

 下载:
下载:




-
Most cervical cancers could be prevented with human papillomavirus (HPV) vaccines, routine cervical screenings, and prompt treatment for precancerous lesions. However, screening programs worldwide are inadequate, and cervical cancer remains the fourth most common malignancy diagnosed among women globally [1, 2]. Over the past 20 years, the incidence and mortality rates of cervical cancer in women in China have increased every year [3]. Although traditional treatments such as surgery and chemoradiation are widely utilized in patients with cervical cancer, effective treatment options remain limited. Molecular targeted therapy is considered a potential treatment for cervical cancer because of its effectiveness and fewer side effects than traditional therapies. At present, targeted drugs for cervical cancer, such as the EGFR-targeting drug gefitinib, have greatly reformed the current treatment strategy, and targeted VEGF drugs, including Bevacizumab and Pazopanib, are in preclinical studies [4]. Searching for new oncogenic drivers in cervical cancer has become important for the development of targeted therapy.
Splicing regulator protein transformer 2β (Tra2β), which contains two ribonucleoprotein domains and serine/arginine-rich domains, was originally identified as a new putative RNA-binding protein (RA301) in rat reoxygenated astrocytes, which is currently recognized as a sequence-specific pre-mRNA splicing enhancer [5]. It binds to the exonic splicing enhancer sequence, which is often found in an A/G-rich context, thereby modulating constitutive and alternative splicing of pre-mRNAs [6]. Accumulating body of evidence suggests that the RNA-binding protein Tra2β is deregulated in various malignant tumors, including colon cancer [7], breast cancer [8], endometrial carcinoma [9], lung squamous cell carcinoma [10], and prostate cancer [11]. Boris et al. showed that among 105 patients with cervical cancer, patients with high nuclear expression levels of Tra2β had poor prognoses compared with patients with weak amounts of nuclear Tra2β. However, the critical role of Tra2β in carcinogenesis remains elusive [12]. In this study, we primarily focused on the role of the splicing regulator Tra2β as a gene modulator in cervical cancer cells and provided a theoretical basis for potential targeted therapy for cervical cancer.
-
Gene Expression Profiling Interactive Analysis (GEPIA,
http://gepia.cancer-pku.cn/ ) was utilized to analyze the Tra2β expression level and correlation between Tra2β expression and tumor stage in patients with cervical cancer from a TCGA cohort. cBioPortal databases (http://www.cbioportal.org) were used to investigate the Tra2β gene expression, mutation, and correlation among EGR1, SP1, NOLC1, and Tra2β in CESC. Cervical cancer microarray data obtained from GSE7803 (Platform: GPL96) and GSE9750 (Platform: GPL96) were used to analyze the SP1 expression. -
Human normal cervical epithelial cells (HcerEpic) and human cervical cancer cell lines (Hela and Siha cells) were obtained from the Cell Bank of Chinese Academy of Sciences (Shanghai, China) and cultured in RPMI-1640 (Sigma-Aldrich, St Louis, USA) medium containing 10% fetal bovine serum (Gibco, Grand Island, NY, USA) at 37 ℃ in a humidified 5% CO2 incubator. All the cell lines used in this study were free of mycoplasma contamination.
-
Full-length Tra2β1 cDNA (human) was granted by Han Jiahuai Lab (Xiamen University, Xiamen, China) and then cloned into the pLVX-EGFP-IRES-puro lentiviral vector. 293T cells were transfected with the recombinant lentiviral plasmids, psPAX2 packaging plasmids, and pMD2.G envelope plasmids. After incubating for 48 h, cell media were collected and used to infect Hela and Siha cells at 30% confluence. After incubating for 24 h, the virus-containing media were replaced by fresh media. After an additional 48 h, fresh media containing puromycin (2 μg/mL, Sigma) were added to select infected cells for stable Tra2β expression. The mRNA or protein level of the selected cells was subsequently assessed by q-PCR or Western blot.
-
The sequences of the small interfering RNAs (siRNAs) used in this study are as follows: Tra2β-siRNA-1: 5′-AGCUAAAGAACGUGCCAAUTT-3′, Tra2β-siRNA-2: 5′-CCGAUGUGUCUAUUGUAUATT-3′; SP1-siRNA: 5′-GCCAATAGCTACTCAACTA′; si-NC: 5′-AGUCUCCACGUGUACGUTT-3′ (Genepharma, Suzhou, China). Hela or Siha cells were cultured in six-well plates (6 × 105 cells/well), reaching 30% to 50% confluence before transfection. Then, the cells were transfected with 100 nmol/L of Tra2β siRNAs, SP1 siRNAs, or non-target siRNAs using Lipofectamine™ 2000 (Invitrogen, USA) in a serum-free medium. After incubation for 6 h, the medium was replaced by fresh medium containing 10% fetal bovine serum, and the cells were incubated for another 24 h, followed by Western blot, MTT, colony formation, and Transwell assay.
-
A total of 3 × 104 Hela or Siha cells stably overexpressing Tra2β or transfected with siRNAs targeting Tra2β1 were seeded in RPMI-1640 medium in 96-well plates at 0.1 mL/well. MTT staining was used to determine the absorbance at 570 nm and assess cell viability.
-
A total of 1,000 Hela or Siha cells stably overexpressing Tra2β or transfected with siRNAs targeting Tra2β1 were seeded in six-well plates for 2 weeks. After 2 weeks, the cells were rinsed with phosphate-buffered saline (PBS), fixed with 4% methanol for 10 min, and stained with crystal violet solution for 5 min. Then, the crystal violet was removed, and the wells were rinsed, dried, and scanned.
-
Western blot analysis was performed as previously described [13]. Total protein was extracted from cells or tumor xenograft tissues using a total protein extraction kit (Abcam, USA) according to the manufacturer’s instructions. The BCA protein assay kit (Thermo Fisher Scientific, USA) was used to determine protein concentration. The following antibodies were used for staining: rabbit-Tra2β (Proteintech, USA), rabbit-GAPDH (Proteintech, USA), and rabbit-SP1 (Abcam, USA).
-
Total RNA was extracted from Hela or Siha cells using a total RNA extraction kit (Synthgene Biotech., China) according to the manufacturer’s instructions and then transcribed into complementary DNA by using TransScript® one-step gDNA removal and cDNA synthesis supermix (TransGen, China). Quantitative real-time PCR reactions were performed using the PerfectStart® Green qPCR SuperMix (TransGen, China) on the StepOneTM Real-Time PCR System (Applied Biosystems, USA). The primers used were as follows: Tra2β1 (F 5′-ATGACGACGATAAGAGCCCG-3′, R 5′-TCCGTGAGCACTTCCACTTC-3′), SP1 (F 5′-GTGGAGGCAACATCATTGCTG-3′, R 5′-GCCACTGGTACATTGGTCACAT-3′), NOLC1 (F 5′-AGCTGGCCTGACGGTATG-3′, R 5′-TTGGTCTGGCTGAGTACCG-3′), EGR1 (5′-CCCAGTGGCTACCTCCTACC-3′, R 5′-CCTGGGAGAAAAGGTCGCTG-3′), and GAPDH (F 5′-GTCTCCTCTGACTTCAACAGCG-3′, R 5′-ACCACCCTGTTGCTGTAGCCAA-3′). GAPDH was used as a loading control. The Tra2β-relative mRNA expression was calculated using the 2−ΔΔCq method.
-
A total of 4 × 104 transfected Hela or Siha cells stably overexpressing Tra2β or transfected with siRNAs targeting Tra2β1 as required in 200-μL serum-free RPMI-1640 medium were seeded in the upper compartment of the Boyden chambers (Corning, NY, USA), and 700 μL of completed RPMI-1640 containing 10% FBS was added to the lower chamber. After incubation for 12 h at 37 °C, the cells migrating across the 8-μm polycarbonate were fixed, stained with 0.1% crystal violet, and counted.
-
Hela or Siha cells (4 × 104) transfected with Tra2β1 siRNAs or stably overexpressing Tra2β as required were re-suspended in 150 µL of RPMI-1640 medium and then seeded into the upper chamber with 100 μL of precoated Matrigel. Subsequently, 600 µL of completed RPMI-1640 medium containing 10% FBS was set in the lower chamber as a chemoattractant. After incubation for 12 h, the invading cells were fixed, stained with 0.1% crystal violet, and counted.
-
Female BALB/c nude mice aged 4–6 weeks and weighing 18–20 g were purchased from Jiangsu Jicui Yaokang Biotechnology Co., Ltd. (Nanjing, China), with five mice in each group, and all procedures were approved by the committee on the use of live animals in Jinan University (Ethical number: 20200807-01). Equal volumes of PBS and Matrigel were first used to suspend Hela cells at 5 × 107 cells/mL, and a 100-µL cell-containing mixture was subcutaneously injected into the abdomen of mice to establish tumor xenografts. Tumor size was measured with calipers every two days, and tumor volume was calculated using the following formula: V = (length × width2) / 2. At 28 days after injection, the mice were sacrificed, and the tumors were collected for Western blot and immunohistological analyses.
-
For the detection of Tra2β and Ki67 antigens, tissue sections were deparaffinized and microwaved in 10-mmol/L sodium citrate for 10 min and then incubated in H2O2 (0.3%) with methanol for 30 min. Nonspecific binding of antibodies was inhibited by incubating with normal goat serum (5%) for 30 min at room temperature. Then, tissue slides were incubated with rabbit anti-Tra2β polyclonal antibody (40 µg/mL) or mouse anti-Ki67 monoclonal antibody (5 µg/mL) for 18 h at 4 °C. After washing, slides were incubated in horseradish peroxidase-conjugated anti-rabbit IgG or anti-mouse IgG for 30 min, followed by chromogenic detection using DAB as the substrate.
-
Total RNA was extracted from Hela cells by using TRIzol® Reagent (Invitrogen) according to the manufacturer’s protocol. Then, the quality and integrity of total RNA were confirmed by agarose gel electrophoresis, and high-quality samples were sent to the Shanghai Biotechnology Co., Ltd. (Shanghai, China) for library construction and Illumina Genome Analyzer IIx sequencing. Libraries were prepared using the Illumina TruSeq RNA Library Prep Kit v2 (Illumina, San Diego, CA) according to the manufacturer’s protocol [14]. In brief, purified mRNAs were fragmented and converted to cDNA using reverse transcriptase. The resulting cDNAs were converted into double-stranded cDNAs and subjected to end-repair, A-tailing, and adapter ligation. The library was amplified by 8 PCR cycles. The library was quantified using a Qubit® 2.0 Fluorometer (Invitrogen) and validated using an Agilent 2100 bioanalyzer (Agilent, China). Clusters were constructed from the library using Illumina cBot diluted at a concentration of 10 pM and sequenced for 75 cycles on Illumina Genomic Analyzer IIx or 100 cycles on HiSeq-2000 (Illumina). High-quality reads that passed Illumina quality filters were retained for sequence analysis.
-
The input data of gene differential expression included ReadCount data obtained from gene expression level analysis. EdgeR was used for gene differential expression analysis of paired samples [15]. The absolute value of log2 (fold change) > 1 and the false discovery rate < 0.01 were used to identify the significance of different gene expressions. Subsequently, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses of the DEGs were performed using DAVID Tools (
https://david.ncifcrf.gov/ ). -
Confocal immunofluorescence was performed as previously described [13]. Hela cells transfected with pCMV-EGFP-Tra2β1 plasmids or empty vectors based on the experimental requirements were grown on polylysine-precoated slides. Cells were initially incubated with the rabbit-SP1 primary antibody at 4 °C overnight and then with Alexa Flour 592conjugated secondary antibody. Finally, DAPI nuclear counterstaining was performed. Slides were imaged using a 40 × NA 0.75 Oil Dic objective on an inverted laser-scanning confocal microscope (LSM700; Zeiss GmbH, Jena, Germany), and images were captured using Zeiss.
-
RNA IP was utilized to determine the interaction between SP1 mRNA and Tra2β. In brief, Hela cell lysates were incubated with anti-Tra2β for 4 h at 4 °C. Then, beads were added and incubated for another 2 h. The precipitated RNAs were eluted and analyzed by RT-qPCR.
-
The data were initially analyzed to assess for normality using the Shapiro-Wilk test and then presented as means ± SD. When data were only compared with the control, two-way ANOVA followed by Dunnett’s T3 post hoc test was used. Two-way ANOVA followed by Sidak’s post hoc comparison was used when data were compared among all groups. A two-sided P value of less than 0.05 was considered statistically significant. All data were analyzed using GraphPad Prism (La Jolla, USA). All experiments were repeated at least three times.
