

 下载:
下载:



-
Aging is the most significant risk factor for numerous pathological conditions such as diabetes, cardiovascular diseases, neurodegenerative disorders, and cancer[1–6]. Senescence is a cellular response characterized by irreversible growth arrest and loss of proliferative capacity[7]. Although the biological basis of aging is far from understood, researchers have suggested that targeting the aging process itself can improve the outcomes of many age-related pathologies[8]. There is evidence to suggest that insulin-like growth factor (IGF) and insulin signaling pathways regulate aging in worms, insects, and mammals[9]. Studies in mice have revealed that lifespans can be prolonged by the heterozygous deletion of IGF-1 receptors[10], while another study has showed that the lifespans of monkeys can be extended by calorie restriction, which reduces the level of IGF-1[11]. Moreover, the robust extension of longevity in both sexes was detected in mice lacking growth hormone (GH) or GH receptors, in which the circulating level of IGF-1 was significantly suppressed[12–15]. However, the specific role of IGF-1 in the aging process has not been fully elucidated. POLD1, as a catalytic subunit of DNA polymerase δ, plays a critical role in DNA synthesis and DNA repair [16]. Moreover, POLD1 was demonstrated to be down-regulated in replicative senescence, which mediates the aging process[17]. In this study, IGF-1 was observed to inhibit the expression of POLD1 and to accelerate cell aging.
-
2BS cells were purchased from the National Infrastructure of Cell Line Resource (Beijing, China), and authenticated and tested for mycoplasma contamination at the time of purchase. 2BS cells were cultured in minimum essential medium (MEM, Gibco, New York, USA) supplemented with 10% fetal bovine serum (Gibco) and 1% penicillin–streptomycin (Gibco). The cells were cultured in a humidified incubator with 5% CO2 at 37 °C.
-
Human serum specimens from young donors (aged 18–30 years, n = 30) and old donors (aged 70 years and older, n = 30) with no history of disease at the time of blood collection were obtained from hospital staff, medical students, and healthy individuals visiting the health screening center of Xuanwu Hospital (Beijing, China). Subsequently, the serum specimens were sterilized by filtration and stored at −80 °C. The use of human specimens was approved by the institutional review board. Table 1 provides the information on the donors.
Table 1. Characteristics of donors in different ages
Characteristic 20–29 years (n = 35) 40–49 years (n = 33) 60–69 years (n = 30) 80–89 years (n = 24) Gender Male (n) 11 15 16 11 Female (n) 24 18 14 13 Age Mean ± SEM 24.76 ± 1.72 45.04 ± 1.41 64.54 ± 1.53 86.44 ± 3.34 Min-Max 21–29 43–47 62–68 81–89 -
2BS cells were seeded. Two days after incubation, the medium was replaced with medium containing 10% human serum, and the cells were cultured for 4 days to assess the effects of human serum from donors of different ages. Before replacing the culture medium, the cells were washed three times with phosphate-buffered saline. To evaluate the effects of IGF-1 on human aging, the 2BS cells cultured with old-age serum (O-Serum, 70 years of age and older) were treated with the inhibitor of the IGF-1 receptor (IGF-1R) or 50 μmol/L linsitinib (MCE, New Jersey, USA) for 24 h. To examine the effects of IGF-1 on aging, the 2BS cells were treated with 50 μmol/L linsitinib for 24 h, followed by 50 ng/mL IGF-1 (Cell Signaling, Danvers, USA) for 30 min. To detect the expression levels of IGF-1 and POLD1 during the kinetics of DNA repair, the 2BS cells were pre-treated with 50 μmol/L cisplatin (MCE) for 24 h to induce DNA damage, followed by incubation with old-age serum for 4 days or 50 μmol/L linsitinib for 24 h.
-
Six male SAMP8 mice were purchased from the First Affiliated Hospital of Tianjin University of Traditional Chinese Medicine (Tianjin, China). They were divided into two groups according to age as follows: young group (2 months of age, n = 6) and old group (8 months of age, n = 6). All animal experiments were approved by the bioethics committee of Xuanwu Hospital of Capital Medical University and compiled with the guidelines of the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
-
Western blotting analysis was performed as described previously[18]. In brief, the proteins were extracted, and equal concentrations of protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis. Next, the proteins were transferred onto polyvinylidene membranes. The primary antibodies against POLD1 and IGF-1 were purchased from Abcam (Cambridge, UK), and the primary antibody against β-actin was purchased from Zhongshan Boil Tech Co. (Beijing, China). After washing, the membranes were incubated with horseradish peroxidase-conjugated anti-rabbit or anti-mouse IgG secondary antibodies. The Chemi-Doc XRS+ imaging system was used for the chemiluminescent detection of target proteins. Table 2 provides the information on the antibodies.
Table 2. Antibodies used for Western blotting
Protein Manufacturer Cat no. Primary antibodies IGF-1 Abclonal Technology
Co. /ChinaA0830 POLD1 Abcam/UK Ab186407 β-actin Zhongshan Boil Tech
Co. /ChinaTA-09 Secondary antibodies HRP-conjugated
anti-mouse IgGZhongshan Boil Tech
Co. /ChinaZB2305 HRP-conjugated
anti-rabbit IgGZhongshan Boil Tech
Co. /ChinaZB5301 -
Total RNA was extracted using TRIzol reagent and reverse-transcribed according to the manufacturer’s instructions[18]. Target genes were amplified using a SYBR green fluorescence kit (Thermo Scientific, New York, USA) and a Roche 480 cycler (Roche, Basel, Switzerland). Quantification was performed by the 2−ΔΔCt method[18]. Table 3 provides the information on the primers for the amplification of POLD1, IGF-1, and β-actin genes.
Table 3. Sequences used for quantitative real-time polymerase chain reaction
Gene Sequences β-actin Forward ACAGAGCCTCGCCTTTGC Reverse CCACCATCACGCCCTGG IGF-1 Forward TGCTTCCGGAGCTGTGATCT Reverse TCTGGGTCTTGGGCATGTC POLD1 Forward GCTCCGCTCCTACACGCTCAA Reverse GGTCTGGTCGTTCCCATTCTGC -
SA-β-Gal staining was performed using a senescence β-galactosidase staining kit (Beyotime, Beijing, China) according to the manufacturer’s instructions. Positive staining was evaluated after 12–16 h of incubation at 37 °C in a CO2-free atmosphere. The positively-stained cells from three different fields were counted, and the results were presented as the percentage of positive cells.
-
Blood specimens were collected from healthy individuals of different ages. The serum concentrations of IGF-1 and POLD1 were measured using the human IGF-1 ELISA (Mlbio, Shanghai, China) and human POLD1 ELISA (Mlbio), respectively.
-
The proliferative ability of 2BS cells was detected using the CCK-8 assay (Beyotime). 2BS cells were seeded in 96-well plates at a density of 3 × 103 cells/well in 100 μL of medium per well. After adhesion, the cells underwent different treatments. The medium was replaced with 100 μL of fresh medium containing 10 μL of CCK-8, and the plates were incubated for 1 h. The absorbance was measured at a wavelength of 450 nm using a microplate reader.
-
The DNA synthesis rate of 2BS cells was assessed using the EdU kit (Beyotime). The cells were seeded in 96-well plates. After adhesion, the cells underwent different treatments. Next, the cells were incubated with 10 mmol/L EdU for 2 h, followed by staining according to the manufacturer’s instructions[18]. The absorbance was measured at a wavelength of 370 nm using an enzyme immunoassay analyzer.
-
The pLenti-CMV-POLD1 virus, which overexpressed POLD1, and shRNA-POLD1 virus, which silenced POLD1, were transfected into 2BS cells. The pLenti-CMV-POLD1 and shRNA-POLD1 virus were purchased from GeneChem (Shanghai, China). The supernatant was removed after 16 h of incubation and replaced with fresh medium.
-
The comet assay (Trevigen, Gaithersburg, MD, USA) was performed to evaluate DNA damage as previously described[18]. In brief, the cells were treated with 50 μmol/L cisplatin for 24 h to induce DNA damage, followed by incubation with old-age serum or normal serum for 4 days to repair DNA. Next, 50 μmol/L linsitinib was added for 24 h to the cells cultured with old-age serum. The cells were digested into a single-cell suspension, and the cell suspension was combined with the low melting point agarose gel and loaded onto the slides. The slides were immersed in lysis solution for 1 h, followed by unwinding for 1 h at 4 °C. Electrophoresis was performed at 21 volts for 30 min in alkaline electrophoresis solution. After electrophoresis, the slides were washed with deionized water and then stained with SYBR gold reagent (1:10,000 dilution) for 30 min. The state of the cells was observed with a fluorescence microscope (OLYMPUS, Tokyo Japan).
-
Data were analyzed using GraphPad Prism 8.0 software (San Diego, USA). The Student’s t-test analyzed the difference between the two groups, and one-way analysis of variance analyzed the difference among three or more groups. Pearson’s chi squared test calculated the relationship between IGF-1 and POLD1. P < 0.05 was considered to indicate a statistically significant result.
