transformed malignant human bronchial epithelial cells is involved in cell viability of recipient cells. J Appl Toxicol, 2020; 40, 224−33. doi: 10.1002/jat.3898 [25] Borowicz S, Van Scoyk M, Avasarala S, et al. The soft agar colony formation assay. J Vis Exp, 2014; 92, e51998. [26] Enguita FJ, Leitao AL. Hydroquinone: environmental pollution, toxicity, and microbial answers. Biomed Res Int, 2013; 542168. [27] Li JQ, Jiang SF, Chen Y, et al. Benzene metabolite hydroquinone induces apoptosis of bone marrow mononuclear cells through inhibition of β-catenin signaling. Toxicol in Vitro, 2018; 46, 361−9. doi: 10.1016/j.tiv.2017.08.018 [28] Liang BX, Chen YC, Yuan WX, et al. Down-regulation of miRNA-451a and miRNA-486-5p involved in benzene-induced inhibition on erythroid cell differentiation in vitro and in vivo. Arch Toxicol, 2018; 92, 259−72. doi: 10.1007/s00204-017-2033-7 [29] Bullock MD, Silva AM, Kanlikilicer-Unaldi P, et al. Exosomal non-coding RNAs: diagnostic, prognostic and therapeutic applications in cancer. Non-Coding RNA, 2015; 1, 53−68. doi: 10.3390/ncrna1010053 [30] AnvariFar H, Amirkolaie AK, Miandare HK, et al. Apoptosis in fish: environmental factors and programmed cell death. Cell Tissue Res, 2017; 368, 425−39. doi: 10.1007/s00441-016-2548-x [31] Han WT, Wang S, Jiang LL, et al. Diallyl trisulfide (DATS) suppresses benzene-induced cytopenia by modulating haematopoietic cell apoptosis. Environ Pollut, 2017; 231, 301−10. doi: 10.1016/j.envpol.2017.07.069 [32] Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol, 2007; 35, 495−516. doi: 10.1080/01926230701320337 [33] Pistritto G, Trisciuoglio D, Ceci C, et al. Apoptosis as anticancer mechanism: function and dysfunction of its modulators and targeted therapeutic strategies. Aging (Albany NY), 2016; 8, 603−19. [34] Majtnerová P, T Roušar. An overview of apoptosis assays detecting DNA fragmentation. Mol Biol Rep, 2018; 45, 1469−78. doi: 10.1007/s11033-018-4258-9 [35] Ray T, Chakrabarti MK, Pal A. Hemagglutinin protease secreted by V. cholerae induced apoptosis in breast cancer cells by ROS mediated intrinsic pathway and regresses tumor growth in mice model. Apoptosis, 2016; 21, 143−54. doi: 10.1007/s10495-015-1194-1 [36] Hu XG, Jiang HJ, Jiang XJ. Downregulation of lncRNA ANRIL inhibits proliferation, induces apoptosis, and enhances radiosensitivity in nasopharyngeal carcinoma cells through regulating miR-125a. Cancer Biol Ther, 2017; 18, 331−8. doi: 10.1080/15384047.2017.1310348 [37] Sun Y, Li L, Wang L, et al. Body fluids-derived exosomes: paving the novel road to lung cancer diagnosis and therapy. Anticancer Agents Med Chem, 2017; 17, 1734−40. [38] Lee MS, Jeong MH, Lee HW, et al. PI3K/AKT activation induces PTEN ubiquitination and destabilization accelerating tumourigenesis. Nat Commun, 2015; 6, 7769. doi: 10.1038/ncomms8769 [39] Wang XX, Huang HQ, Young KH. The PTEN tumor suppressor gene and its role in lymphoma pathogenesis. Aging (Albany NY), 2015; 7, 1032−49. [40] Yuan L, Lv YR, Li HC, et al. Deubiquitylase OTUD3 regulates PTEN stability and suppresses tumorigenesis. Nat Cell Biol, 2015; 17, 1169−81. doi: 10.1038/ncb3218 [41] Andreas E, Hoelker M, Neuhoff C, et al. MicroRNA 17-92 cluster regulates proliferation and differentiation of bovine granulosa cells by targeting PTEN and BMPR2 genes. Cell Tissue Res, 2016; 366, 219−30. doi: 10.1007/s00441-016-2425-7 [42] Garofalo M, Di Leva G, Romano G, et al. miR-221&222 regulate TRAIL resistance and enhance tumorigenicity through PTEN and TIMP3 downregulation. Cancer Cell, 2009; 16, 498−509. doi: 10.1016/j.ccr.2009.10.014 [43] Sarkar S, Dubaybo H, Ali S, et al. Down-regulation of miR-221 inhibits proliferation of pancreatic cancer cells through up-regulation of PTEN, p27(kip1), p57(kip2), and PUMA. Am J Cancer Res, 2013; 3, 465−77. [44] de Araujo WM, Robbs BK, Bastos LG, et al. PTEN overexpression cooperates with lithium to reduce the malignancy and to increase cell death by apoptosis via PI3K/Akt suppression in colorectal cancer cells. J Cell Biochem, 2016; 117, 458−69. doi: 10.1002/jcb.25294 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1255
- HTML全文浏览量: 564
- PDF下载量: 83
- 被引次数: 0

 Quick Links
Quick Links