-
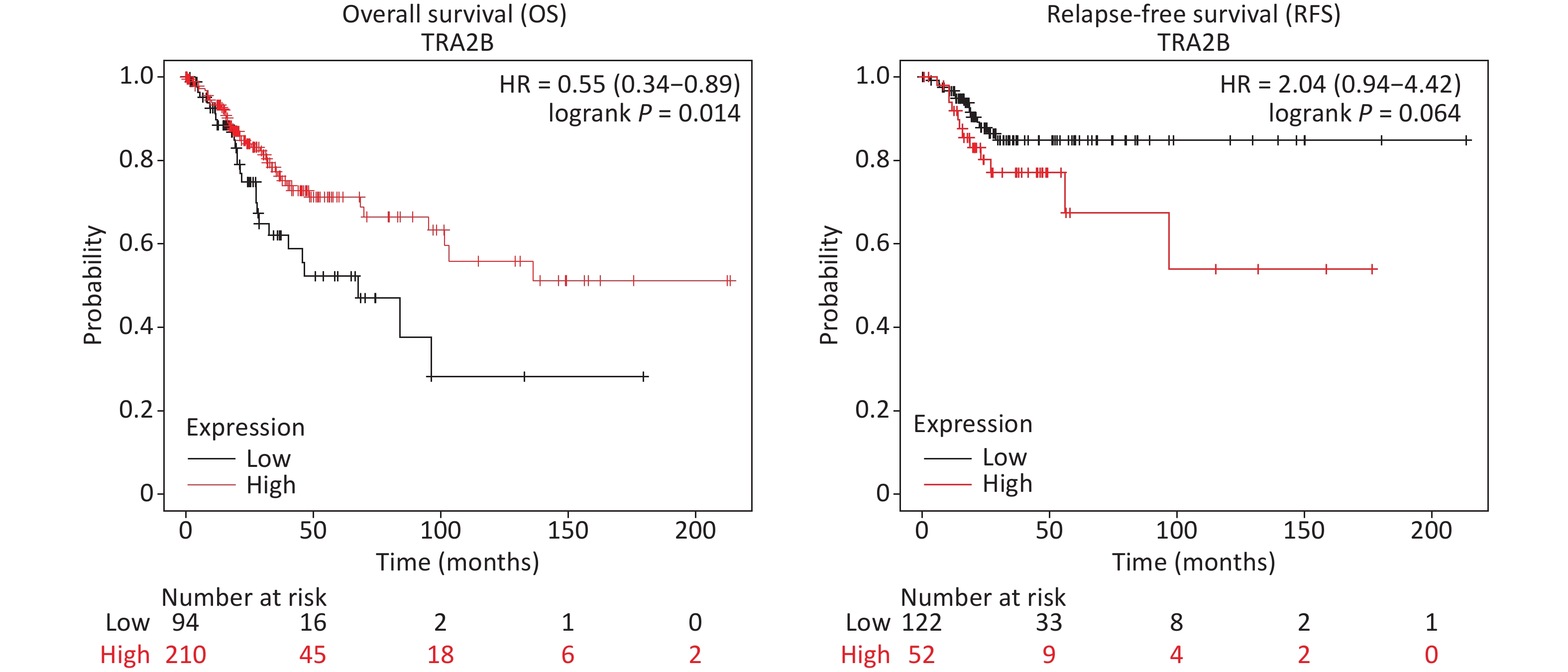
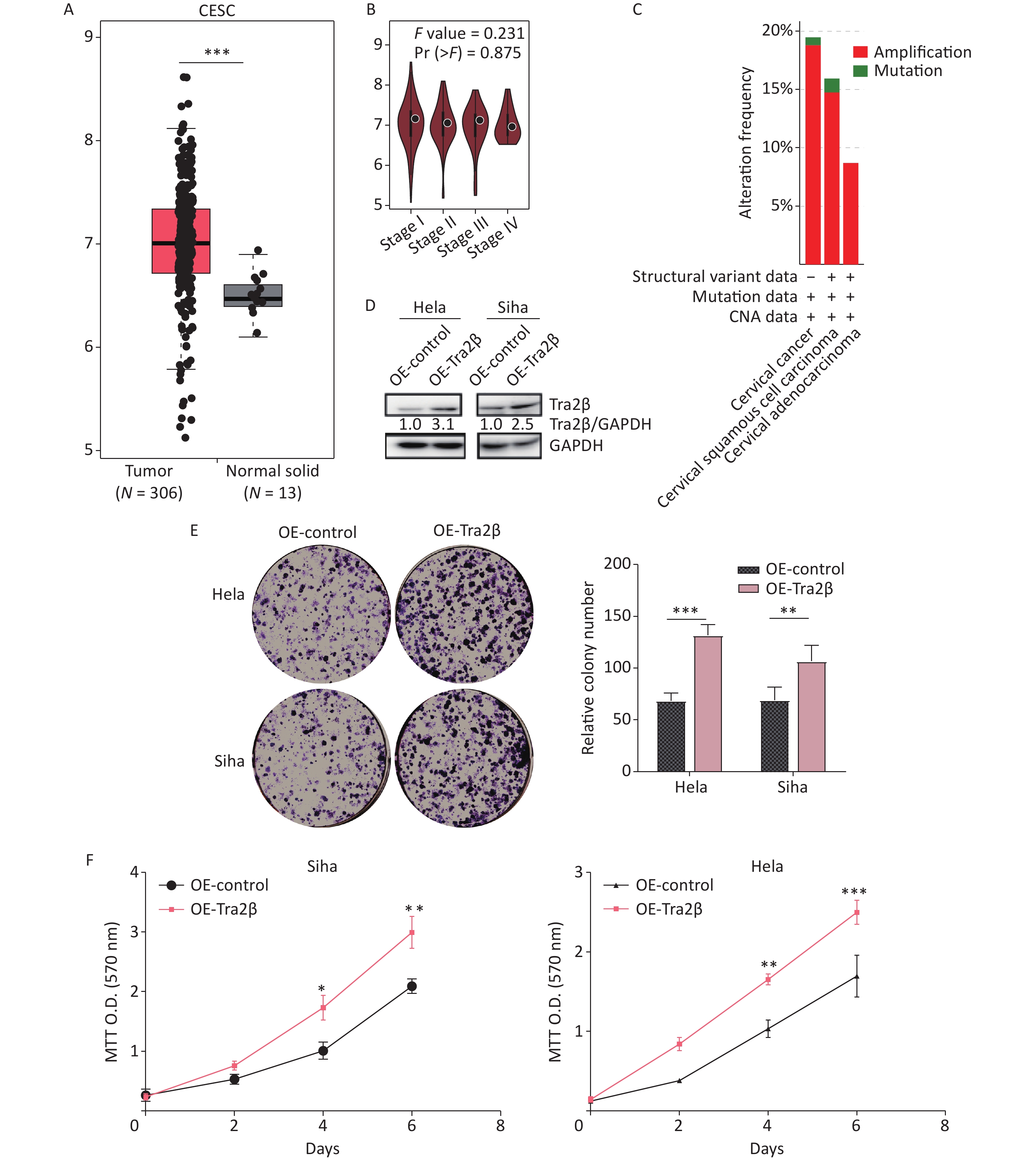
First, we investigated the transcriptional data of Tra2β in patients with cervical squamous cell carcinoma and endocervical adenocarcinoma (CESC) from GEPIA and cBioPortal databases. Data from GEPIA showed that the expression level of Tra2β was higher in cervical cancer tissues than in normal cervix tissues (Figure 1A). We also analyzed the expression of Tra2β with tumor stage for CESC. As shown in Figure 1B, no significant difference in Tra2β was observed among different tumor stages. The frequency of copy number amplification, deletion, and genetic mutation of Tra2β in patients with cervical cancer is shown in Figure 1C. About 17% of the clinical samples (102/605) had genetic amplification of Tra2β, and nearly 0.8% of the samples (5/605) had genomic mutations. However, Kaplan-Meier plotter showed that patients with high Tra2β mRNA expression had better overall survival (OS, P = 0.014), and no significant association was found between Tra2β expression levels and relapse-free survival (RFS) in patients with cervical squamous cell carcinoma (P = 0.064, Supplementary Figure S1 available in www.besjournal.com). In testing whether the molecular properties of Tra2β are relevant in an oncologic context, we performed colony formation assays on Hela and Siha cells with Tra2β overexpression or knockdown. We generated stable Tra2β overexpression in Hela and Siha cell lines using the recombinant lentiviral expression system. The exogenous Tra2β expression level in these cells was verified using Western blot (Figure 1D). Overexpressed Tra2β significantly promoted the cell clone formation in vitro (Figure 1E). In providing further evidence of Tra2β facilitating the cell growth of cervical cancer, MTT assays were performed on Hela and Siha cell lines with Tra2β overexpression. Consistent with the abovementioned results, Tra2β overexpression significantly increased the growth of Hela and Siha cells compared with control cells (Figure 1F).
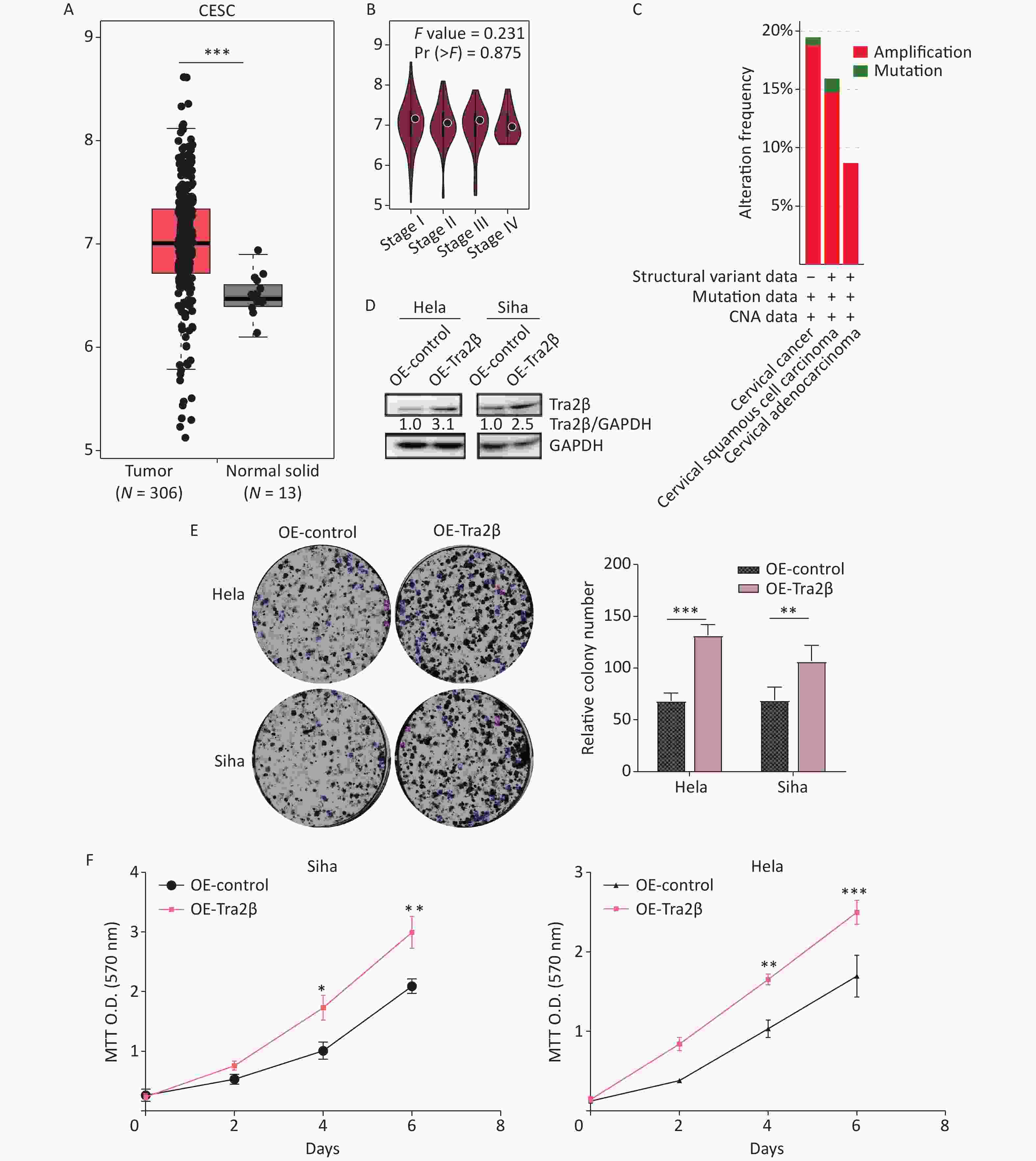
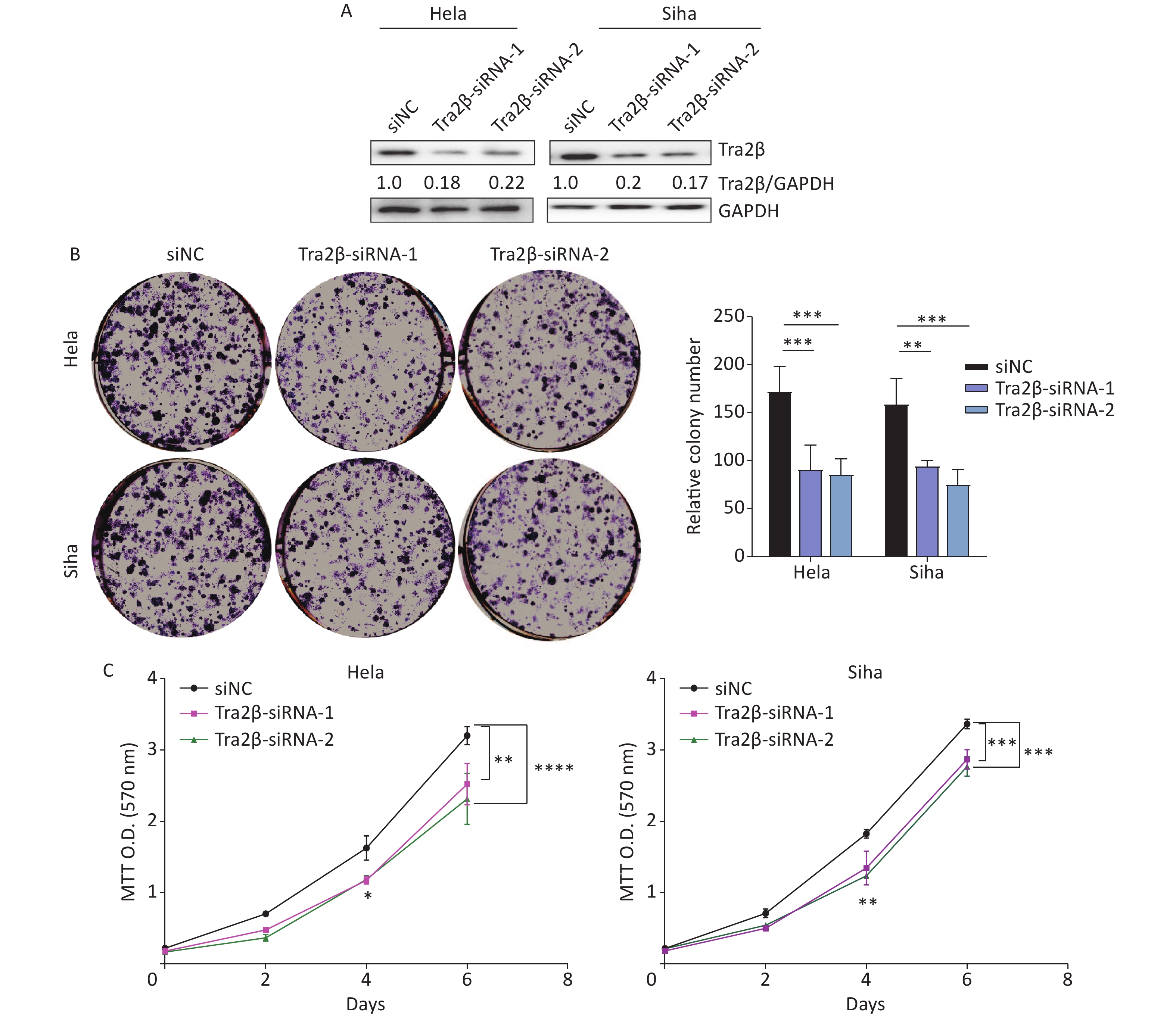
Figure 1. Overexpression of Tra2β promoted the proliferation of Hela and Siha cells in vitro. (A) The expression of Tra2β in patients with CESC (GEPIA,