-
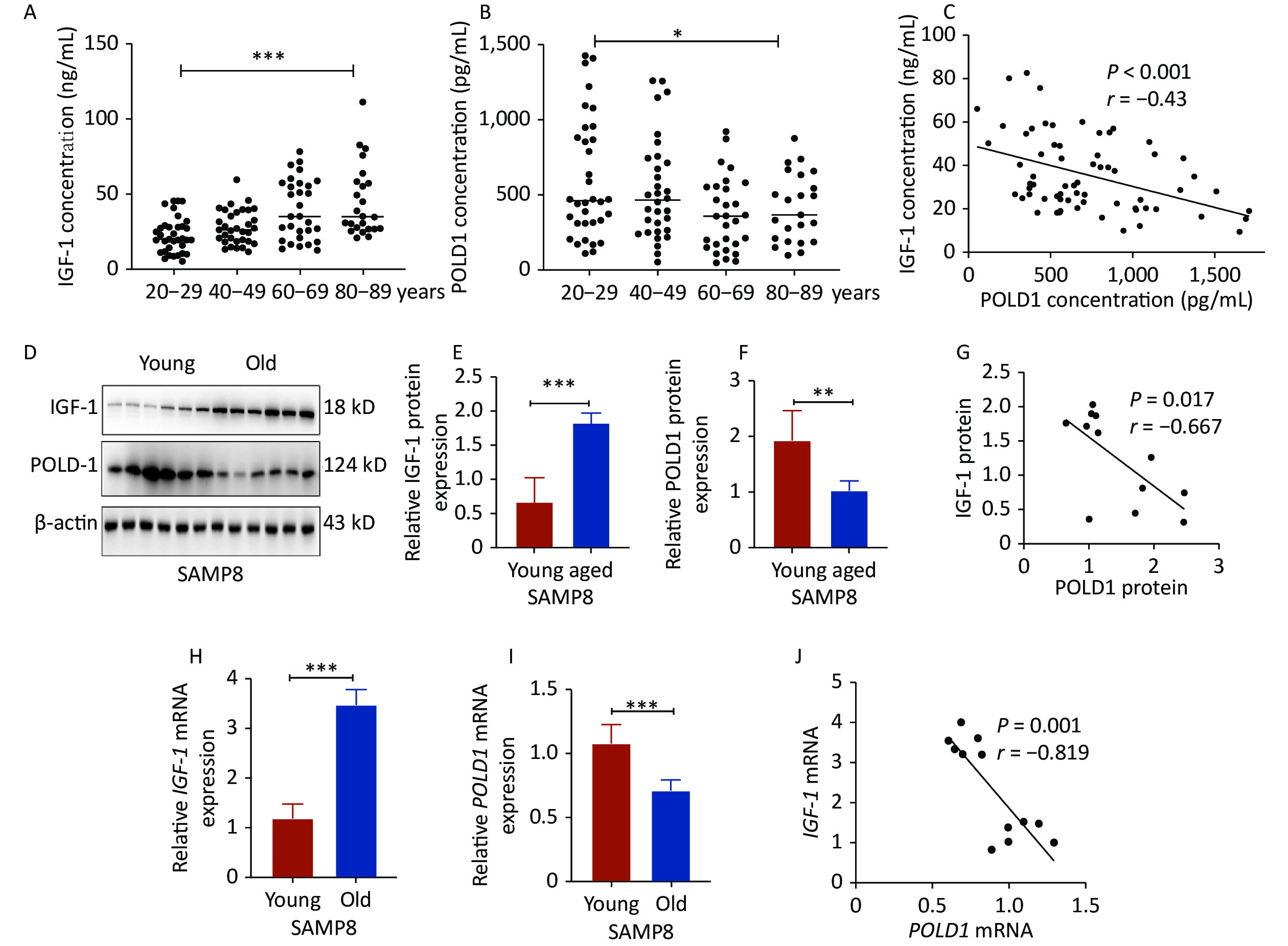
The levels of IGF-1 and POLD1 were analyzed in the serum of donors of different ages (20–29, 40–49, 60–69, 80–89 years old). The data showed a significant increase in the IGF-1 level (P < 0.001) and a significant decrease in the POLD1 level (P < 0.05) with age. A negative correlation was found between IGF-1 and POLD1 concentrations (P < 0.001, r = −0.43, Figure 1A–C, Supplementary Figure S1, available in www.besjournal.com). The protein and mRNA levels of IGF-1 and POLD1 were measured in hippocampal tissues from young (2 months old) and old (8 months old) SAMP8 mice. The results showed a significant increase in the IGF-1 level (protein level, P < 0.001; mRNA level, P < 0.001) and a significant decrease in the POLD1 level (protein level, P < 0.01; mRNA level, P < 0.05) with aging. There was a negative correlation between IGF-1 and POLD1 levels (protein levels, P = 0.017, r = −0.667; mRNA levels, P = 0.001, r = −0.819, Figure 1D–J).
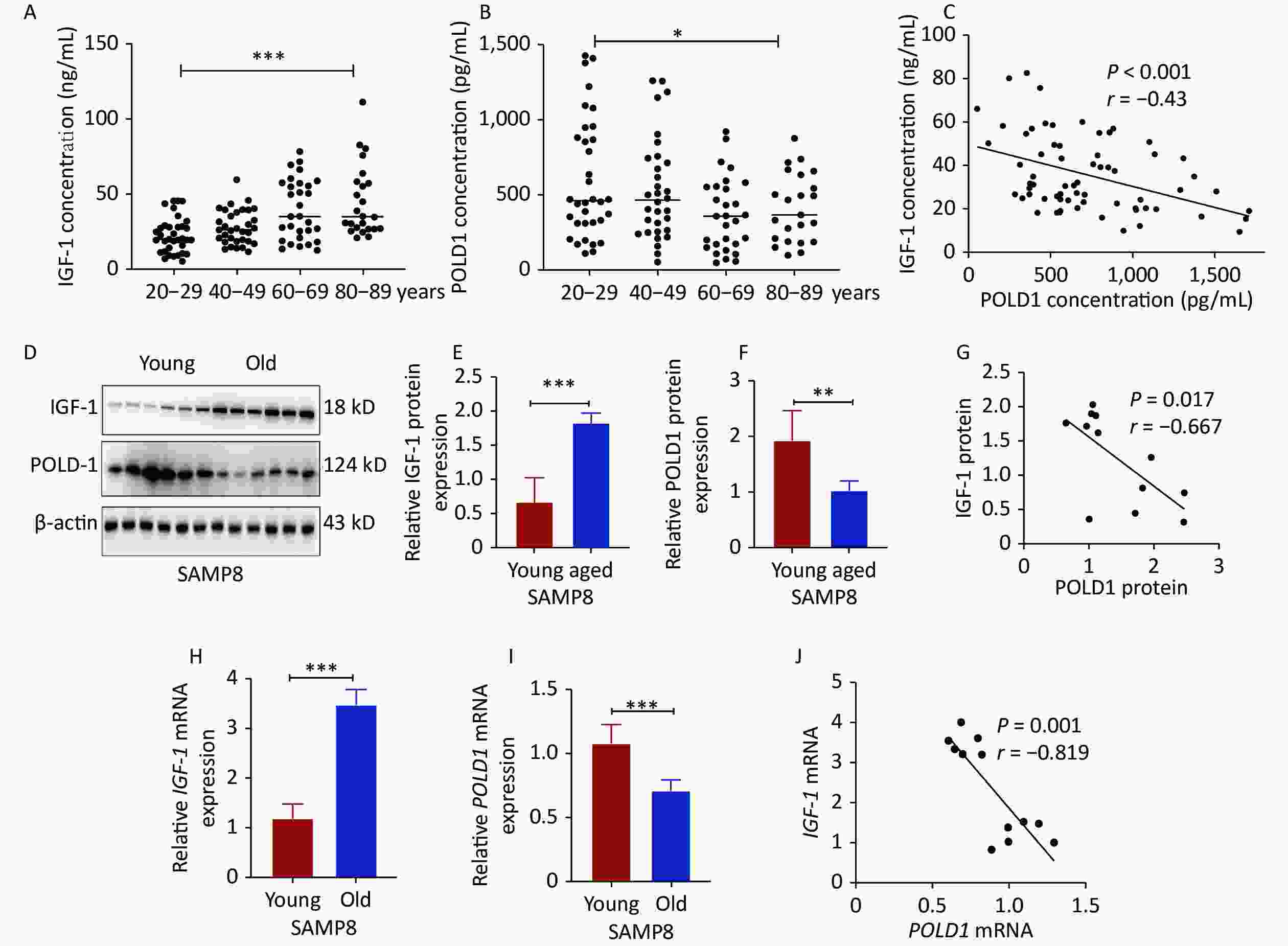
Figure 1. IGF-1 and POLD1 levels in the serum from healthy individuals of different ages and the hippocampal tissues from young and old SAMP8 mice. (A) IGF-1 concentrations in the human serum of different ages. (B) The concentrations of POLD1 in the human serum of different ages. (C) The relationship between IGF-1 and POLD1 concentrations in the human serum of different ages. (D–F) The protein expression levels of IGF-1 and POLD1 in the hippocampus of young and old SAMP8 mice. (G) The relationship between IGF-1 and POLD1 protein levels in the hippocampus of SAMP8 mice. (H–I) IGF-1 and POLD1 mRNA expression levels in the hippocampus of young and old SAMP8 mice. (J) The relationship between IGF-1 and POLD1 mRNA levels in the hippocampus of SAMP8 mice. (20–29, n = 35; 40–49, n = 33; 60–69, n = 30; 80–89, n = 24; young mice, 2 months, n = 6; old mice, 8 months, n = 6; *P < 0.05, ***P < 0.001).

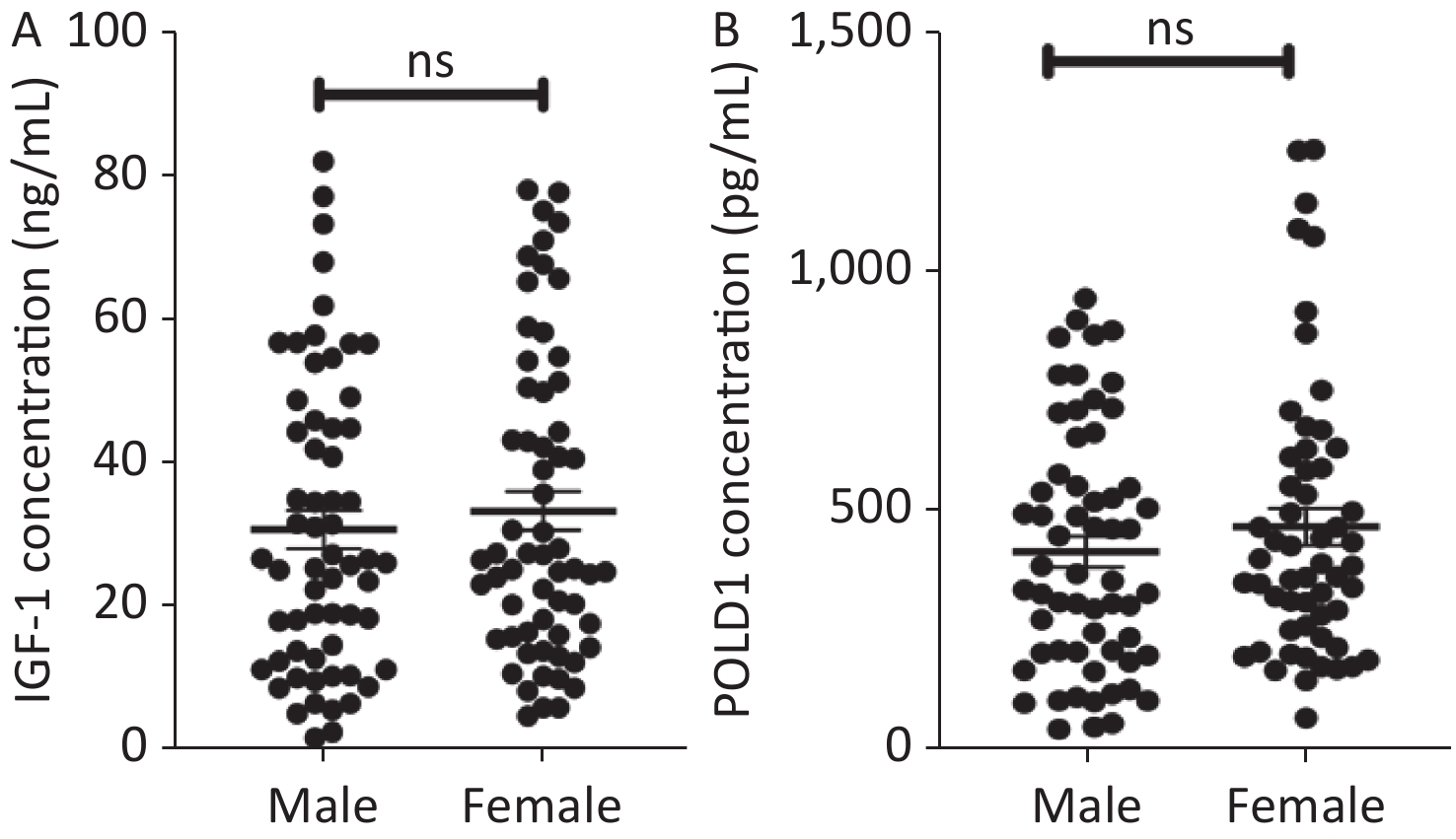
Figure S1. The expression of IGF-1 and POLD1 in human serum from male and female people. (A) IGF-1 concentrations in the human serum from male and female people. (B) The concentrations of POLD1 in the human serum from male and female people. (ns P > 0.05).