http://gepia.cancer-pku.cn/ ). (B) Correlation between Tra2β expression and tumor stage in patients with CESC (GEPIA,http://gepia.cancer-pku.cn/ ). (C) Tra2β gene expression and mutation analysis in CESC (cBioPortal,http://www.cbioportal.org/ ). (D) The confirmation of stable Tra2β overexpression in Hela and Siha cells using Western blot assay. (E) Cell proliferation ability of Tra2β overexpression in Hela and Siha cells was assessed by colony formation assays. (F) Cell viability of cells of Tra2β overexpression in Hela and Siha cells was measured by MTT assays. Two-way ANOVA with Dunnett comparison (E and F) was used. The data shown are the mean ± SD of three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.Two siRNAs were designed and tested, and their respective cell lines were proven effective, reducing protein levels by more than 70% (Figure 2A). The downregulation of Tra2β significantly reduced clone formation in Hela and Siha cells (Figure 2B). Consistently, MTT colorimetric assay also showed that Tra2β silencing suppressed the proliferation of Hela and Siha cells compared with control cells (Figure 2C). Collectively, these results indicated that Tra2β promotes Hela and Siha cell growth in vitro.
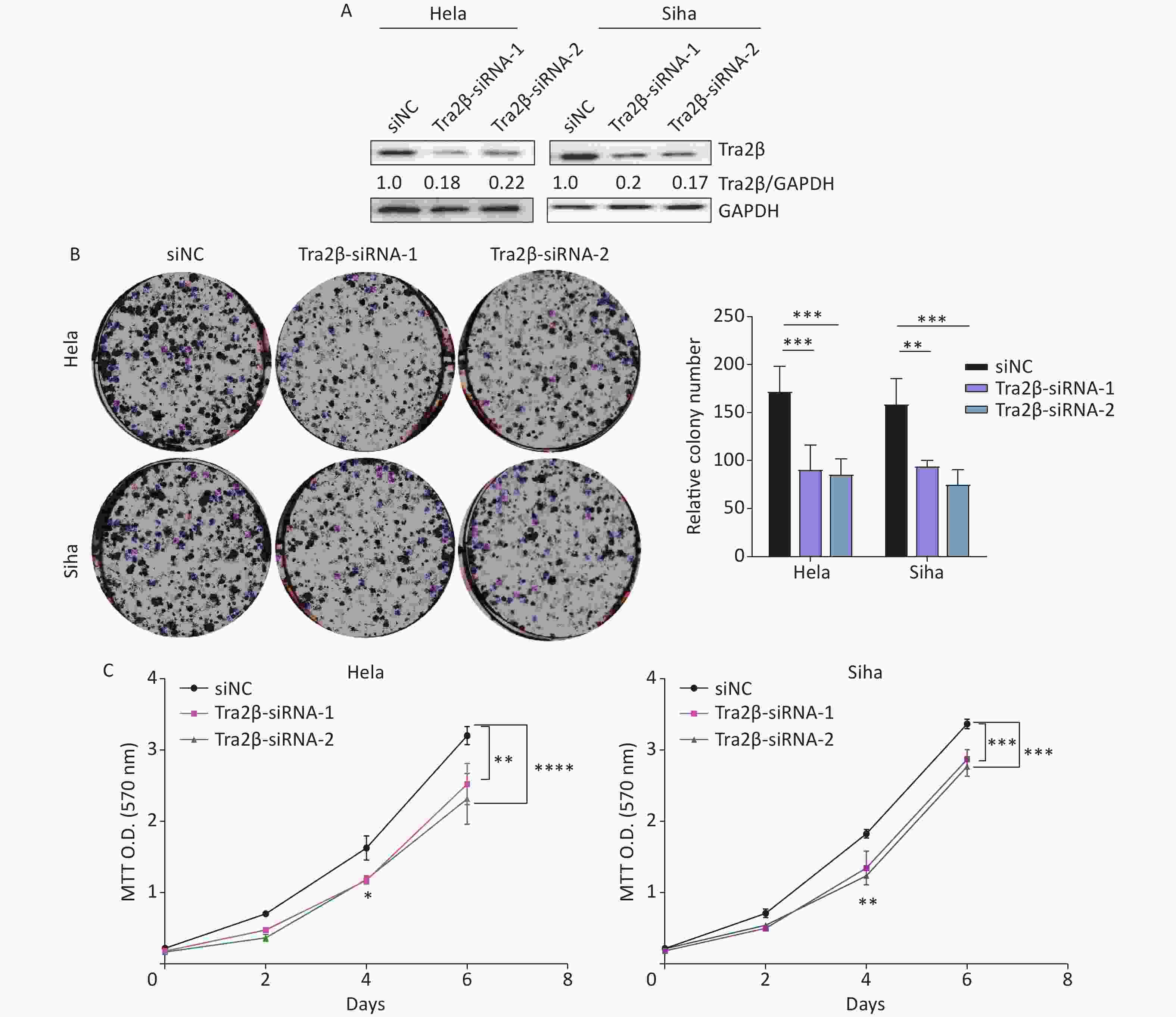
Figure 2. Knockdown of Tra2β suppressed the proliferation of Hela and Siha cells in vitro. siRNAs targeting Tra2β were transfected into Hela and Siha cells, followed by Western blot assay (A), colony formation assays (B), and MTT assays (C). Two-way ANOVA with Dunnett comparison (B and C) was used. The data shown are the mean ± SD of three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
-
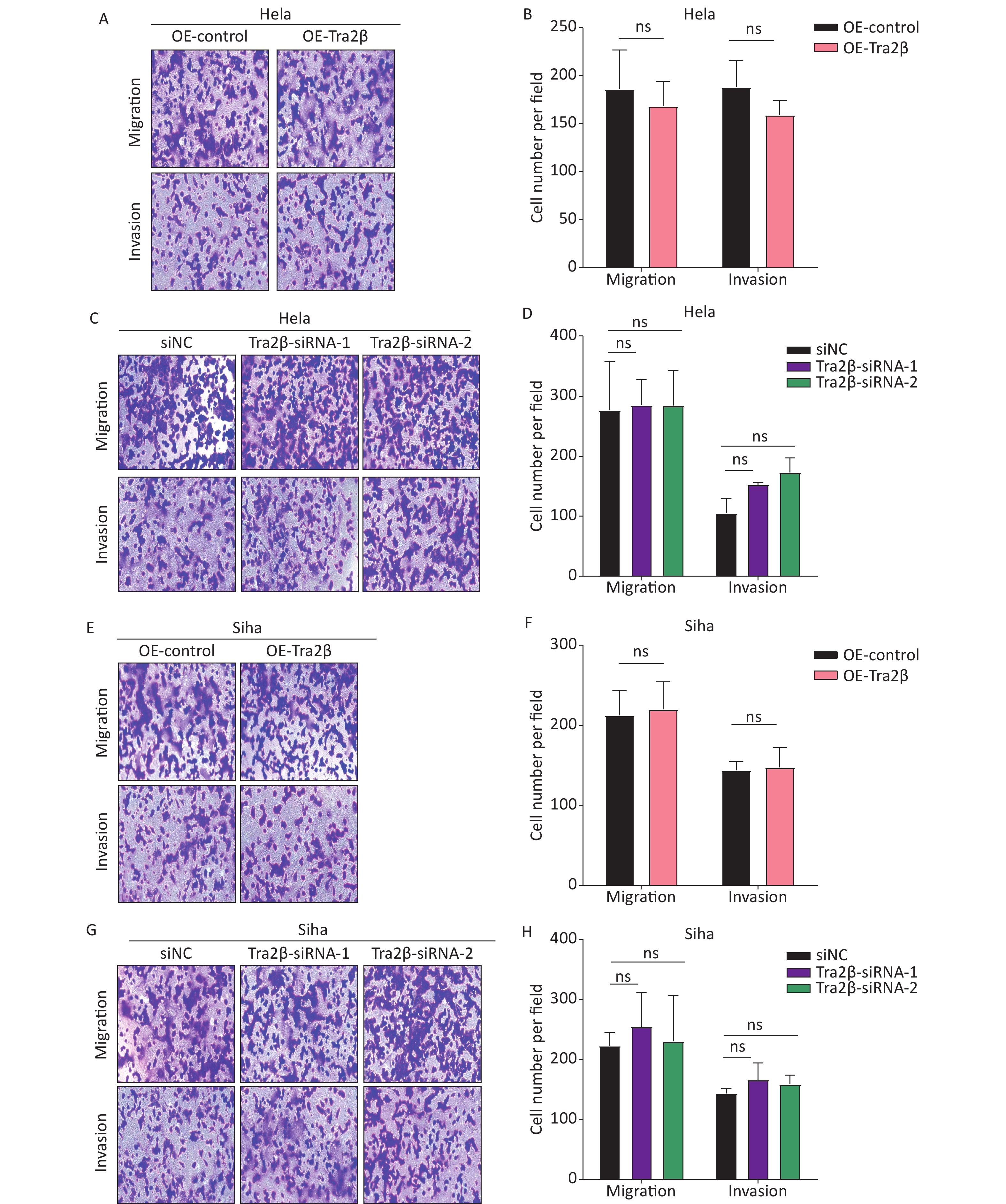
The migration and invasion of cancer cells are identified as key factors in cancer metastasis; thus, we further investigated the effects of Tra2β on the migration and invasion of Hela and Siha cells through in vitro Transwell migration and Matrigel invasion. Compared with control cells, the migratory and invasive abilities of Hela cells have not changed significantly either with Tra2β overexpression (Figure 3A and B) or knockdown (Figure 3C and D). Consistently, no significant changes in migration and invasion of Siha cells were found between the OE-Tra2β group and the control group (Figures 3E and F) or between the Tra2β knockdown group and the control group (Figure 3G and H). These results indicated that the alteration of Tra2β expression does not affect the migration and invasion of cervical cancer in vitro.