-
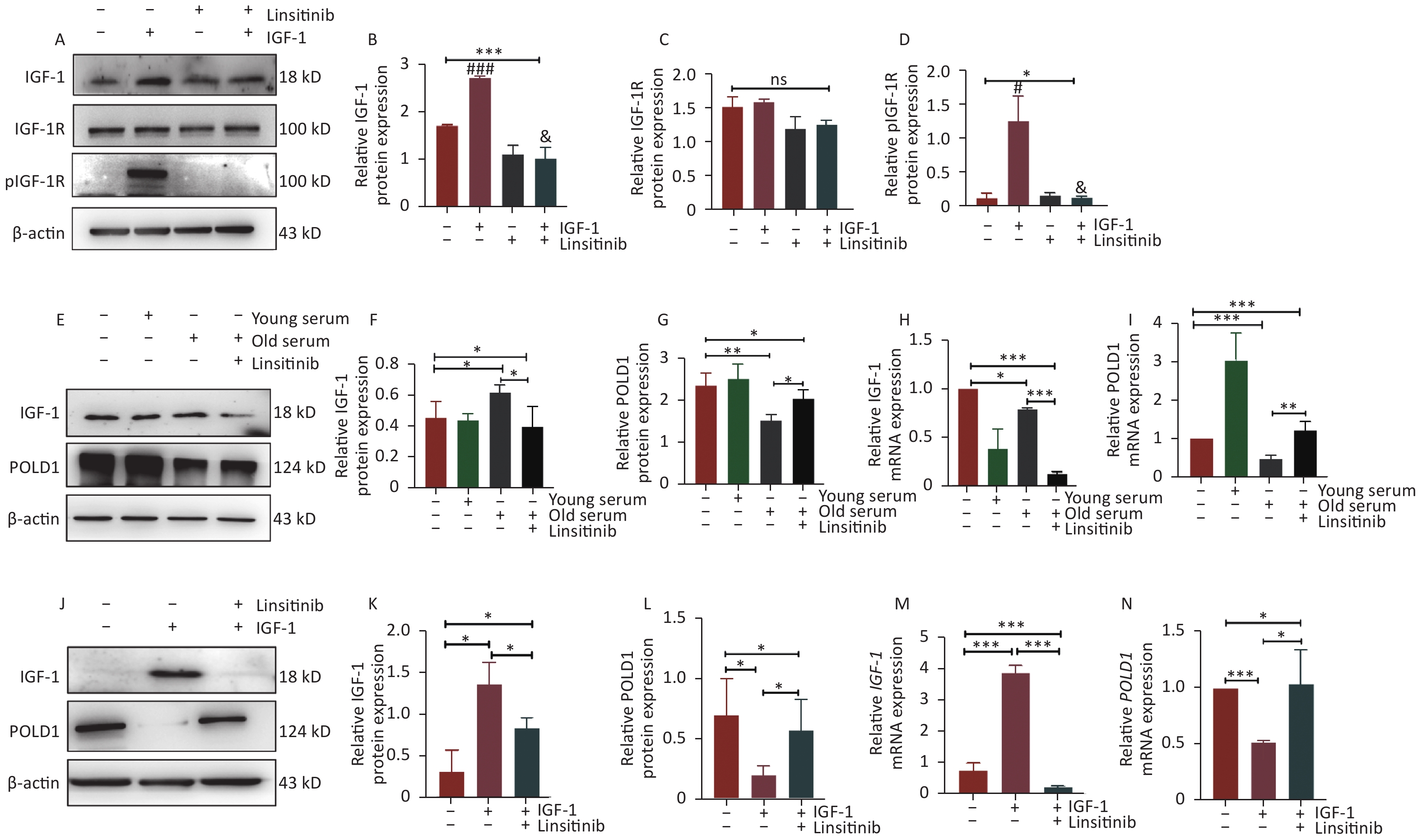
Linsitinib, as an inhibitor of IGF-1R, could inhibit the phosphorylation of IGF-1R to weaken the effect of IGF-1 (Figure 2A–D). To investigate the effect of IGF-1 on POLD1 expression, the POLD1 expression level was examined in the cells incubated with 10% human serum from young (18–30 years old) and old (≥ 70 years old) healthy donors, after considering the different concentrations of IGF-1 in the serum from the two groups of donors. Linsitinib was added to the cells cultured with the old-age human serum to examine the expression of IGF-1 and POLD1. The results showed that the expression level of POLD1 was higher in the cells cultured with young-age human serum than the cells cultured with old-age human serum. Moreover, the expression of POLD1 was up-regulated after linsitinib was added to the cells cultured with old-age human serum (Figure 2E–I). These results indicate that serum IGF-1 negatively regulates the expression of POLD1 by binding to IGF-1R.
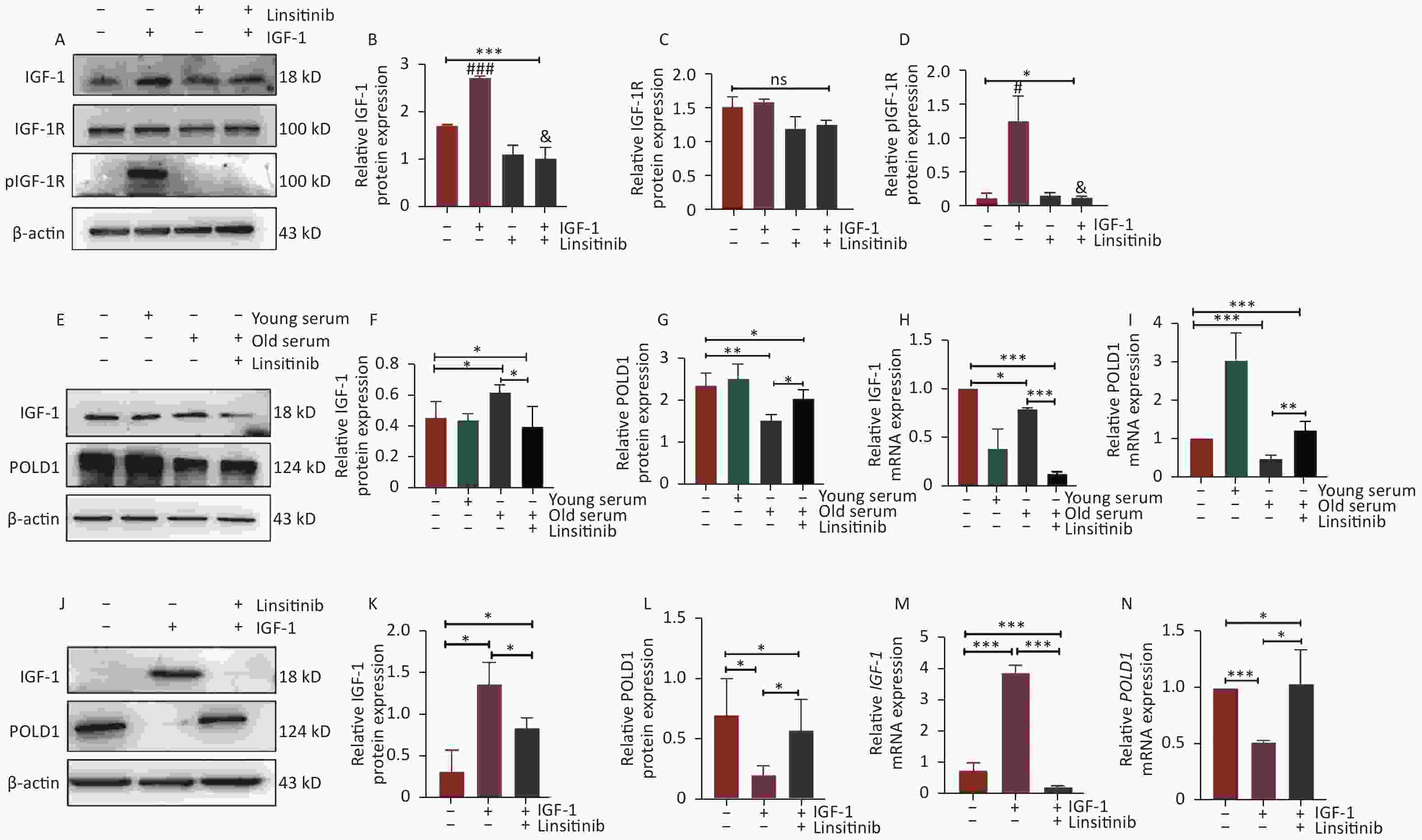
Figure 2. Expression levels of POLD1 and IGF-1 in 2BS cells incubated with young-age donor serum, old-age donor serum, old-age donor serum + linsitinib, and IGF-1 and IGF-1 + linsitinib, respectively. (A–D) The protein expression levels of IGF-1, IGF-1R, and pIGF-1R in 2BS cells cultured with IGF-1 or linsitinib and IGF-1 + linsitinib (#IGF-1 vs. NC, P < 0.05; ###IGF-1 vs. NC, P < 0.001; &IGF-1 + linsitinib vs. IGF-1, P < 0.05). (E–I) The protein and mRNA expression levels of IGF-1 and POLD1 in 2BS cells cultured with young-age donor serum, old-age donor serum, and old-age donor serum + linsitinib, respectively. (J–N) The protein and mRNA expression levels of IGF-1 and POLD1 in 2BS cells incubated with IGF-1 or IGF-1 + linsitinib. (Date are shown as the mean ± SEM, with three independent experiments in each group; ns P > 0.05, *P < 0.05, ***P < 0.001).
To further verify the effect of IGF-1 on POLD1 expression, the 2BS cells were treated with IGF-1 and linsitinib. The results showed that the protein and mRNA expression levels of IGF-1 were up-regulated significantly, whereas POLD1 levels were down-regulated in the cells cultured with IGF-1. By contrast, IGF-1 levels were down-regulated, whereas POLD1 levels were up-regulated in the cells cultured with linsitinib (Figure 2J–N). These results demonstrate that IGF-1 inhibits POLD1 expression by binding to IGF-1R.