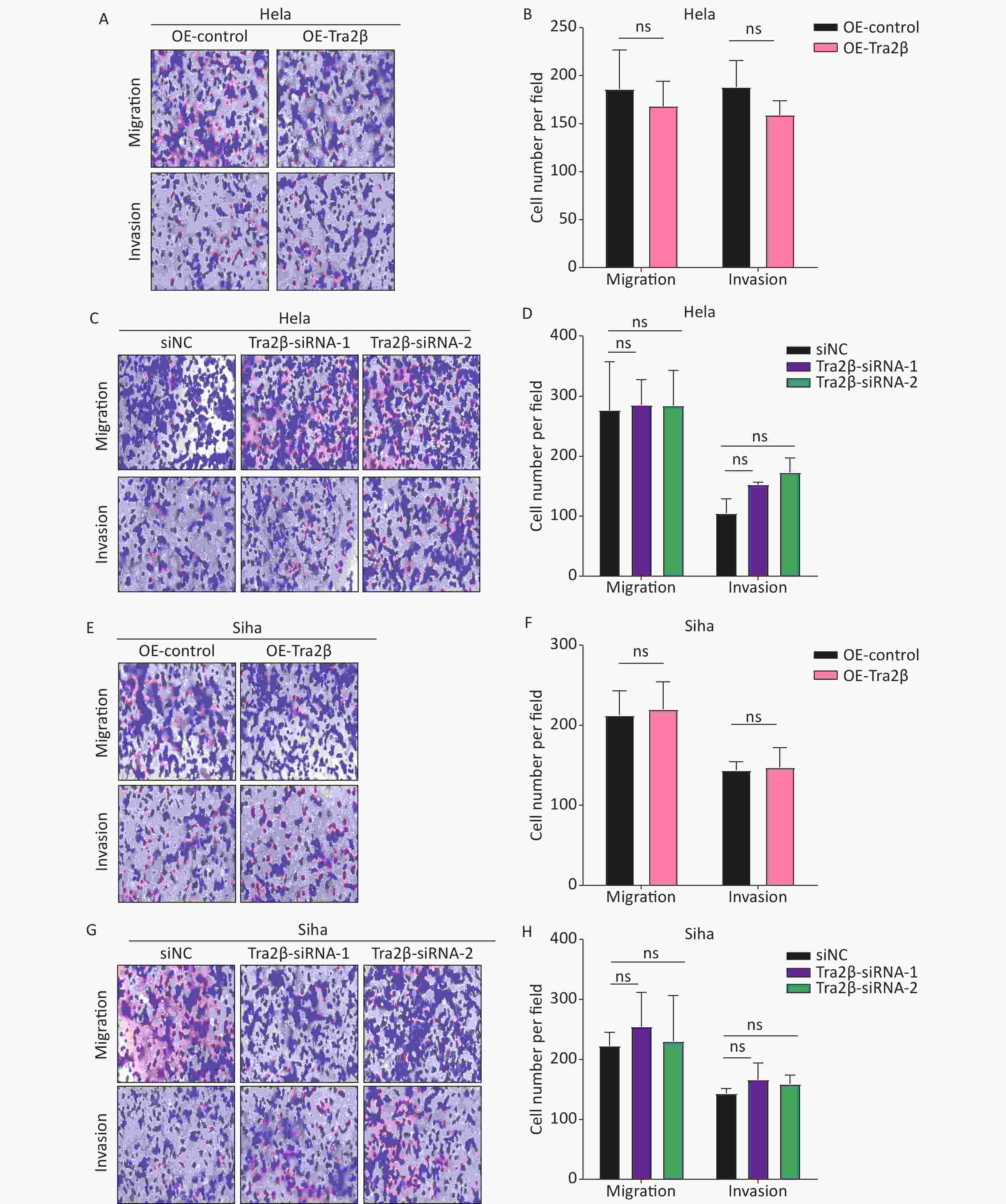
Figure 3. Tra2β had no effect on the migration and invasion of Hela and Siha cells in vitro. (A–D) Transwell assays were conducted to test the migratory (top) and invasive (bottom) ability of Hela cells with stable Tra2β overexpression (A and B) or Tra2β knockdown (C and D). (E–H) Transwell assays were conducted to test the migratory (top) and invasive (bottom) ability of Siha cells with stable Tra2β overexpression (E and F) or Tra2β knockdown (G and H). Five random fields of each well were counted under a microscope with a magnification of ×100 and followed by statistical analysis (B, D, F, and H). Two-way ANOVA with Dunnett comparison (B, D, F, and H) was used. Data shown are the mean ± SD of three independent experiments; ns indicates no significant difference.
-
Tumorigenicity experiments were performed by subcutaneous injection of 5 × 106 Hela cells into the abdomen of female nude mice to confirm the role of Tra2β in driving cervical cancer growth. Twenty-eight days after injection, tumor size in the Tra2β-overexpressed group was generally larger than that in the control group (Figure 4A), and the average weight of the tumor in the Tra2β-overexpressed group was evidently heavier than that in the control group (Figure 4B). In addition, tumor growth was significantly accelerated in the Tra2β-overexpressed group (Figure 4C). Moreover, Tra2β overexpression increased the percentage of Ki-67-positive cells in the tumor xenografts of nude mice compared with the control group (Figure 4D). Moreover, the IHC assay showed that Tra2β was preferentially expressed in the nucleus, which was consistent with previous studies [12,16]. Collectively, these observations indicated that Tra2β plays a role in regulating cervical cancer formation and growth.
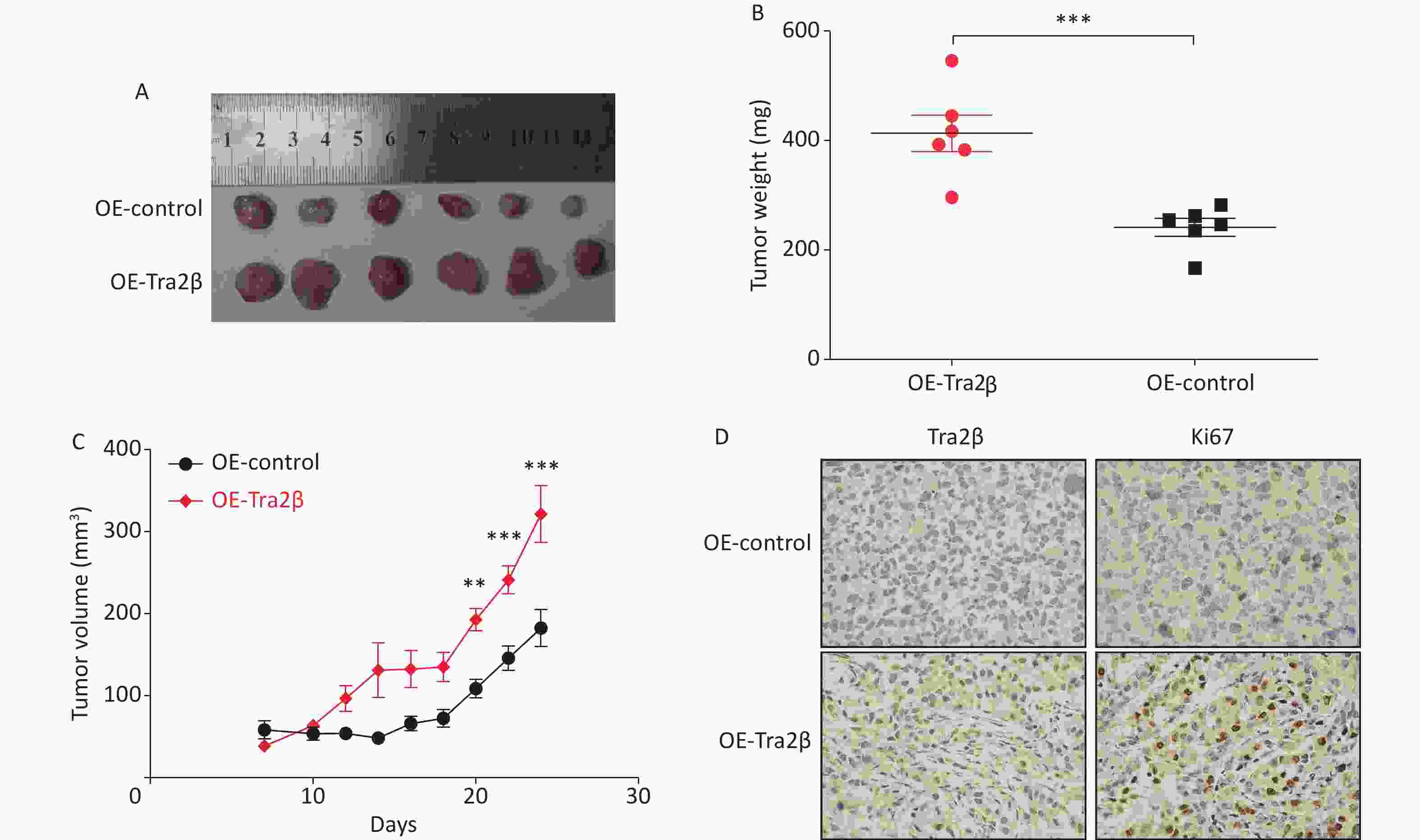
Figure 4. Tra2β promoted the tumor growth of cervical cancer in vivo. (A) Formation of tumors in nude mice injected with Tra2β-overexpressed or control Hela cells. (B) Measurement of subcutaneous tumor weight. (C) Tumor growth curves of nude mice during the experiment. (D) Tra2β and Ki67 proliferation index analysis in tumors by immunohistochemistry. Magnification of (D): ×400. Two-way ANOVA with Dunnett comparison (C) and unpaired t-test (B) were used, **P < 0.01, ***P < 0.001.