-
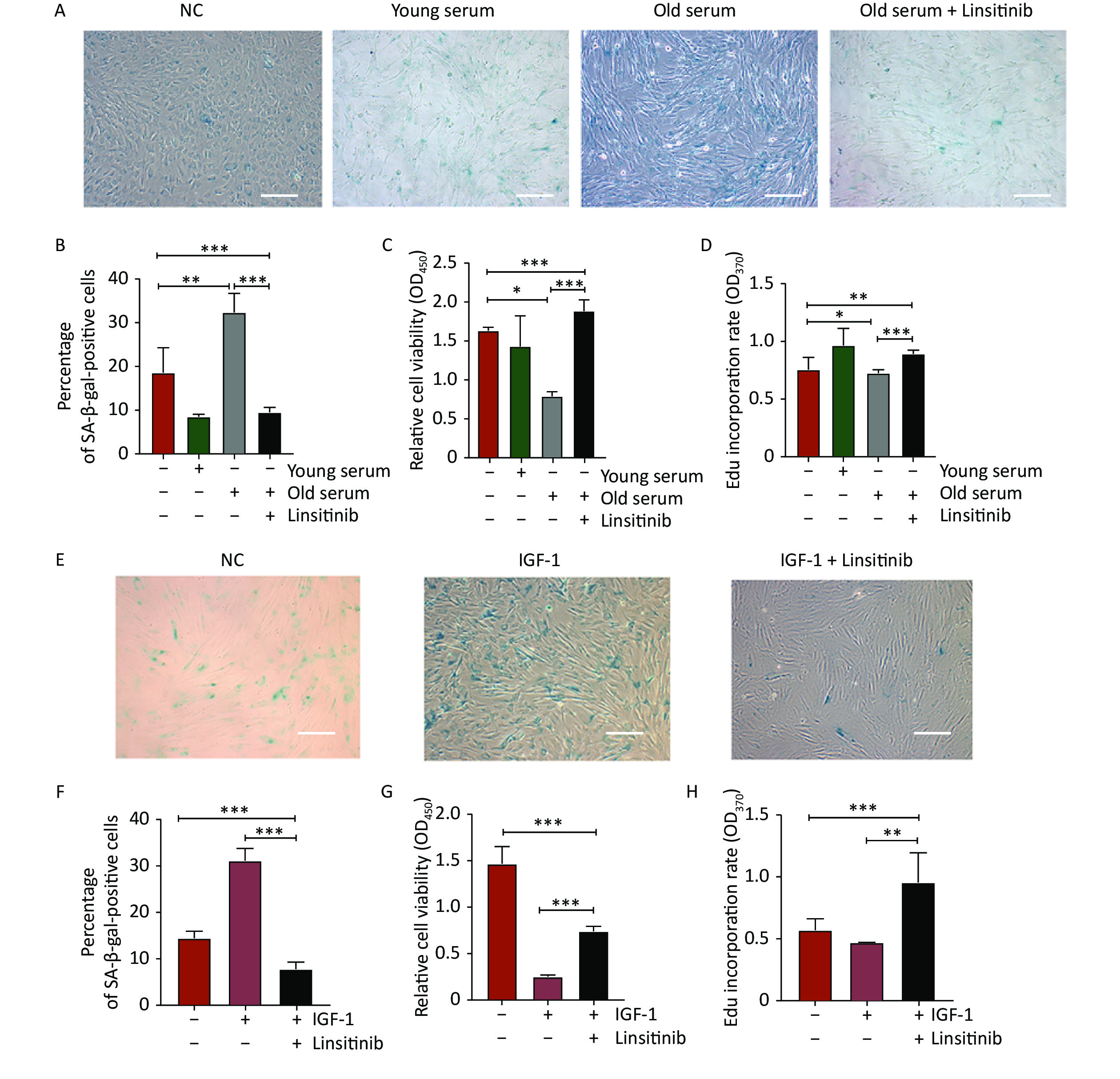
To further explore the effect of IGF-1 on cell aging, the 2BS cells were cultured with medium containing 10% serum from young donors or old donors to detect SA-β-gal positivity, cell proliferation, and DNA synthesis. The percentage of SA-β-Gal positive cells was significantly lower in the cells incubated with the young-age donor serum, but significantly higher in the cells cultured with the old-age donor serum. The increase of the SA-β-gal positive rate induced by old-age donor serum was reversed by treatment with linsitinib (Figure 3A–B). The proliferation capacity increased significantly in the cells cultured with young-age donor serum, but decreased in the cells incubated with old-age donor serum. The decrease of the cell proliferative ability induced by old-age donor serum was observed to be reversed by linsitinib (Figure 3C). The EdU incorporation rate was determined to detect DNA synthesis. The rate of EdU incorporation was higher in the cells cultured with young-age donor serum and lower in the cells incubated with old-age donor serum. Similarly, the reduction of the EdU incorporation rate induced by old-age donor serum could also be reversed by linsitinib (Figure 3D). These results indicate that IGF-1 accelerates the aging of 2BS cells, after considering the different concentrations of IGF-1 in the serum of donors of different ages.
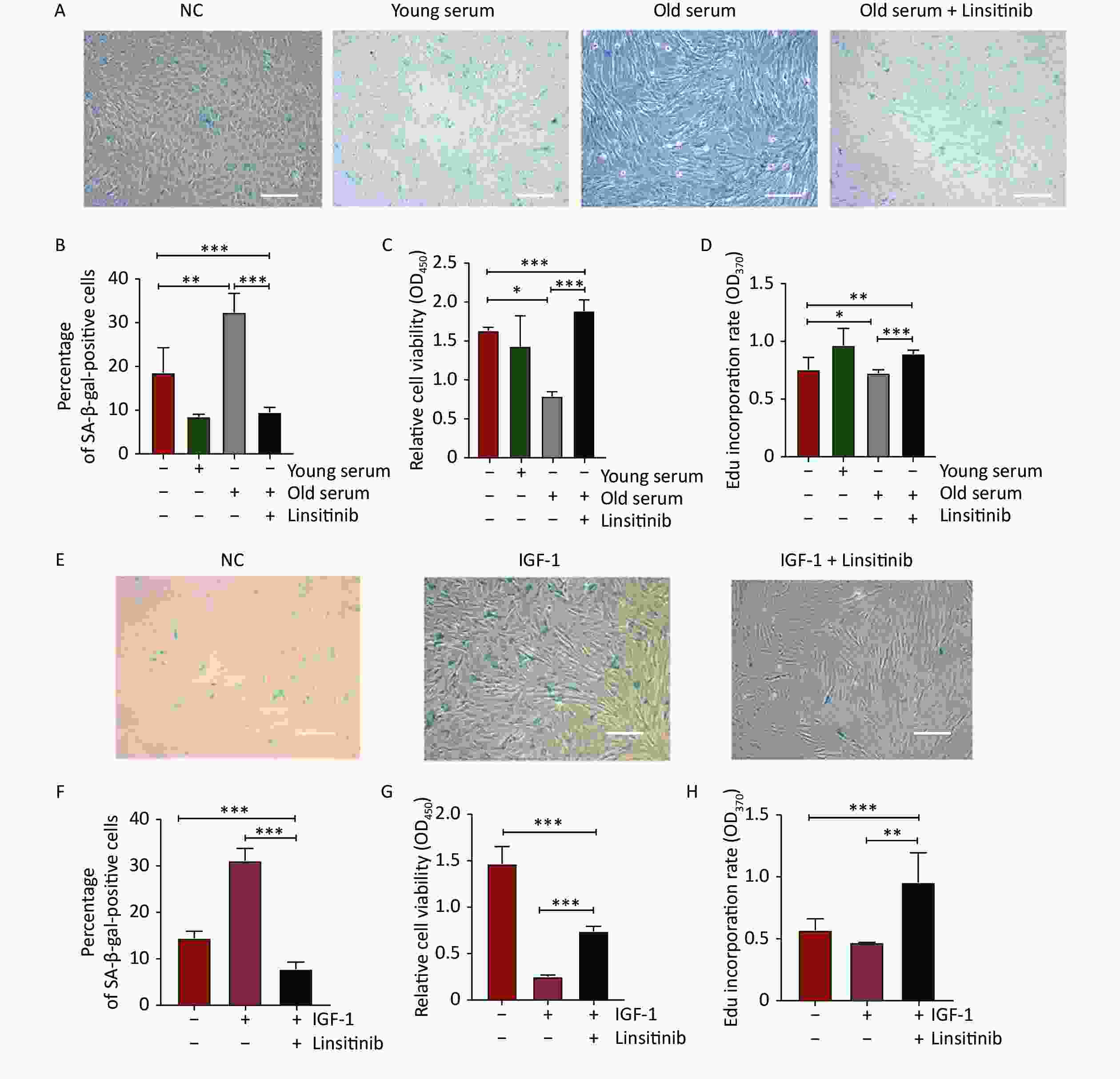
Figure 3. Changes of senescent biomarkers in the 2BS cells incubated with young-age donor serum, old-age donor serum, old-age donor serum + linsitinib, IGF-1 and IGF-1 + linsitinib, respectively. (A–B) The results of SA-β-gal staining of 2BS cells cultured with young-age donor serum, old-age donor serum, and old-age donor serum + linsitinib, respectively (The images were captured with an Olympus IX73 microscope. Scale bars, 50 μm. Magnification, ×100). (C–D) The proliferation capacity and the EdU incorporation rate of 2BS cells cultured with young-age donor serum, old-age donor serum, and old-age donor serum + linsitinib, respectively. (E–F) The results of SA-β-gal staining of 2BS cells incubated with IGF-1 and IGF-1 + linsitinib, respectively (Magnification, ×100). (G–H) The proliferation capacity and the EdU incorproration rate of 2BS cells treated with IGF-1 and IGF-1 + linsitinib, respectively. (Date are shown as the mean ± SEM, with three independent experiments in each group; *P < 0.05, **P < 0.01, ***P < 0.001)
To further verify the effect of IGF-1 on cell aging, the levels of SA-β-gal staining, cell proliferation, and DNA synthesis were determined in 2BS cells treated with IGF-1 and linsitinib. The results showed that the percentage of SA-β-gal positive cells treated with IGF-1 increased significantly, whereas it decreased significantly when the cells were cultured with linsitinib and IGF-1 together (Figure 3E–F). The proliferative capacity and the EdU incorporation rate of 2BS cells treated with IGF-1 were lower than those in cells cultured with medium containing no IGF-1, while they were increased significantly when linsitinib was added into the medium containing IGF-1 (Figure 3G–H).
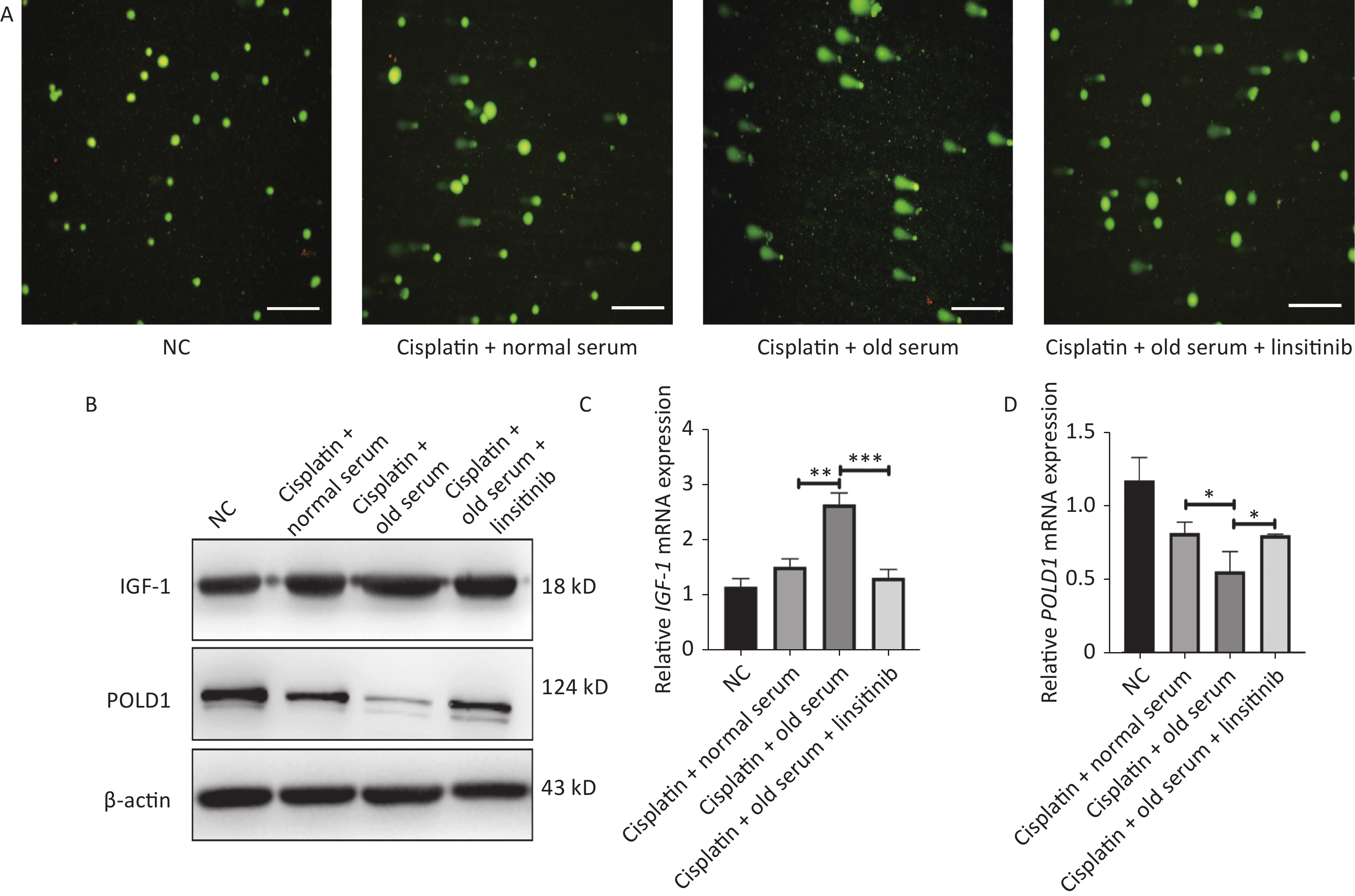
We also measured the effect of IGF-1 on DNA repair. We treated the 2BS cells with cisplatin (50 μmol/L, 24 h) to induce DNA damage, and the expression levels of IGF-1 and POLD1 were detected after treating the stressed cells with old-age donor serum or linsitinib. The comet tail area (expressed as a percentage) represents the level of DNA breakage. Our results showed that the DNA repair ability was lower in cells treated with old-age serum, and this effect was reversed by the addition of linsitinib. Additionally, the IGF-1 expression level was higher and the POLD1 expression level was lower in cells treated with old-age serum, and this effect was reversed by the addition of linsitinib (Supplementary Figure S2, available in
www.besjournal.com ). Taken together, these results demonstrate that IGF-1 promotes aging of 2BS cells.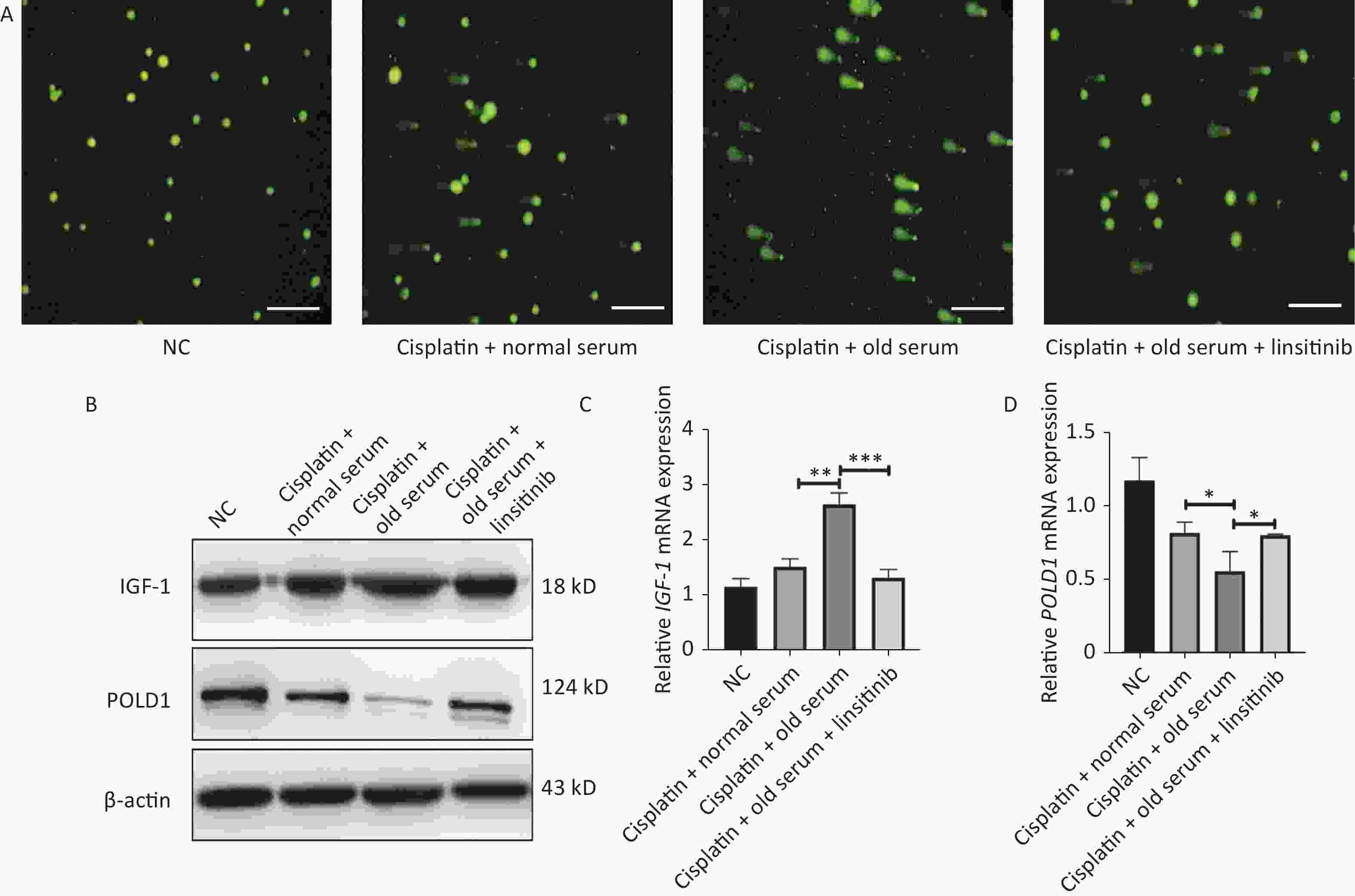
Figure S2. The expression of IGF-1 and POLD1 in the cells treated with old serum or linsitinib after the cispaltin to induce DNA damage. (A) The comet assay to assess DNA repair ability in the cells treating with old serum or linsitinib after the cispaltin to induce DNA damage (Scale bars, 50 μm). (B) The protein expression of IGF-1 and POLD1 in cells treating with old serum or linsitinib after the cispaltin to induce DNA damage. (C–D) The mRNA expression of IGF-1 and POLD1 in cells treating with old serum or linsitinib after the cispaltin to induce DNA damage. (*P < 0.05, **P < 0.01, ***P < 0.001)
-
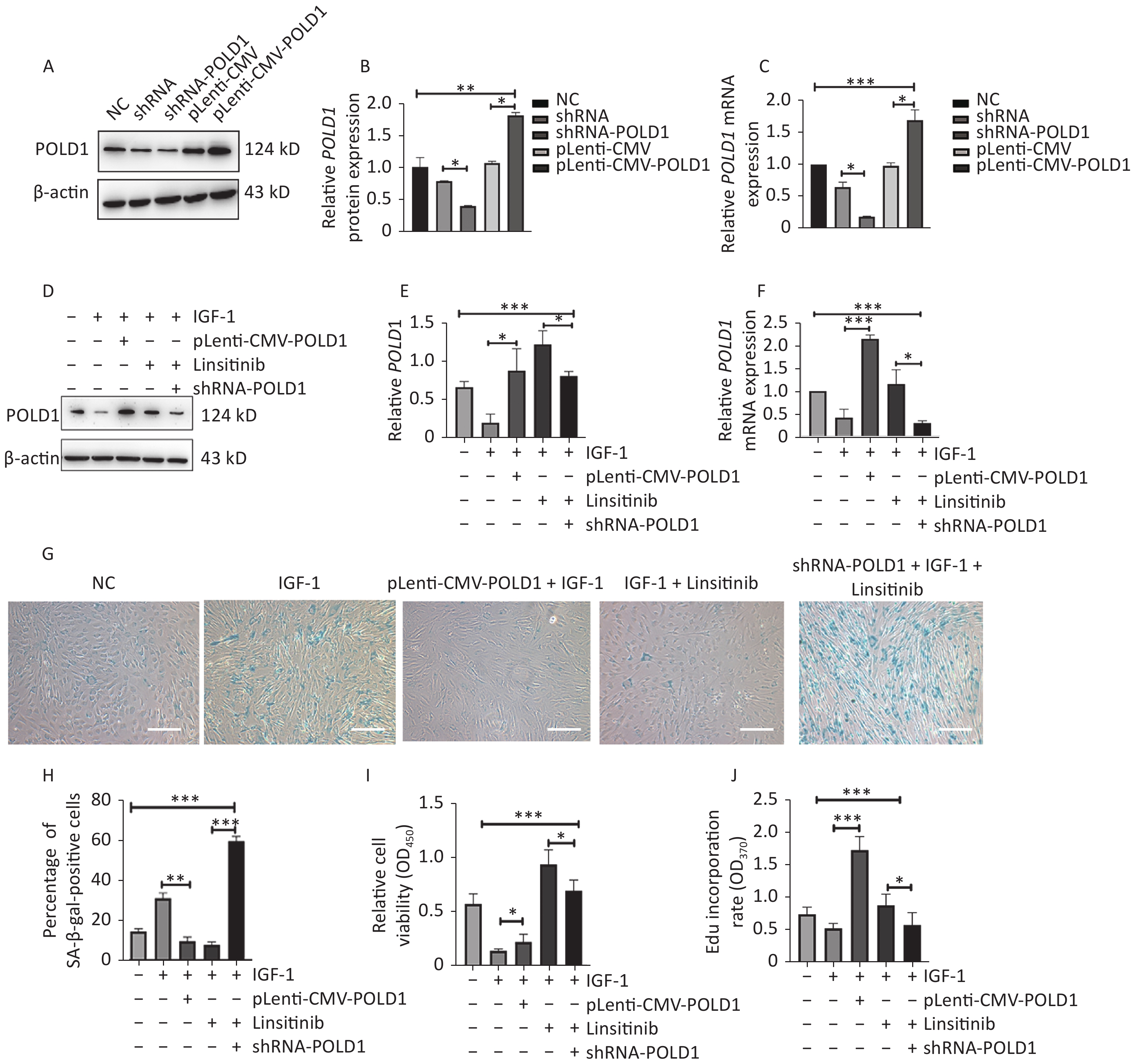
To determine whether IGF-1 promoted aging by targeting POLD1, we generated 2BS cells with increased or decreased POLD1 expression by transfecting pLenti-CMV-POLD1 or shRNA-POLD1, respectively (Figure 4A–C). Lower POLD1 expression was observed in the cells cultured with IGF-1, whereas higher expression was noted when linsitinib was added. Compared with control cells, higher POLD1 expression was observed in the cells transfected with pLenti-CMV-POLD1 when IGF-1 was added. Moreover, decreased POLD1 expression was noted in the shRNA-POLD1-transfected cells when IGF-1 and linsitinib were added (Figure 4D–F).