-
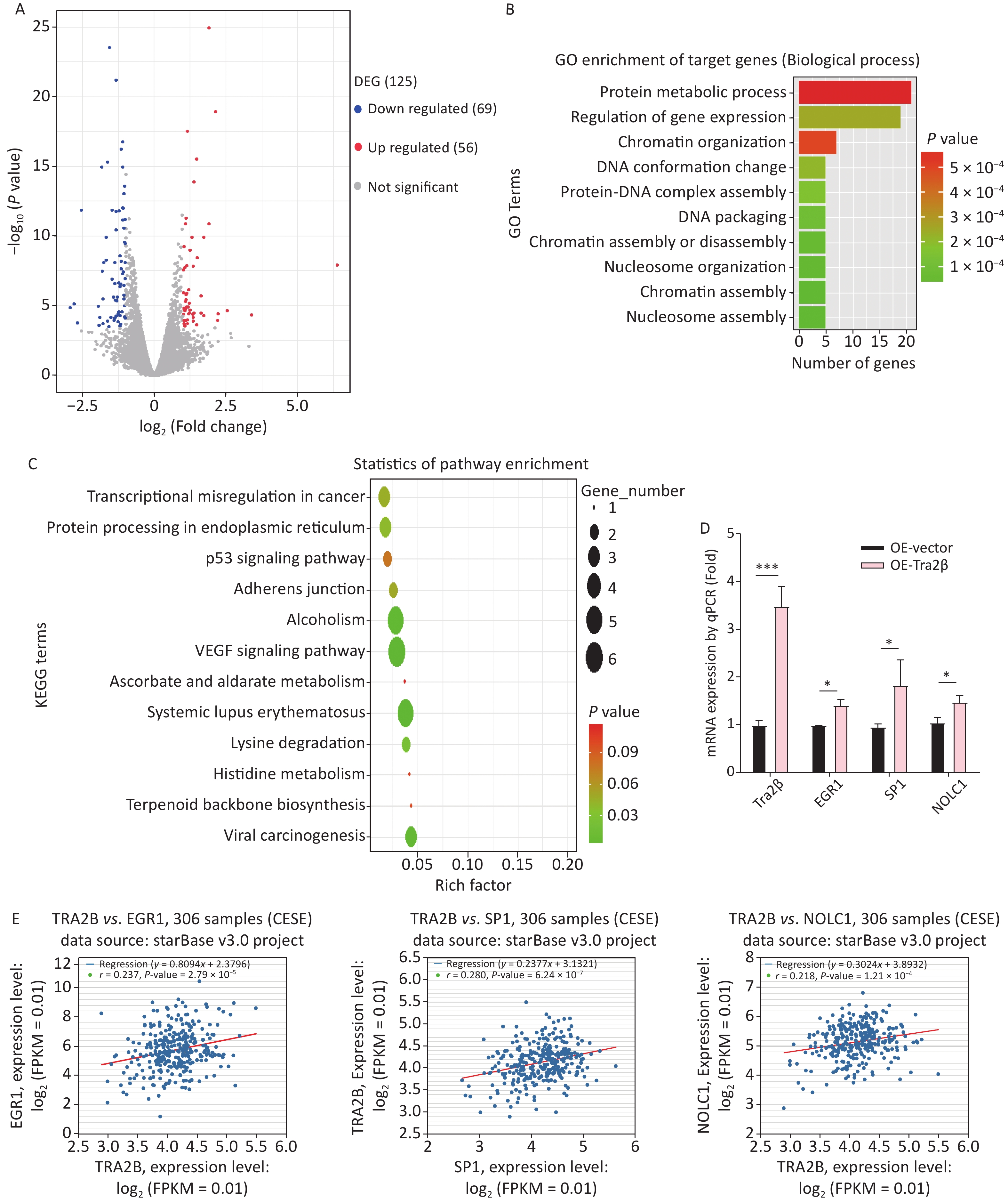
The major RNA-binding site for Tra2β is GAA, particularly the AGAA-rich sequence, which lies within multiple mRNAs [17]. The transcriptomic profiles of Hela cells with stable Tra2β overexpression and control cells were compared using the next-generation RNA-Seq to uncover Tra2β-regulated potential targets controlling cell growth. A total of 125 DEGs were identified, among which 56 genes were upregulated, and 69 genes were downregulated (Figure 5A and Supplementary Table S1 available in www.besjournal.com). GO (Figure 5B) and KEGG pathway (Figure 5C) analyses identified significantly enriched pathways in DEGs, including the regulation of gene expression, protein–DNA complex assembly, chromatin assembly, protein metabolic process, transcriptional misregulation in cancer, VEGF signaling pathway, adherence junction, and p53 signaling pathway. RT-qPCR was used to confirm the RNA-Seq data. Of the top 10 DEGs, three (EGR1, SP1, and NOLC1) were randomly selected. As presented in Figure 5D, all these genes were highly expressed in Tra2β-overexpressed Hela cells, as compared with control cells, which is with the RNA-Seq data. Furthermore, the correlation of EGR1, SP1, or NOLC1 with Tra2β in CESC tissues was analyzed by using the ENCORI website. As shown in Figure 5E, EGR1, SP1, and NOLC1 were positively correlated with Tra2β expression in 306 CESC tissue samples. Overall, Tra2β plays an important biological role in cervical cancer–like transcriptional dysregulation.
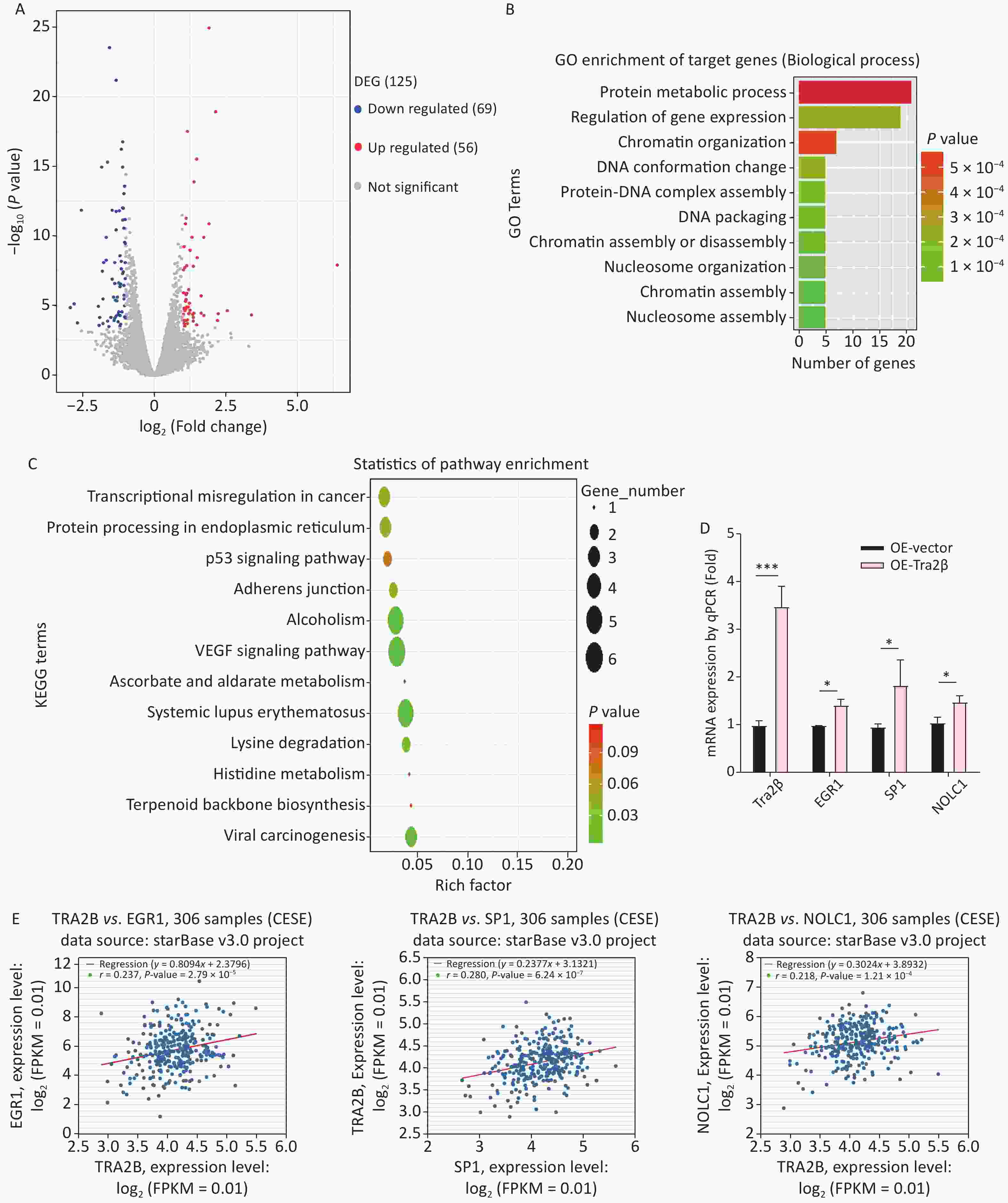
Figure 5. Tra2β could regulate EGR1, SP1, and NOLC1 expression. (A) Volcano plot of global DEGs between Hela cells with stable Tra2β-overexpressed and control cells. (B–C) The function of DEGs was predicted by analysis using GO (B) and KEGG (C) by DAVID Tools (
https://david.ncifcrf.gov/ ). (D) Relative mRNA levels of Tra2β, EGR1, SP1, and NOLC1 in Hela cells with stable Tra2β-overexpressed or control cells were used to perform RT-qPCR experiments. The mRNA level of GAPDH was set as the internal control. (E) The correlation among EGR1, SP1, NOLC1, and Tra2β in 306 patients with CESC was analyzed by using the ENCORI database (http://starbase.sysu.edu.cn/index.php ). Two-way ANOVA with Dunnett comparison (D) was used, *P < 0.05, ***P< 0.001. 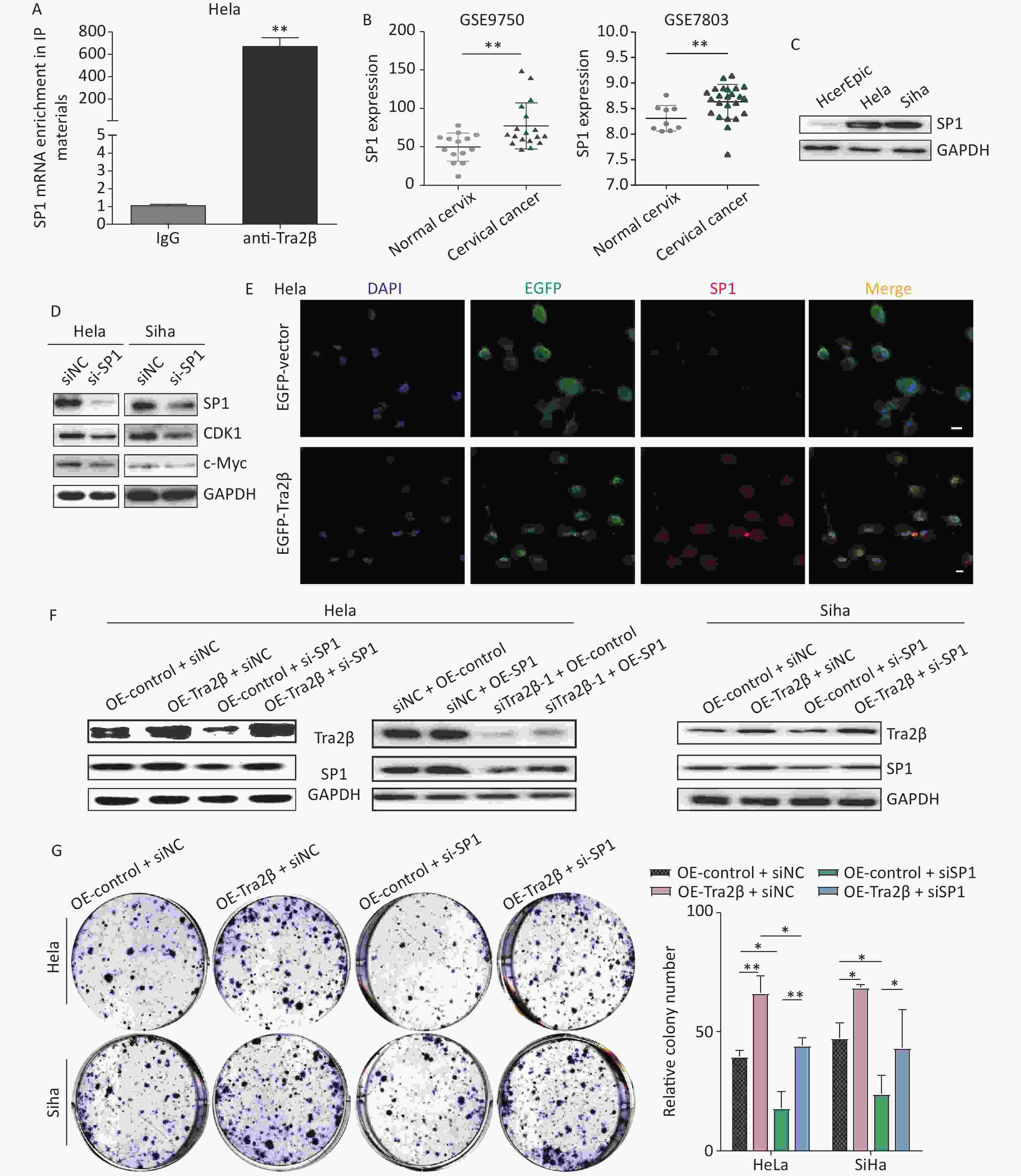
-
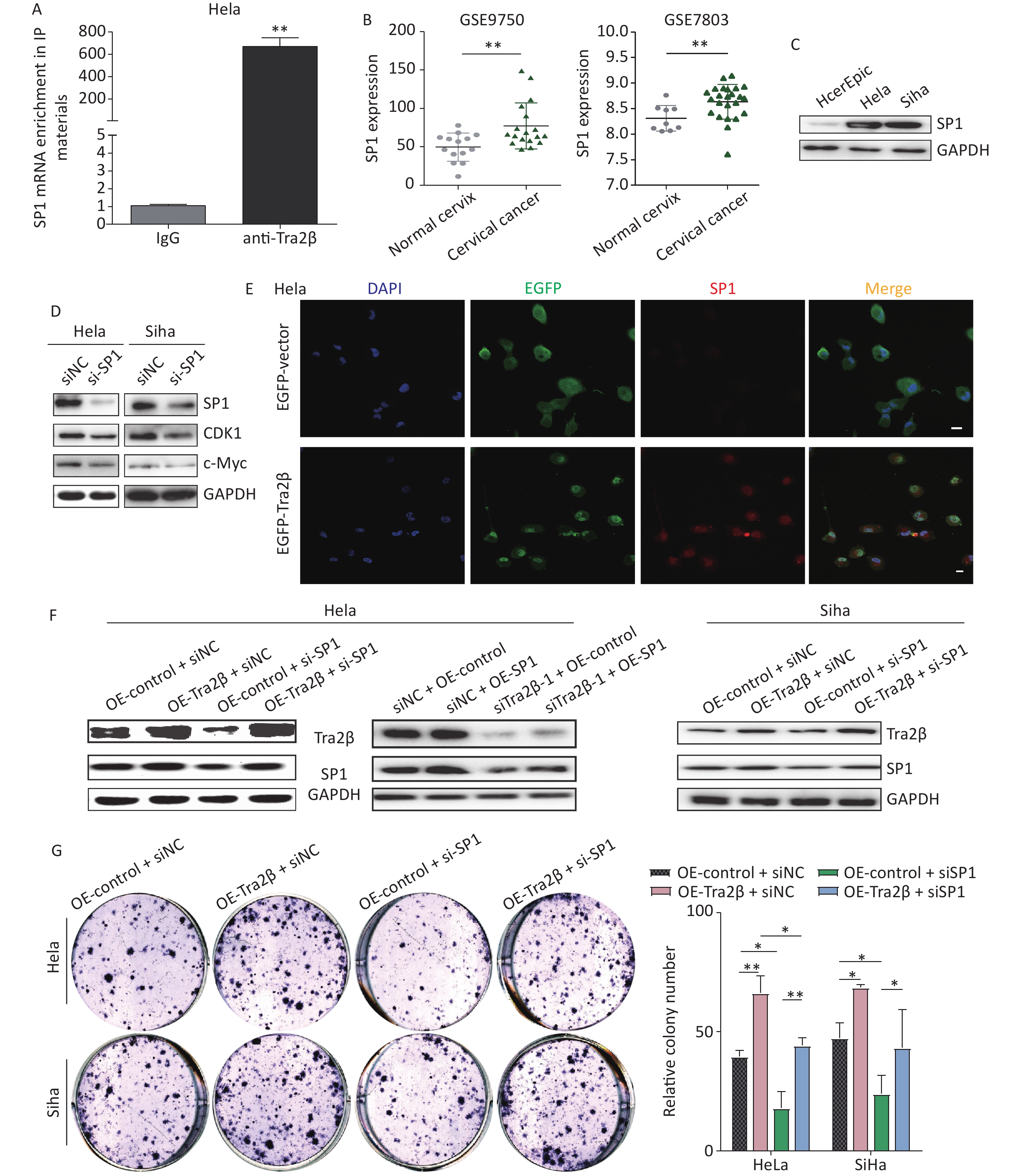
Based on previous reports, SP1 is identified as a transcription factor by binding to promoters of target genes and is believed to play important roles in the regulation of various key biological functions, such as cell proliferation, clonogenicity, cycle progression, and apoptosis [18]. First, the RNA-IP assay confirmed that Tra2β proteins could bind with SP1 mRNA (Figure 6A). In exploring the significance of SP1 in the development of cervical cancer, we first validated the SP1 expression of cervical cancer in