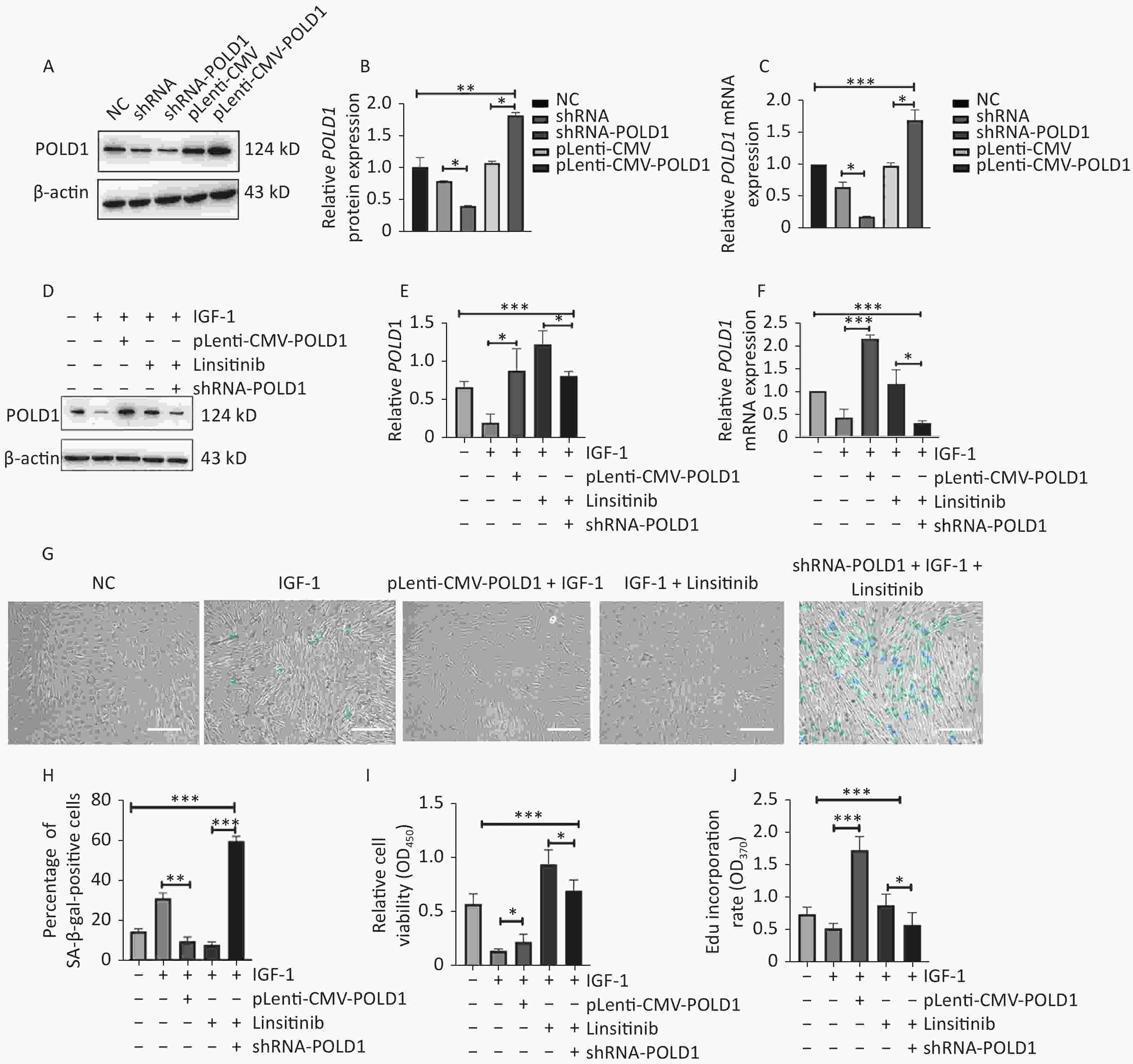
Figure 4. Changes of senescent biomarkers in the 2BS cells treated with IGF-1, IGF-1 + pLenti-CMV-POLD1, IGF-1 + linsitinib, and IGF-1 + linsitinib + shRNA-POLD1, respectively. (A–B) The protein expression level of POLD1 in 2BS cells. (C) The mRNA expression level of POLD1 in 2BS cells. (D–E) The protein expression of POLD1 in 2BS cells with different treatments. (F) The mRNA expression of POLD1 in 2BS cells with different treatments. (G) SA-β-gal staining of 2BS cells with different treatments (The images were captured with an Olympus IX73 microscope. Scale bars, 50 μm. Magnification, ×100). (H) The percentage of SA-β-gal-positive cells counted after each treatment. (I) The proliferation capacity of 2BS cells with different treatments measured by the CCK8 assay. (J) The EdU incorporation rate of the cells after different treatments. (Data are shown as the mean ± SEM, with three independent experiments in each group; *P < 0.05, **P < 0.01, ***P < 0.001).
Compared with control cells, the SA-β-Gal positive rate was higher in the cells cultured with IGF-1, but lower in the pLenti-CMV-POLD1-transfected cells incubated with IGF-1. Furthermore, the SA-β-Gal positive rate was increased significantly in the shRNA-POLD1-transfected cells when cultured with IGF-1 and linsitinib (Figure 4G–H). The proliferative capacity and the EdU incorporation rate were lower in the cells cultured with IGF-1; however, they were elevated significantly in the cells transfected with pLenti-CMV-POLD1 when cultured with IGF-1. The proliferative capacity and the EdU incorporation rate of the cells treated with linsitinib were reduced after transfection with shRNA-POLD1 (Figure 4I–J). These results demonstrate that IGF-1 promotes cell aging by inhibiting POLD1 expression.
-
As a key mediator of the growth hormone pathway, IGF-1 plays a vital role in cell development. It had been reported to modulate aging in many kinds of organisms, including fruit flies, nematodes, and mice, although the results on the role of IGF-1 in aging are contradictory[19–23]. In genetically-modified knockdown and knockout models, down-regulation of the GH/IGF-1 signaling pathway generally improves the health of mice and reduces age-associated pathologies such as immunosenescence and cancer[24]. In addition, previous studies have demonstrated that the GH/IGF-1 axis plays a critical role in the repair of DNA damage in cancer cells by altering DNA repair-related genes at an early stage[25–27].
POLD1, as the central mediator of DNA replication and DNA damage repair, has been reported to play an essential role in cell senescence. It can inhibit cell senescence by promoting cell proliferation, DNA synthesis, and DNA repair[28]. In the present study, a negative correlation between the levels of IGF-1 and POLD1 was demonstrated. In serum specimens from healthy donors and hippocampal tissues from SAMP8 mice, an age-related increase of IGF-1 and an age-associated decline of POLD1 were observed. To illustrate the effect of IGF-1 on the expression of POLD1, 2BS cells, cultured with serum from either young or old donors, were used to examine POLD1 expression. The results showed that the serum of young donors, which had a lower concentration of IGF-1, could up-regulate the POLD1 expression level. By contrast, the serum of old donors, which had a higher level of IGF-1, could down-regulate the POLD1 expression level, and the POLD1 level could be reversed by treatment with an IGF-1 receptor inhibitor (linsitinib). In addition, it was observed that POLD1 expression could be down-regulated in 2BS cells cultured in the presence of IGF-1, and likewise, the POLD1 level could be reversed by treatment with linsitinib. These results demonstrate that IGF-1 inhibits the expression of POLD1 by binding to IGF-1R, although the details of this mechanism remain unclear.
Although the effect of IGF-1 on aging has been verified in previous studies, its effect on cellular senescence was observed in this study. A higher percentage of SA-β-gal-positive cells was observed in the cells cultured with old-age donor serum or IGF-1. By contrast, young-age donor serum or linsitinib reduced the percentage of SA-β-gal-positive cells. A strikingly lower rate of cell proliferation and EdU incorporation was observed in the cells cultured with old-age donor serum or IGF-1. In addition, significant increases in the rate of cell proliferation and the rate of EdU incorporation were detected in the cells incubated with young-age donor serum or linsitinib. Furthermore, the ability of cells to induce DNA repair was decreased and the expression of IGF-1 increased in the cells incubated with old-age donor serum. These results indicate that IGF-1 promotes the aging of 2BS cells by inhibiting DNA synthesis, reducing cell proliferation capacity, and decreasing DNA repair ability.
As discussed above, DNA polymerase δ is responsible for DNA replication and DNA damage repair. To investigate the mechanism of IGF-1 in promoting aging, the role of POLD1 in the senescence of IGF-1-treated 2BS cells was examined. The percentage of SA-β-gal-positive cells incubated with IGF-1 was reduced in pLenti-CMV-POLD1-transfected cells and elevated in shRNA-POLD1-transfected 2BS cells treated with linsitinib. Moreover, the rate of proliferation and the rate of EdU incorporation of the IGF-1-treated cells were increased in pLenti-CMV-POLD1-transfected cells and decreased in shRNA-POLD1-transfected cells. These results verify that IGF-1 promotes cell aging by inhibiting POLD1-mediated DNA synthesis and cell proliferation.
With regard to the mechanism of the IGF-1R complex in regulating POLD1 expression, no direct evidence was reported. A recent study has provided very strong support for the role of the IGF-1 signaling pathway in the control of aging via InRβ, IRS-1 or IRS-2, and PI3K, which can affect genes involved in the control of longevity in mammals[25]. We assume that IGF-1 affects the expression of POLD1 through some pathway, although the specific mechanism needs to be verified in the future.