the Gene Expression Omnibus database, including GSE7803 and GSE9750. As shown in Figure 6B, SP1 mRNA expression was higher in cervical cancer tissues than in normal cervical tissues. Then, the protein level of SP1 was examined in normal cervical epithelial cells (HcerEpic) and cervical cancer cells (Hela and Siha). Western blot showed that the level of SP1 in the two cervical cancer cell lines was evidently higher than the normal cervical epithelial cells (Figure 6C). SP1-targeted siRNAs downregulated the expression of SP1 and its major downstream oncogenes (c-Myc and CDK1) [19], which was consistent with previous findings, showing that high SP1 expression is correlated with the malignant progression of cervical cancer (Figure 6D). The expression of Tra2β and SP1 in tumor xenografts of nude mice formed by Hela cells was significantly higher than those in adjacent normal tissues (Supplementary Figure S2A available in www.besjournal.com). In obtaining evidence regarding the Tra2β/SP1 axis in cultured Hela cells, we performed indirect immunofluorescence staining of Tra2β and SP1 followed by confocal scanning. The cells were transfected with pCMV-EGFP-Tra2β1 or pCMV-EGFP empty vector for 24 h and then subjected to fluorescence staining as described in “Materials and Methods”. Figure 6E shows that in cells transfected with pCMV-EGFP empty vector, the EGFP protein was evenly distributed throughout the cell, with weak SP1 staining intensity. On the contrary, in cells transfected with pCMV-EGFP-Tra2β1-expressing plasmids, the EGFP-Tra2β protein was primarily concentrated in the nucleus, accompanied by high-intensity immunoreactive SP1 in the nucleus, which was conducive to the biological activity of transcription factor SP1. Functional rescue assays were performed by using Western blot, colony formation, and MTT to clarify whether SP1 contributes to the promotion effects of cervical cancer cell growth by Tra2β. As shown in Figure 6F, SP1 siRNAs effectively diminished SP1 expression in Siha cells and, to a lesser extent, in Hela cells. Tra2β overexpression (OE-Tra2β + si-NC group) dramatically upregulated SP1 expression when compared with the OE-control + si-NC group. On the contrary, the overexpression of SP1 (si-Tra2β + OE-SP1) alleviated SP1 deficiency compared with the si-Tra2β + OE-control group. Therefore, SP1 knockdown hindered the proliferation of Hela and Siha cells by Tra2β overexpression (Figure 6G and H). Collectively, these findings indicated that the Tra2β/SP1 axis promotes cervical cancer cell growth.
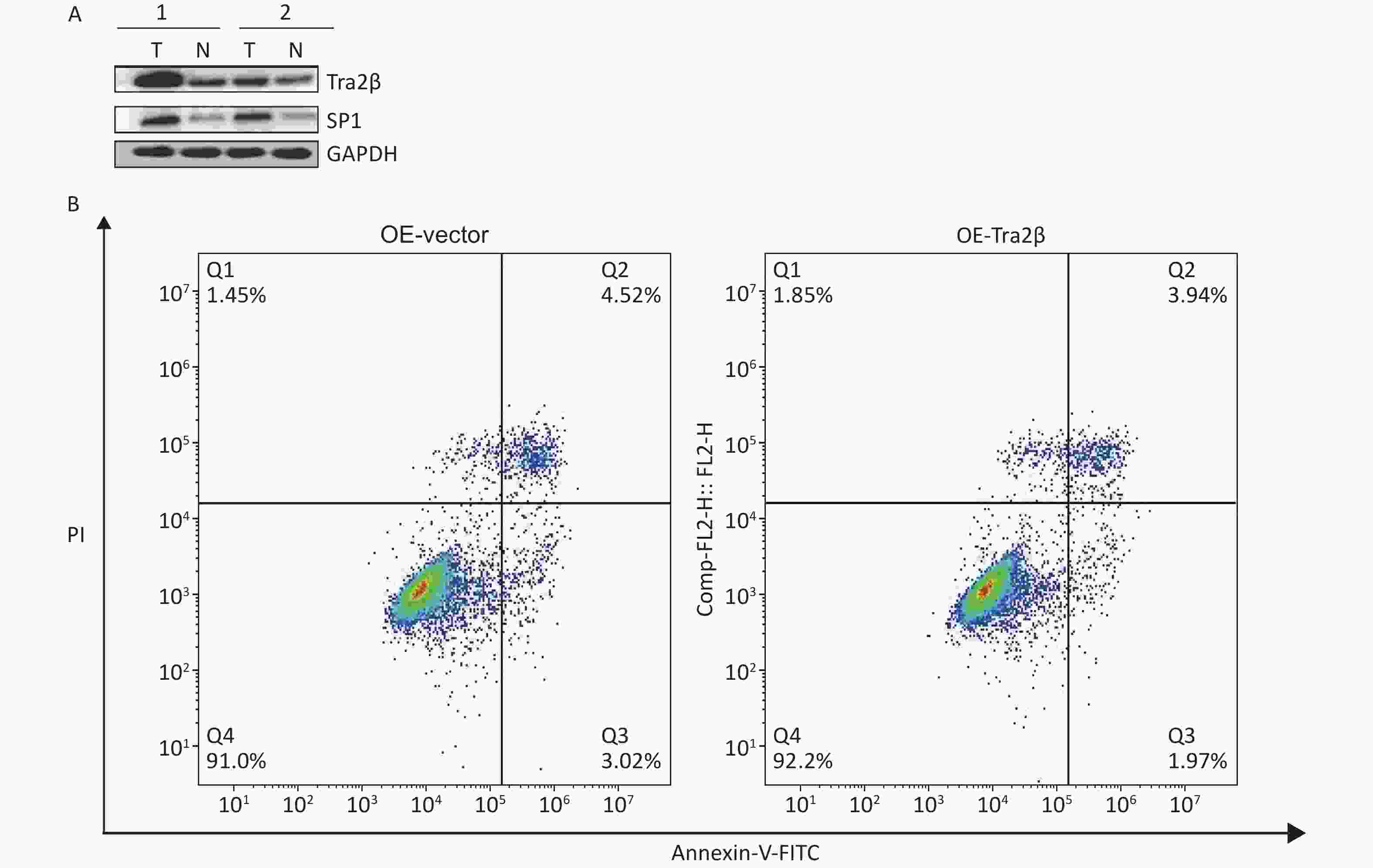
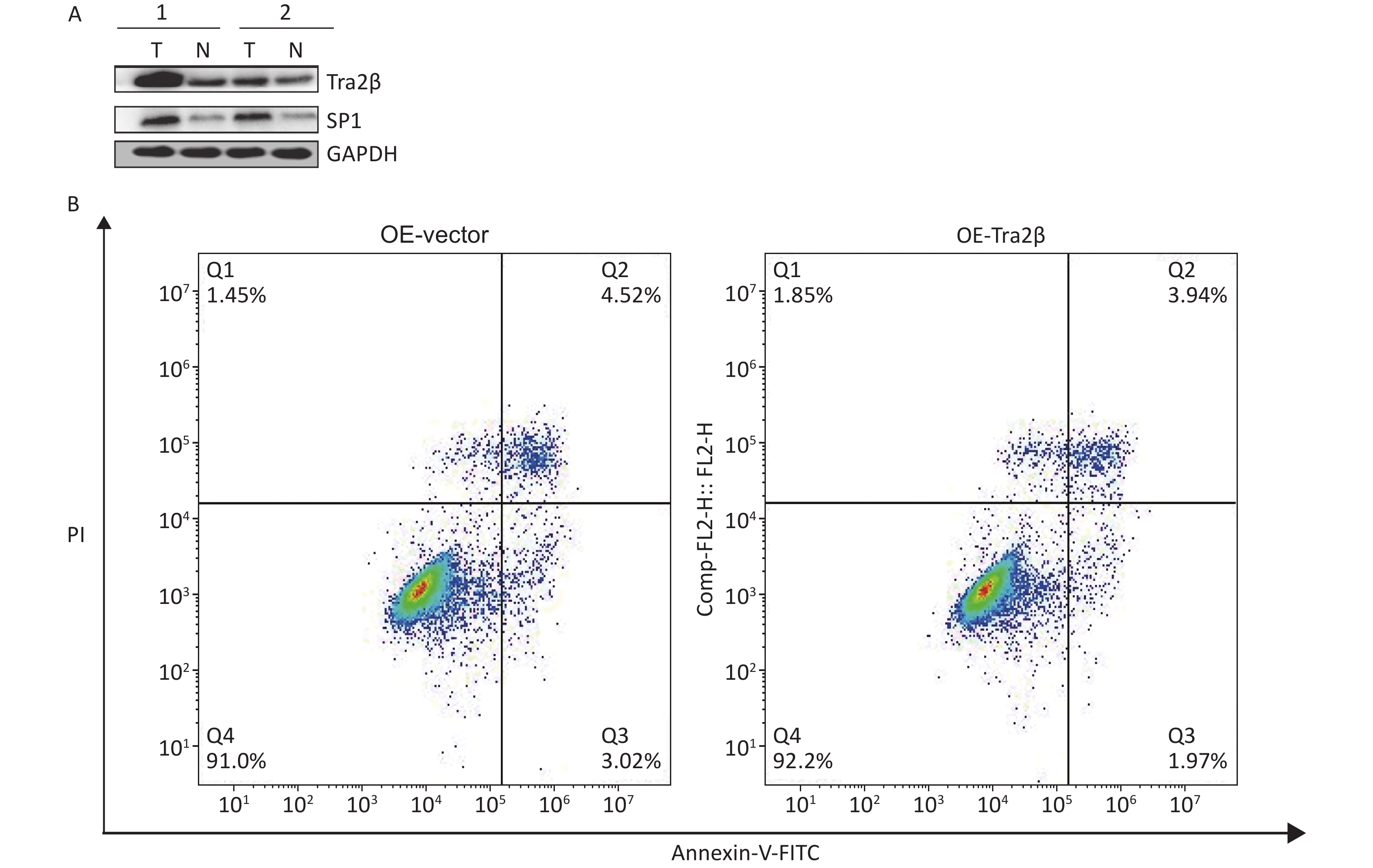
Figure S2. (A) Expressions of Tra2β and SP1 were analyzed in two representative tumor tissues in nude mice formed by Hela cells. T: tumor tissues; N: adjacent normal tissues. (B) Annexin-V FITC/PI assay was used to detect apoptotic cells in Tra2β-overexpressed or control Hela cells.