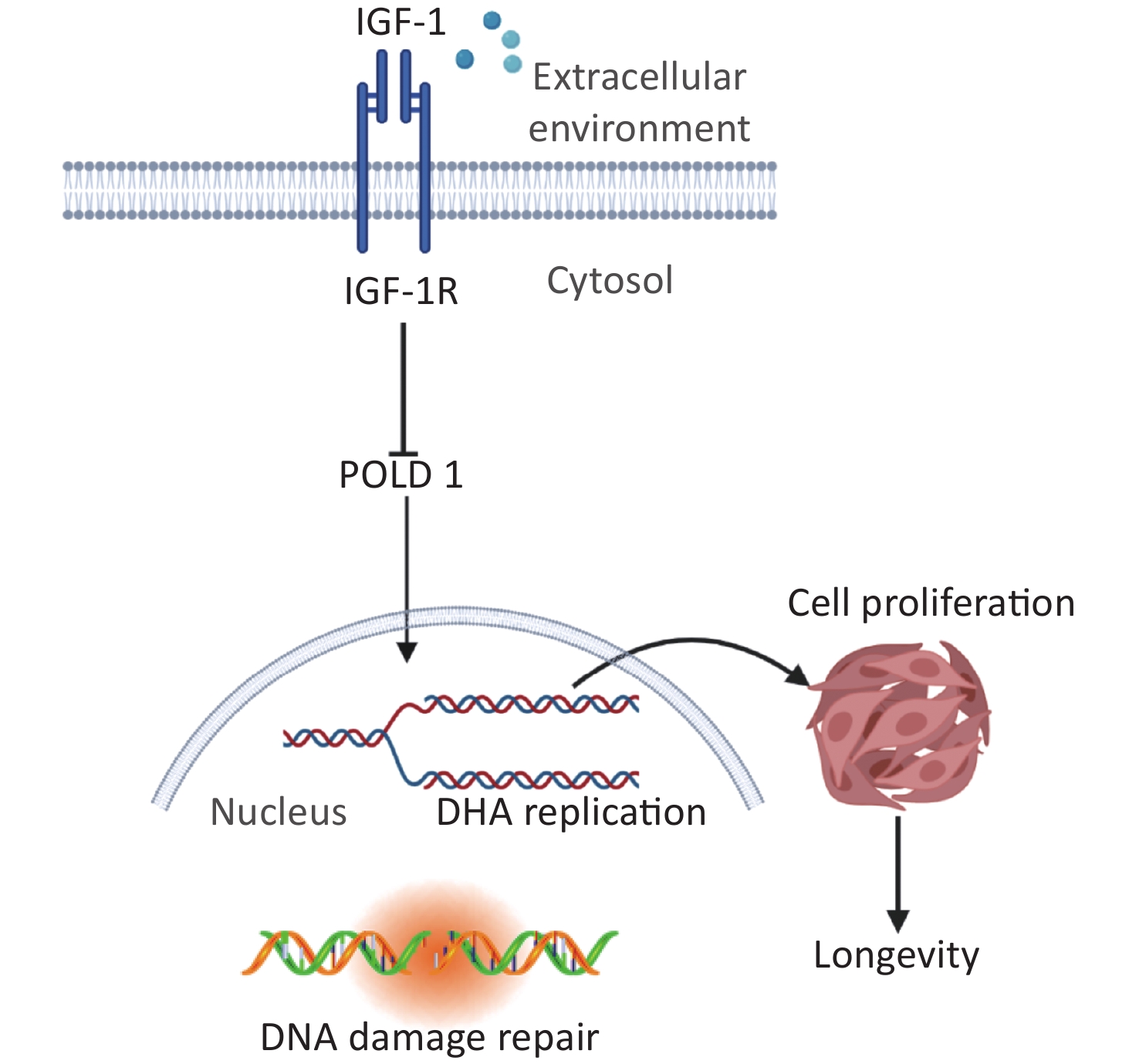
This study offers insight into the regulatory mechanism of IGF-1 in the aging process. We propose that IGF-1 up-regulation can inhibit the expression of POLD1 by binding to IGF-1R, which in turn suppresses cell proliferation and DNA synthesis (Figure 5). We believe that regulating the levels of IGF-1 can significantly delay aging and further in-depth exploration is warranted.
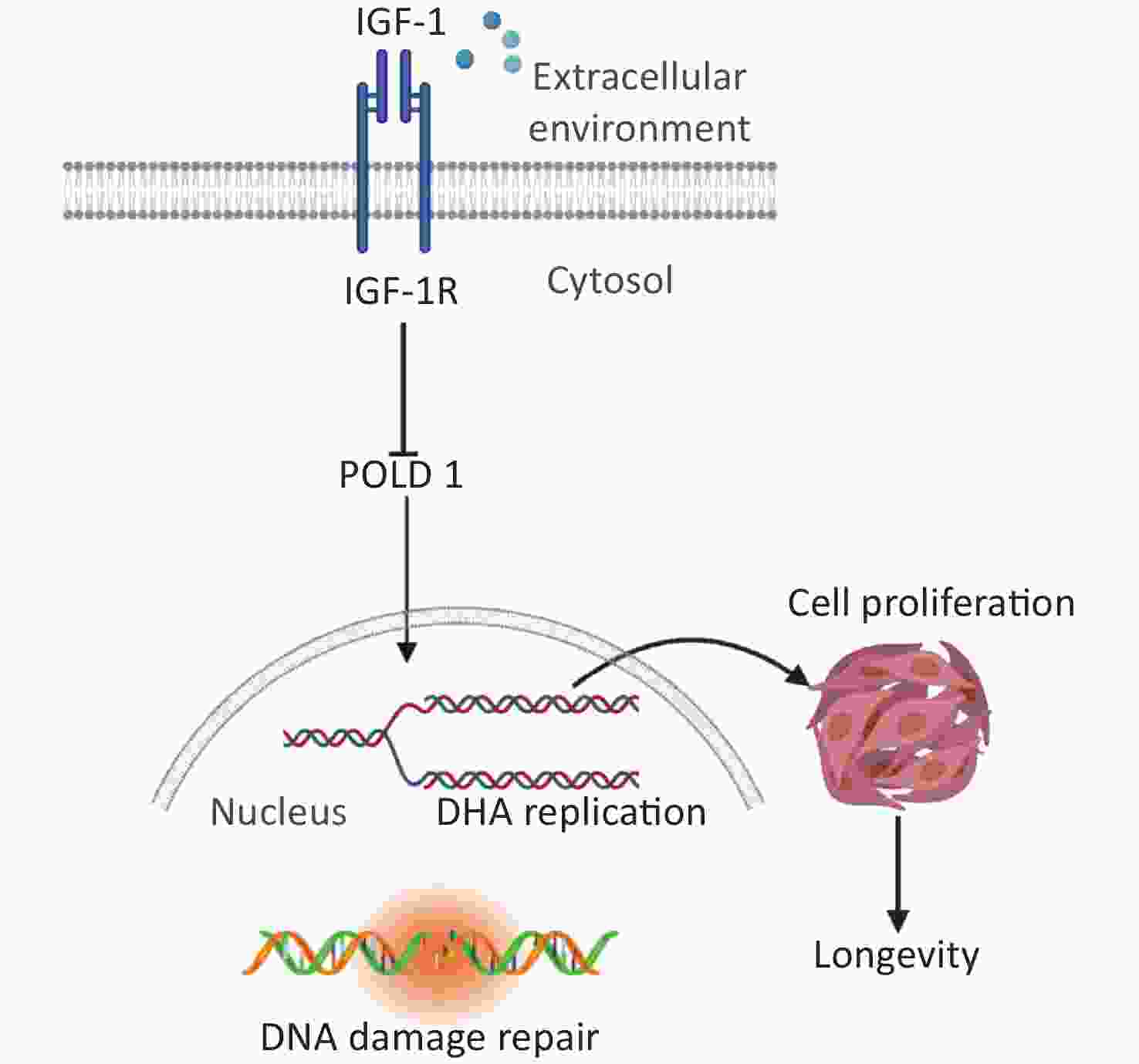
Figure 5. Schematic drawing of the regulatory mechanism of IGF-1 on cell aging.
-
HOU Yu Li performed the experiments and wrote the first draft of the manuscript. WANG Yi Fei, SONG Qiao, and ZHANG Xiao Min collected the data. LIU Jing and WANG Ya Qi provided methodological guidance. CUI Yu Ting, FU Jing Xuan, FENG Zi Yi, and ZHANG Chi provided the clinical specimens. WANG Pei Chang reviewed the manuscript and provided the funding.
-
All procedures adhered to the ethical standards of the responsible committee on human experimentation (institutional and national) and to the tenets of the Helsinki Declaration of 1975, as revised in 2000.
doi: 10.3967/bes2022.127
-
Abstract:
Objective The individual cascades of the insulin-like growth factor-1 (IGF-1) signaling pathway and the molecular mechanism of aging have not been fully clarified. In the current study, we explored the effect of DNA polymerase delta 1 (POLD1) on the IGF-1 signaling pathway in cell aging. Methods First, we analyzed the relationship between IGF-1 and POLD1 expression in aging. To investigate the effect of IGF-1 on POLD1 expression and aging, the 2BS cells were incubated with young-age or old-age human serum, IGF-1 protein, or linsitinib. Next, the effect of IGF-1 on aging was examined in the 2BS cells with increased or decreased POLD1 expression to clarify the molecular mechanism. Results In this study, we found that IGF-1 expression increased and POLD1 expression decreased with aging in human serum and hippocampal tissues of SAMP8 mice, and a negative relationship between IGF-1 and POLD1 expression was observed. Furthermore, the cells cultured with old-age human serum or IGF-1 showed lower POLD1 expression and more pronounced senescence characteristics, and the effect could be reversed by treatment with linsitinib or overexpression of POLD1, while the effect of linsitinib on cell aging could be reversed with the knockdown of POLD1. Conclusion Taken collectively, our findings demonstrate that IGF-1 promotes aging by binding to IGF-1R and inhibiting the expression of POLD1. These findings offer a new target for anti-aging strategies. -
Key words:
- Insulin-like growth factor-1 /
- DNA polymerase delta 1 /
- Aging /
- Linsitinib
注释:1) CONFLICTS OF INTEREST: -
Figure 1. IGF-1 and POLD1 levels in the serum from healthy individuals of different ages and the hippocampal tissues from young and old SAMP8 mice. (A) IGF-1 concentrations in the human serum of different ages. (B) The concentrations of POLD1 in the human serum of different ages. (C) The relationship between IGF-1 and POLD1 concentrations in the human serum of different ages. (D–F) The protein expression levels of IGF-1 and POLD1 in the hippocampus of young and old SAMP8 mice. (G) The relationship between IGF-1 and POLD1 protein levels in the hippocampus of SAMP8 mice. (H–I) IGF-1 and POLD1 mRNA expression levels in the hippocampus of young and old SAMP8 mice. (J) The relationship between IGF-1 and POLD1 mRNA levels in the hippocampus of SAMP8 mice. (20–29, n = 35; 40–49, n = 33; 60–69, n = 30; 80–89, n = 24; young mice, 2 months, n = 6; old mice, 8 months, n = 6; *P < 0.05, ***P < 0.001).
S1. The expression of IGF-1 and POLD1 in human serum from male and female people. (A) IGF-1 concentrations in the human serum from male and female people. (B) The concentrations of POLD1 in the human serum from male and female people. (ns P > 0.05).
Figure 2. Expression levels of POLD1 and IGF-1 in 2BS cells incubated with young-age donor serum, old-age donor serum, old-age donor serum + linsitinib, and IGF-1 and IGF-1 + linsitinib, respectively. (A–D) The protein expression levels of IGF-1, IGF-1R, and pIGF-1R in 2BS cells cultured with IGF-1 or linsitinib and IGF-1 + linsitinib (#IGF-1 vs. NC, P < 0.05; ###IGF-1 vs. NC, P < 0.001; &IGF-1 + linsitinib vs. IGF-1, P < 0.05). (E–I) The protein and mRNA expression levels of IGF-1 and POLD1 in 2BS cells cultured with young-age donor serum, old-age donor serum, and old-age donor serum + linsitinib, respectively. (J–N) The protein and mRNA expression levels of IGF-1 and POLD1 in 2BS cells incubated with IGF-1 or IGF-1 + linsitinib. (Date are shown as the mean ± SEM, with three independent experiments in each group; ns P > 0.05, *P < 0.05, ***P < 0.001).
Figure 3. Changes of senescent biomarkers in the 2BS cells incubated with young-age donor serum, old-age donor serum, old-age donor serum + linsitinib, IGF-1 and IGF-1 + linsitinib, respectively. (A–B) The results of SA-β-gal staining of 2BS cells cultured with young-age donor serum, old-age donor serum, and old-age donor serum + linsitinib, respectively (The images were captured with an Olympus IX73 microscope. Scale bars, 50 μm. Magnification, ×100). (C–D) The proliferation capacity and the EdU incorporation rate of 2BS cells cultured with young-age donor serum, old-age donor serum, and old-age donor serum + linsitinib, respectively. (E–F) The results of SA-β-gal staining of 2BS cells incubated with IGF-1 and IGF-1 + linsitinib, respectively (Magnification, ×100). (G–H) The proliferation capacity and the EdU incorproration rate of 2BS cells treated with IGF-1 and IGF-1 + linsitinib, respectively. (Date are shown as the mean ± SEM, with three independent experiments in each group; *P < 0.05, **P < 0.01, ***P < 0.001)
S2. The expression of IGF-1 and POLD1 in the cells treated with old serum or linsitinib after the cispaltin to induce DNA damage. (A) The comet assay to assess DNA repair ability in the cells treating with old serum or linsitinib after the cispaltin to induce DNA damage (Scale bars, 50 μm). (B) The protein expression of IGF-1 and POLD1 in cells treating with old serum or linsitinib after the cispaltin to induce DNA damage. (C–D) The mRNA expression of IGF-1 and POLD1 in cells treating with old serum or linsitinib after the cispaltin to induce DNA damage. (*P < 0.05, **P < 0.01, ***P < 0.001)
Figure 4. Changes of senescent biomarkers in the 2BS cells treated with IGF-1, IGF-1 + pLenti-CMV-POLD1, IGF-1 + linsitinib, and IGF-1 + linsitinib + shRNA-POLD1, respectively. (A–B) The protein expression level of POLD1 in 2BS cells. (C) The mRNA expression level of POLD1 in 2BS cells. (D–E) The protein expression of POLD1 in 2BS cells with different treatments. (F) The mRNA expression of POLD1 in 2BS cells with different treatments. (G) SA-β-gal staining of 2BS cells with different treatments (The images were captured with an Olympus IX73 microscope. Scale bars, 50 μm. Magnification, ×100). (H) The percentage of SA-β-gal-positive cells counted after each treatment. (I) The proliferation capacity of 2BS cells with different treatments measured by the CCK8 assay. (J) The EdU incorporation rate of the cells after different treatments. (Data are shown as the mean ± SEM, with three independent experiments in each group; *P < 0.05, **P < 0.01, ***P < 0.001).
Table 1. Characteristics of donors in different ages
Characteristic 20–29 years (n = 35) 40–49 years (n = 33) 60–69 years (n = 30) 80–89 years (n = 24) Gender Male (n) 11 15 16 11 Female (n) 24 18 14 13 Age Mean ± SEM 24.76 ± 1.72 45.04 ± 1.41 64.54 ± 1.53 86.44 ± 3.34 Min-Max 21–29 43–47 62–68 81–89  下载: 导出CSV
下载: 导出CSV
Table 2. Antibodies used for Western blotting
Protein Manufacturer Cat no. Primary antibodies IGF-1 Abclonal Technology
Co. /ChinaA0830 POLD1 Abcam/UK Ab186407 β-actin Zhongshan Boil Tech
Co. /ChinaTA-09 Secondary antibodies HRP-conjugated
anti-mouse IgGZhongshan Boil Tech
Co. /ChinaZB2305 HRP-conjugated
anti-rabbit IgGZhongshan Boil Tech
Co. /ChinaZB5301
下载: 导出CSV
Table 3. Sequences used for quantitative real-time polymerase chain reaction
Gene Sequences β-actin Forward ACAGAGCCTCGCCTTTGC Reverse CCACCATCACGCCCTGG IGF-1 Forward TGCTTCCGGAGCTGTGATCT Reverse TCTGGGTCTTGGGCATGTC POLD1 Forward GCTCCGCTCCTACACGCTCAA Reverse GGTCTGGTCGTTCCCATTCTGC
下载: 导出CSV
-
[1] Li ZW, Wang DD, Chen XT, et al. PRMT1-mediated EZH2 methylation promotes breast cancer cell proliferation and tumorigenesis. Cell Death Dis, 2021; 12, 1080. doi: 10.1038/s41419-021-04381-5 [2] Palmer AK, Gustafson B, Kirkland JL, et al. Cellular senescence: at the nexus between ageing and diabetes. Diabetologia, 2019; 62, 1835−41. doi: 10.1007/s00125-019-4934-x [3] Evans MA, Sano S, Walsh K. Cardiovascular disease, aging, and clonal hematopoiesis. Annu Rev Pathol Mech Dis, 2020; 15, 419−38. doi: 10.1146/annurev-pathmechdis-012419-032544 [4] Mathys H, Davila-Velderrain J, Peng ZY, et al. Single-cell transcriptomic analysis of Alzheimer's disease. Nature, 2019; 570, 332−7. doi: 10.1038/s41586-019-1195-2 [5] Sharpless NE, Sherr CJ. Forging a signature of in vivo senescence. Nat Rev Cancer, 2015; 15, 397−408. doi: 10.1038/nrc3960 [6] Shay JW. Role of telomeres and telomerase in aging and cancer. Cancer Discov, 2016; 6, 584−93. doi: 10.1158/2159-8290.CD-16-0062 [7] Gao SC, Zhang XM, Song Q, et al. POLD1 deficiency is involved in cognitive function impairment in AD patients and SAMP8 mice. Biomed Pharmacother, 2019; 114, 108833. doi: 10.1016/j.biopha.2019.108833 [8] Mchugh D, Gil J. Senescence and aging: Causes, consequences, and therapeutic avenues. J Cell Biol, 2018; 217, 65−77. doi: 10.1083/jcb.201708092 [9] Junnila RK, List EO, Berryman DE, et al. The GH/IGF-1 axis in ageing and longevity. Nat Rev Endocrinol, 2013; 9, 366−76. doi: 10.1038/nrendo.2013.67 [10] Holzenberger M, Dupont J, Ducos B, et al. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature, 2003; 421, 182−7. doi: 10.1038/nature01298 [11] Mattison JA, Roth GS, Beasley TM, et al. Impact of caloric restriction on health and survival in rhesus monkeys from the NIA study. Nature, 2012; 489, 318−21. doi: 10.1038/nature11432 [12] Green CL, Lamming DW, Fontana L. Molecular mechanisms of dietary restriction promoting health and longevity. Nat Rev Mol Cell Biol, 2022; 23, 56−73. doi: 10.1038/s41580-021-00411-4 [13] Flurkey K, Papaconstantinou J, Miller RA, et al. Lifespan extension and delayed immune and collagen aging in mutant mice with defects in growth hormone production. Proc Natl Acad Sci USA, 2001; 98, 6736−41. doi: 10.1073/pnas.111158898 [14] Duran-Ortiz S, List EO, Basu R, et al. Extending lifespan by modulating the growth hormone/insulin-like growth factor-1 axis: coming of age. Pituitary, 2021; 24, 438−56. doi: 10.1007/s11102-020-01117-0 [15] Aguiar-Oliveira MH, Bartke A. Growth hormone deficiency: health and longevity. Endocr Rev, 2019; 40, 575−601. doi: 10.1210/er.2018-00216 [16] Murdocca M, Spitalieri P, De Masi C, et al. Functional analysis of POLD1 p. ser605del variant: the aging phenotype of MDPL syndrome is associated with an impaired DNA repair capacity. Aging (Albany NY), 2021; 13, 4926−45. [17] Gao SC, Song Q, Liu J, et al. E2F1 mediates the downregulation of POLD1 in replicative senescence. Cell Mol Life Sci, 2019; 76, 2833−50. doi: 10.1007/s00018-019-03070-z [18] Hou YL, Song Q, Gao SC, et al. CTCF mediates replicative senescence through POLD1. Front Cell Dev Biol, 2021; 9, 618586. doi: 10.3389/fcell.2021.618586 [19] Kenyon C, Chang J, Gensch E, et al. A C. elegans mutant that lives twice as long as wild type. Nature, 1993; 366, 461−4. doi: 10.1038/366461a0 [20] Lee SH, Lee HY, Yu M, et al. Extension of Drosophila lifespan by Korean red ginseng through a mechanism dependent on dSir2 and insulin/IGF-1 signaling. Aging (Albany NY), 2019; 11, 9369−87. [21] Duran-Ortiz S, List EO, Ikeno Y, et al. Growth hormone receptor gene disruption in mature-adult mice improves male insulin sensitivity and extends female lifespan. Aging Cell, 2021; 20, e13506. [22] Rahmani J, Montesanto A, Giovannucci E, et al. Association between IGF-1 levels ranges and all-cause mortality: A meta-analysis. Aging Cell, 2022; 21, e13540. [23] Grigolon G, Araldi E, Erni R, et al. Grainyhead 1 acts as a drug-inducible conserved transcriptional regulator linked to insulin signaling and lifespan. Nat Commun, 2022; 13, 107. doi: 10.1038/s41467-021-27732-4 [24] Spadaro O, Goldberg EL, Camell CD, et al. Growth hormone receptor deficiency protects against age-related NLRP3 inflammasome activation and immune senescence. Cell Rep, 2016; 14, 1571−80. doi: 10.1016/j.celrep.2016.01.044 [25] Plummer JD, Postnikoff SD, Tyler JK, et al. Selenium supplementation inhibits IGF-1 signaling and confers methionine restriction-like healthspan benefits to mice. eLife, 2021; 10, e62483. doi: 10.7554/eLife.62483 [26] Podlutsky A, Valcarcel-Ares MN, Yancey K, et al. The GH/IGF-1 axis in a critical period early in life determines cellular DNA repair capacity by altering transcriptional regulation of DNA repair-related genes: implications for the developmental origins of cancer. Geroscience, 2017; 39, 147−60. doi: 10.1007/s11357-017-9966-x [27] Poreba E, Durzynska J. Nuclear localization and actions of the insulin-like growth factor 1 (IGF-1) system components: Transcriptional regulation and DNA damage response. Mutat Res Rev Mutat Res, 2020; 784, 108307. doi: 10.1016/j.mrrev.2020.108307 [28] Layer JV, Debaize L, Van Scoyk A, et al. Polymerase δ promotes chromosomal rearrangements and imprecise double-strand break repair. Proc Natl Acad Sci USA, 2020; 117, 27566−77. doi: 10.1073/pnas.2014176117 -
 22145Supplementary Materials.pdf
22145Supplementary Materials.pdf

-

 点击查看大图
点击查看大图
计量
- 文章访问数: 1964
- HTML全文浏览量: 869
- PDF下载量: 202
- 被引次数: 0

 Quick Links
Quick Links