-
The occurrence and development of cervical cancer is a broad pathological process, which involves not only the changes of proto-oncogenes E6 and E7 that promote the degradation of tumor suppressors p53 and pRb [20], but also the intricate changes in the expression of various regulatory proteins that play a key role in cell proliferation, differentiation, drug resistance, apoptosis, migration, and invasion. Despite the development of treatments for cervical cancer, such as surgical resection, chemotherapy, and radiotherapy, treatment failure and mortality in patients with cervical cancer remain high. Some recent reports have revealed that the abnormal expression of splicing factors in cancer cells might have important therapeutic significance [21]. Tra2β protein expression is important for cancer cell biology, and whether it exerts pro-tumor or anti-tumor effects depends on the important proteins and tumor type that it regulates [16,22]. Watermann et al. reported that the significant induction of Tra2β at mRNA and protein levels is associated with invasive breast cancer [23]. Kuwano et al. reported that Tra2β inhibits the apoptosis of human colon cancer cells by upregulating the expression of Bcl-2 through its competition with miR-204 and the sensitivity of human lung adenocarcinoma cells (A549) to anticancer drugs [24]. The downregulation of Tra2β modified alternative splicing of CD44 pre-mRNAs and inhibited cell growth from regulating cellular oxidative response in a gastric cancer cell line [25]. Data from GEPIA showed that Tra2β expression was higher in cervical carcinoma tissues than that in normal cervical tissues, and the expression of Tra2β was not associated with tumor stages (Figure 1A–B). However, patients with high Tra2β mRNA expression had better OS (P = 0.014, Supplementary Figure S1). The abovementioned findings and bioinformatic analysis of the clinical significance of Tra2β in cervical cancer led us to wonder whether Tra2β played oncogenic or tumor-suppressive roles in cervical cancer progression. Combining MTT, colony formation, Transwell assays, and nude mouse tumor formation experiments, we clearly demonstrated that Tra2β plays an important role in the growth of cervical cancer cells (Figures 1, 2, and 4) and that Tra2β had no effect on cell migration, invasion, and apoptosis (Figure 3, Supplementary Figure S2B). The promotive effects of Tra2β on cell viability and proliferation might be mediated by the upregulation of transcription factor SP1 (Figures 5 and 6).
Previous studies have demonstrated the function of SP1 as a transcription factor, contributing to cell proliferation and colony-forming capacity [26]. Deng et al. showed that high SP1 expression was positively correlated with advanced International Federation of Gynecology and Obstetrics stage, lymph node metastasis, and lymphovascular space invasion of cervical cancer [19]. Nearly all cases of cervical cancer were due to HPV infection, particularly HPV16 and HPV18. The oncogenic potential of high-risk HPV types 16 and 18 depends on the expression of two viral oncogenes, E6 and E7. The HPV-18 upstream regulatory region contains SP1-binding sites, which have been shown to control the expression of E6 and E7 genes [27]. The downstream target genes of SP1 include well-known cyclin-dependent kinase inhibitors p27 and p21; CDK1, which is essential for G1/S and G2/M phase transitions of the eukaryotic cell cycle [19]; and oncogenes Bmi1, c-Myc, KLF4, and ABCG2, which were related to the acquisition of cancer stem cell phenotypes. Hence, we first hypothesized and focused on the possibility that Tra2β might regulate the proliferation of cervical cancer cells by regulating the expression of SP1 and strived to verify this hypothesis. As shown in Figure 6, we have established a functional link between Tra2β and SP1 and demonstrated that the function of Tra2β in the regulation of the proliferative ability of Hela and Siha cells is dependent on SP1 expression. The knockdown of SP1 accelerated the overexpression of SP1 and inhibited G2/M phase arrest in cervical cancer cells [19], indicating that Tra2β might play an important regulatory role in radioresistance and drug resistance in patients with cervical cancer who receive radiotherapy or chemotherapy. Metformin, an anti-diabetes drug, has been shown to induce the downregulation of SP1 and significantly reduce the mortality rate in elderly diabetic women diagnosed with cervical cancer [28]. If we can disrupt the stimulation of SP1 by Tra2β in cervical cancer, then the therapeutic effect of metformin is improved in elderly patients with cervical cancer and diabetes. Furthermore, the mechanism through which Tra2β regulates SP1 transcriptional levels must be investigated, whether by regulating alternative splicing, stabilizing mRNA, or enhancing transcription.
Considering that the G/A-rich enhancer elements are often found biologically important in alternatively spliced tumor genes, a total of 125 DEGs induced by Tra2β overexpression were screened in our study, and GO and KEGG analyses of DEGs revealed changes in biological functions and metabolic pathways, including the regulation of gene expression, transcriptional misregulation in cancer, VEGF signaling pathway, and p53 signaling pathway, which were related to the occurrence and development of malignant tumors [29, 30]. Therefore, the changes in biological behavior in cervical cancer progression promoted by Tra2β may be broader phenotypic changes rather than a single regulatory effect.
Among the prognostic subgroups of cervical squamous cell carcinoma, patients with high Tra2β expression had better OS, and no significant association was observed between Tra2β expression levels and RFS (Supplementary Figure S1). First, Tra2β has no effect on the migration and invasion ability of cervical cancer cells (Figure 3), and tumor metastasis was the main cause of death in patients with cervical cancer, which may explain the small effect of Tra2β on prognosis. Second, our phenotypic results were obtained from immortalized cervical cancer cell lines and nude mice xenograft tumor models, excluding the role of tumor immunity. Third, the prognosis of cervical cancer is related to perioperative factors, including anesthesia and anesthesia technology. Zhang et al. reported that sevoflurane enhances the proliferation and metastatic potential of cervical cancer cells by upregulating histone deacetylase 6, thereby increasing recurrence risk [31]. Another possible factor is the distinct cellular sublocalization of Tra2β in patients with cervical cancer. Li et al. demonstrated that the nuclear and cytoplasmic localizations of the Tra2β protein were correlated with its phosphorylation status. When Tra2β proteins are localized in the nucleus, active transcription and splicing occur. However, when Tra2β proteins shuttle to the cytoplasm, translational regulation, mRNA stability, and mRNA localization are their primary functions [32]. Liu et al. pointed out the potential role of Tra2β in mRNA trafficking, leading to the localized expression of the protein, which could influence cancer development [33]. Boris et al. reported that only high nuclear expression levels of splicing factor Tra2β have a direct regulatory influence on alternative mRNA processing and pronounced effects on the cellular function and tumor biology of the affected tumors, leading to poor patient outcomes [12]. These factors were cross-talked, which led to phenotypic changes. Our study has some limitations. For example, only two immortalized cervical cancer cell lines were used in the current study. Consequently, the biology of these cells is not as newly isolated as that of primary tumor samples from patients. The present study was focused on the molecular mechanism of Tra2β-induced cervical cancer cell malignancy in cell culture experiments and nude mouse models, whereas whether potential phenotype changes may occur during clinical trials and the mechanism through which Tra2β affects disease progression in patients with cervical cancer remain unknown and deserve further investigations.
Our study demonstrated the function of the Tra2β/SP1 axis in cervical cancer in vitro and in vivo, which provides a comprehensive understanding of the pathogenesis and improves the treatment strategy of cervical cancer.
-
None.
-
LI Mo Juan: Study design and execution, data analysis, writing-original draft, writing-review, and editing. XIONG Dan: Formal analysis, writing-original draft, Writing-review and editing. WANG Shuai: Data curation, software, formal analysis. HUANG Hao: Funding acquisition, conceptualization, supervision, writing-review, and editing.
-
The manuscript is approved by all authors for publication.
-
Datasets used and/or analyzed in this study can be obtained from the corresponding author upon reasonable request.
-
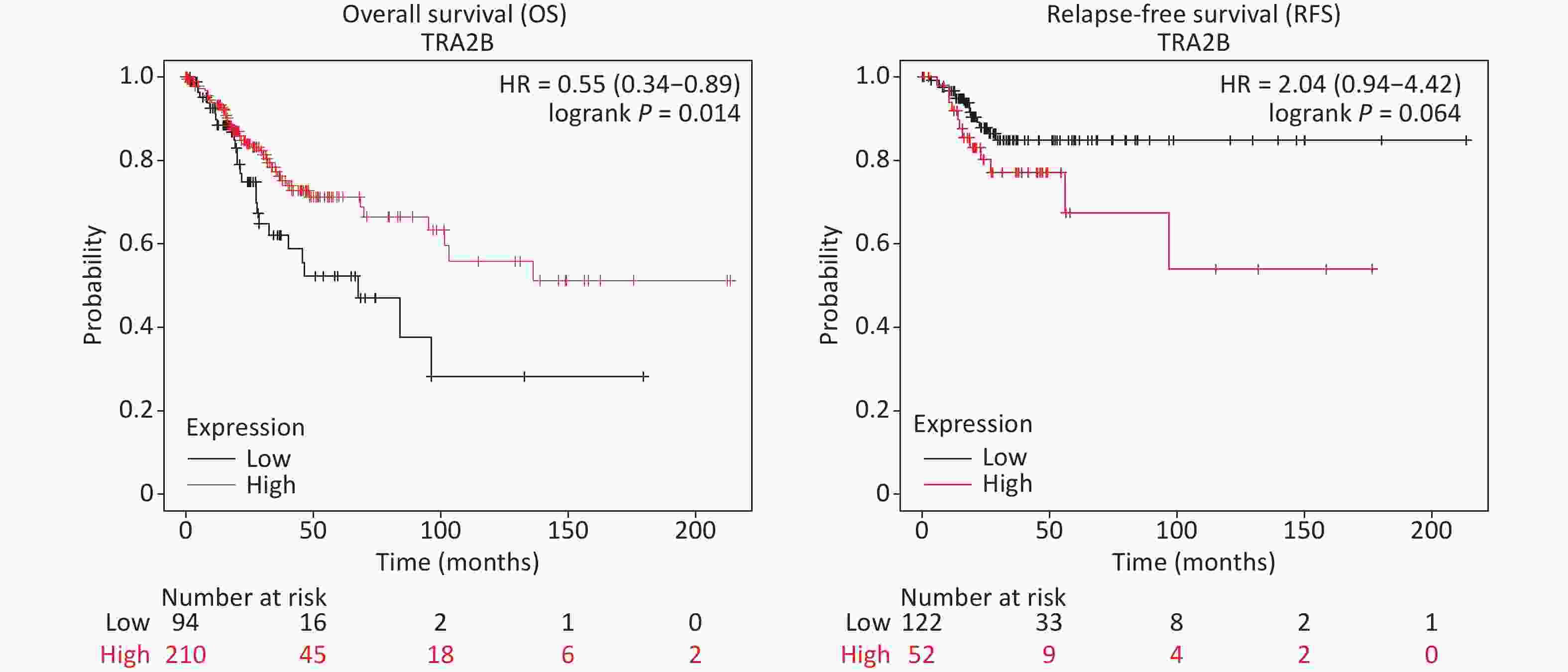
Figure S1. Survival differences of patients with CESC with high and low Tra2β gene expression.
doi: 10.3967/bes2023.018
Tra2β Enhances Cell Proliferation by Inducing the Expression of Transcription Factor SP1 in Cervical Cancer
-
Abstract:
Objective In this study, the role and potential mechanism of transformer 2β (Tra2β) in cervical cancer were explored. Methods The transcriptional data of Tra2β in patients with cervical cancer from Gene Expression Profiling Interactive Analysis (GEPIA) and cBioPortal databases were investigated. The functions of Tra2β were evaluated by using Western blot, MTT, colony formation, Transwell assays, and nude mouse tumor formation experiments. Target genes regulated by Tra2β were studied by RNA-seq. Subsequently, representative genes were selected for RT-qPCR, confocal immunofluorescence, Western blot, and rescue experiments to verify their regulatory relationship. Results The dysregulation of Tra2β in cervical cancer samples was observed. Tra2β overexpression in Siha and Hela cells enhanced cell viability and proliferation, whereas Tra2β knockdown showed the opposite effect. Alteration of Tra2β expression did not affect cell migration and invasion. Furthermore, tumor xenograft models verified that Tra2β promoted cervical cancer growth. Mechanically, Tra2β positively regulated the mRNA and protein level of SP1, which was critical for the proliferative capability of Tra2β. Conclusion This study demonstrated the important role of the Tra2β/SP1 axis in the progression of cervical cancer in vitro and in vivo, which provides a comprehensive understanding of the pathogenesis of cervical cancer. -
Key words:
- Tra2β /
- Cervical cancer /
- Proliferation /
- SP1 /
- Molecular targeted therapy
注释: -
Figure 1. Overexpression of Tra2β promoted the proliferation of Hela and Siha cells in vitro. (A) The expression of Tra2β in patients with CESC (GEPIA,
http://gepia.cancer-pku.cn/ ). (B) Correlation between Tra2β expression and tumor stage in patients with CESC (GEPIA,http://gepia.cancer-pku.cn/ ). (C) Tra2β gene expression and mutation analysis in CESC (cBioPortal,http://www.cbioportal.org/ ). (D) The confirmation of stable Tra2β overexpression in Hela and Siha cells using Western blot assay. (E) Cell proliferation ability of Tra2β overexpression in Hela and Siha cells was assessed by colony formation assays. (F) Cell viability of cells of Tra2β overexpression in Hela and Siha cells was measured by MTT assays. Two-way ANOVA with Dunnett comparison (E and F) was used. The data shown are the mean ± SD of three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.
Figure 2. Knockdown of Tra2β suppressed the proliferation of Hela and Siha cells in vitro. siRNAs targeting Tra2β were transfected into Hela and Siha cells, followed by Western blot assay (A), colony formation assays (B), and MTT assays (C). Two-way ANOVA with Dunnett comparison (B and C) was used. The data shown are the mean ± SD of three independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Figure 3. Tra2β had no effect on the migration and invasion of Hela and Siha cells in vitro. (A–D) Transwell assays were conducted to test the migratory (top) and invasive (bottom) ability of Hela cells with stable Tra2β overexpression (A and B) or Tra2β knockdown (C and D). (E–H) Transwell assays were conducted to test the migratory (top) and invasive (bottom) ability of Siha cells with stable Tra2β overexpression (E and F) or Tra2β knockdown (G and H). Five random fields of each well were counted under a microscope with a magnification of ×100 and followed by statistical analysis (B, D, F, and H). Two-way ANOVA with Dunnett comparison (B, D, F, and H) was used. Data shown are the mean ± SD of three independent experiments; ns indicates no significant difference.
Figure 4. Tra2β promoted the tumor growth of cervical cancer in vivo. (A) Formation of tumors in nude mice injected with Tra2β-overexpressed or control Hela cells. (B) Measurement of subcutaneous tumor weight. (C) Tumor growth curves of nude mice during the experiment. (D) Tra2β and Ki67 proliferation index analysis in tumors by immunohistochemistry. Magnification of (D): ×400. Two-way ANOVA with Dunnett comparison (C) and unpaired t-test (B) were used, **P < 0.01, ***P < 0.001.
Figure 5. Tra2β could regulate EGR1, SP1, and NOLC1 expression. (A) Volcano plot of global DEGs between Hela cells with stable Tra2β-overexpressed and control cells. (B–C) The function of DEGs was predicted by analysis using GO (B) and KEGG (C) by DAVID Tools (
https://david.ncifcrf.gov/ ). (D) Relative mRNA levels of Tra2β, EGR1, SP1, and NOLC1 in Hela cells with stable Tra2β-overexpressed or control cells were used to perform RT-qPCR experiments. The mRNA level of GAPDH was set as the internal control. (E) The correlation among EGR1, SP1, NOLC1, and Tra2β in 306 patients with CESC was analyzed by using the ENCORI database (http://starbase.sysu.edu.cn/index.php ). Two-way ANOVA with Dunnett comparison (D) was used, *P < 0.05, ***P< 0.001.
S2. (A) Expressions of Tra2β and SP1 were analyzed in two representative tumor tissues in nude mice formed by Hela cells. T: tumor tissues; N: adjacent normal tissues. (B) Annexin-V FITC/PI assay was used to detect apoptotic cells in Tra2β-overexpressed or control Hela cells.
-
[1] Burd EM. Human papillomavirus and cervical cancer. Clin Microbiol Rev, 2003; 16, 1−17. doi: 10.1128/CMR.16.1.1-17.2003 [2] Moore DH. Cervical cancer. Obstet Gynecol, 2006; 107, 1152−61. doi: 10.1097/01.AOG.0000215986.48590.79 [3] Zhang SK, Xu HF, Zhang LY, et al. Cervical cancer: epidemiology, risk factors and screening. Chin J Cancer Res, 2020; 32, 720−8. doi: 10.21147/j.issn.1000-9604.2020.06.05 [4] Marquina G, Manzano A, Casado A. Targeted agents in cervical cancer: beyond bevacizumab. Curr Oncol Rep, 2018; 20, 40. doi: 10.1007/s11912-018-0680-3 [5] Matsuo N, Ogawa S, Imai Y, et al. Cloning of a novel RNA binding polypeptide (RA301) induced by hypoxia/reoxygenation. J Biol Chem, 1995; 27, 28216−22. [6] Tsukamoto Y, Matsuo N, Ozawa K, et al. Expression of a novel RNA-splicing factor, RA301/Tra2β, in vascular lesions and its role in smooth muscle cell proliferation. Am J Pathol, 2001; 158, 1685−94. doi: 10.1016/S0002-9440(10)64124-7 [7] Nishikawa T, Kuwano Y, Takahara Y, et al. HnRNPA1 interacts with G-quadruplex in the TRA2B promoter and stimulates its transcription in human colon cancer cells. Sci Rep, 2019; 9, 10276. doi: 10.1038/s41598-019-46659-x [8] Mitobe Y, Iino K, Takayama KI, et al. PSF Promotes ER-positive breast cancer progression via posttranscriptional regulation of ESR1 and SCFD2. Cancer Res, 2020; 80, 2230−42. [9] Paudel D, Ouyang YQ, Huang QT, et al. Expression of TRA2B in endometrial carcinoma and its regulatory roles in endometrial carcinoma cells. Oncol Lett, 2019; 18, 2455−63. [10] Li F, Fang ZY, Zhang J, et al. Identification of TRA2B-DNAH5 fusion as a novel oncogenic driver in human lung squamous cell carcinoma. Cell Res, 2016; 26, 1149−64. doi: 10.1038/cr.2016.111 [11] Diao Y, Wu D, Dai ZJ, et al. Prognostic value of transformer 2β expression in prostate cancer. Int J Clin Exp Pathol, 2015; 8, 6967−73. [12] Gabriel B, Zur Hausen A, Bouda J, et al. Significance of nuclear hTra2-beta1 expression in cervical cancer. Acta Obstet Gynecol Scand, 2009; 88, 216−21. doi: 10.1080/00016340802503021 [13] Li MJ, Xiong D, Huang H, et al. Ezrin promotes the proliferation, migration, and invasion of ovarian cancer cells. Biomed Environ Sci, 2021; 34, 139−51. [14] Wang T, Cui YZ, Jin JJ, et al. Translating mRNAs strongly correlate to proteins in a multivariate manner and their translation ratios are phenotype specific. Nucleic Acids Res, 2013; 41, 4743−54. doi: 10.1093/nar/gkt178 [15] Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics, 2010; 26, 139−40. doi: 10.1093/bioinformatics/btp616 [16] Best A, Dagliesh C, Ehrmann I, et al. Expression of Tra2β in Cancer Cells as a Potential Contributory Factor to Neoplasia and Metastasis. Int J Cell Biol, 2013; 2013, 843781. doi: 10.1155/2013/843781 [17] Grellscheid SN, Dalgliesh C, Rozanska A, et al. Molecular design of a splicing switch responsive to the RNA binding protein Tra2β. Nucleic Acids Res, 2011; 39, 8092−104. doi: 10.1093/nar/gkr495 [18] Zhang JP, Zhang H, Wang HB, et al. Down-regulation of Sp1 suppresses cell proliferation, clonogenicity and the expressions of stem cell markers in nasopharyngeal carcinoma. J Transl Med, 2014; 12, 222. doi: 10.1186/s12967-014-0222-1 [19] Deng YR, Chen XJ, Chen W, et al. Sp1 contributes to radioresistance of cervical cancer through targeting G2/M cell cycle checkpoint CDK1. Cancer Manag Res, 2019; 11, 5835−44. doi: 10.2147/CMAR.S200907 [20] Rabachini T, Boccardo E, Andrade R, et al. HPV-16 E7 expression up-regulates phospholipase D activity and promotes rapamycin resistance in a pRB-dependent manner. BMC Cancer, 2018; 18, 485. doi: 10.1186/s12885-018-4392-8 [21] Yim EK, Park JS. The role of HPV E6 and E7 oncoproteins in HPV-associated cervical carcinogenesis. Cancer Res Treat, 2005; 37, 319−24. doi: 10.4143/crt.2005.37.6.319 [22] Yang LX, Tao T, Wang YF, et al. Knocking down the expression of TRA2β inhibits the proliferation and migration of human glioma cells. Pathol Res Pract, 2015; 211, 731−9. [23] Watermann DO, Tang YS, Zur Hausen A, et al. Splicing factor Tra2-β1 is specifically induced in breast cancer and regulates alternative splicing of the CD44 gene. Cancer Res, 2006; 66, 4774−80. doi: 10.1158/0008-5472.CAN-04-3294 [24] Kuwano Y, Nishida K, Kajita K, et al. Transformer 2β and miR-204 regulate apoptosis through competitive binding to 3' UTR of BCL2 mRNA. Cell Death Differ, 2015; 22, 815−25. doi: 10.1038/cdd.2014.176 [25] Takeo K, Kawai T, Nishida K, et al. Oxidative stress-induced alternative splicing of transformer 2β(SFRS10) and CD44 pre-mRNAs in gastric epithelial cells. Am J Physiol Cell Physiol, 2009; 297, C330−8. doi: 10.1152/ajpcell.00009.2009 [26] Cui PH, Li ZY, Li DH, et al. SP1-induced lncRNA DANCR contributes to proliferation and invasion of ovarian cancer. Kaohsiung J Med Sci, 2021; 37, 371−8. doi: 10.1002/kjm2.12316 [27] Zhang JR, Li SD, Yan Q, et al. Interferon-β induced microRNA-129-5p down-regulates HPV-18 E6 and E7 viral gene expression by targeting SP1 in cervical cancer cells. PLoS One, 2013; 8, e81366. doi: 10.1371/journal.pone.0081366 [28] Safe S, Nair V, Karki K. Metformin-induced anticancer activities: recent insights. Biol Chem, 2018; 399, 321−35. doi: 10.1515/hsz-2017-0271 [29] Beishline K, Azizkhan-Clifford J. Sp1 and the 'hallmarks of cancer'. FEBS J, 2015; 282, 224−58. doi: 10.1111/febs.13148 [30] Stoilov P, Daoud R, Nayler O, et al. Human tra2-beta1 autoregulates its protein concentration by influencing alternative splicing of its pre-mRNA. Hum Mol Genet, 2004; 13, 509−24. doi: 10.1093/hmg/ddh051 [31] Zhang WW, Sheng B, Chen SS, et al. Sevoflurane enhances proliferation, metastatic potential of cervical cancer cells via the histone deacetylase 6 modulation in vitro. Anesthesiology, 2020; 132, 1469−81. doi: 10.1097/ALN.0000000000003129 [32] Li SJ, Qi Y, Zhao JJ, et al. Characterization of nuclear localization signals (NLSs) and function of NLSs and phosphorylation of serine residues in subcellular and subnuclear localization of transformer-2β (Tra2β). J Biol Chem, 2013; 288, 8898−909. doi: 10.1074/jbc.M113.456715 [33] Liu XH, Huang XZ, Ma J, et al. 3'untranslated regions (3'UTR) of Gelsolin mRNA displays anticancer effects in non-small cell lung cancer (NSCLC) cells. Am J Cancer Res, 2021; 11, 3857−76. -
 22131Supplementary Materials.pdf
22131Supplementary Materials.pdf

-

 点击查看大图
点击查看大图
计量
- 文章访问数: 1273
- HTML全文浏览量: 574
- PDF下载量: 92
- 被引次数: 0

 Quick Links
Quick Links