

 下载:
下载:

-
Protein tyrosine phosphatase nonreceptor type 22 (PTPN22), which encodes an intracellular phosphatase protein Lyp, is preferentially expressed in hematopoietic and immune cells. The function of PTPN22 and its biological role have been studied intensely in both human and mouse models during the last decade, because the Arg-to-Trp single nucleotide polymorphism at position 620 has been identified as one of the top genetic risk factors for a variety of autoimmune diseases, including type I diabetes, rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), Hashimoto’s thyroiditis, and Grave’s disease, which are related to adaptive immunity[1]. However, only a few studies have investigated the function of PTPN22 in innate immunity, which could also contribute to autoimmune risk as well as auto-inflammation, such as seen in obesity.
Macrophages are important in obesity, which is a chronic, low-grade inflammatory disease and leads to infiltration of immune cells, including macrophages, T cells, B cells, etc., into adipose tissue. Cytokines and chemokines produced by the recruited immune cells, especially macrophages, influence the gene expression profile in adipocytes, which could lead to type II diabetes and other metabolic diseases[2-3]. Past research on PTPN22, which concentrated mostly on lymphoid cells, has identified it as a negative regulator of T-cell receptor (TCR) signaling by direct dephosphorylation of the Src family kinases as well as other key signaling molecules in T cells[4]. Recent studies in antigen-presenting cells (APCs) also gradually revealed the important roles of PTPN22 in APC function and performance, such as protein ubiquitination, cell polarization and antigen presentation, acting through the TLR signaling pathway[5-6]. Especially, 620 Trp macrophages are found to be associated with enhanced phagocytic and antigen presenting function, as well as morphological changes associated with Vav dephosphorylation[7] and higher TRAF3 lysine 63-linked ubiquitination[5].
In this study, we focused on the effect of PTPN22 619 Arg>Trp on macrophage function by investigating a high-fat diet (HFD)-induced obesity mouse model which was knocked in with either PTPN22 619 Arg or PTPN22 619 Trp allele on the endogenous mouse genetic background[7]. We describe significant effects of the mutant macrophages on adipocytes in vitro, but not in vivo on obesity or insulin resistance, contributing to the explanation of the strong contribution of this polymorphism to autoimmunity risk.
C57BL/6 mice knocked in with either PTPN22 619 Arg or PTPN22 619 Trp human allele that had been processed in our own lab were used throughout the study; the details were described in our recent paper[7]. Homozygous Ptpn22 619 Trp and Ptpn22 619 Arg littermates with C57BL/6 background were used throughout the study. All animals were housed under specific pathogen free conditions in compliance with guidelines of the Animal Care Committee of McGill University (IACUC number: A5006-01). Mice were allowed libitum access to water and a standard chow laboratory diet. Animal room was maintained under controlled conditions of temperature at 25 ± 1 °C, humidity at 55% ± 5%, and a 12 h light/12 dark cycle. For the HFD-induced obesity model, mice were fed with HFD from 6 weeks of age and lasting for16 weeks. Body weights were recorded every 7 days from the start of feeding. For the glucose tolerance test, an intraperitoneal glucose tolerance test (IP-GTT) was performed on both PTPN22 619 Arg and PTPN22 619 Trp mice in the 16th week after the start of feeding with the HFD. Mice were pre-fasted for 18 hours and injected IP with glucose at 2 g/kg body mass. Blood glucose was measured by tail bleeds at 0, 15, 30, 60, and 120 min. The insulin tolerance test (ITT) was performed similarly, except the mice were fasted for only 8 hours. Insulin was used at 1 U/kg body weight. Six mice were used from each genotype.
Mice were euthanized in their home cages with CO2 followed by cervical dislocation according to the Guidelines of the RI-MUHC animal facility, then specific organs were isolated for further procedures. When Flow Cytometry was performed, SVF cells from epididymal fad pads were isolated and incubated with F4/80-PE, CD11b-FITC and CD11c-APC (eBioscience) antibodies for 20 min at 4 °C. Cells were washed in PBS and re-suspended in fluorescence fluorescence activated cell sorting (FACS) buffer. Prepared samples were analyzed with BD FACSCalibur™. Peritoneal macrophages were obtained. Briefly, mice from chow diet feeding mice aged between 8 and 12 weeks were injected intraperitoneally with 5–10 mL ice cold PBS. The mouse peritoneum was massaged gently for about 1 min, then as much fluid as possible was collected. Cells were seeded in a 6-well plate and cultured overnight, then washed with PBS 3 times to purify macrophages by retaining only adherent cells. Cells were polarized to the M1 phenotype by exposure to 500 ng/mL LPS and 20 ng/mL IFNγ for 24 h. M2 macrophages were polarized with 10 ng/mL IL-10 and 10 ng/mL IL-13 for 24 h. Conditioned media were obtained from polarized M1/M2 macrophages by culturing with fresh media for another 24 h after PBS washing. Cells were obtained from at least six mice of each genotype.
Stromal vascular fraction SVF cells were isolated from mouse inguinal fat pads by collagenase II digestion. The digested tissue fluid was passed through a 100-µm-mesh filter and centrifuged at 1,500 rpm (300 g) for 10 min. Cells that floated on the top were decanted, and the cell pellet involving SVF cells was resuspended and washed with PBS. To differentiate into mature adipocytes, cells were culture with DMEM-F12 media supplied with 2 µg/mL Dexamethasone, 0.5 mmol/L IBMX, and 1 µg/mL insulin for 48 h, then changed to DMEM-F12 media with 1 µg/mL insulin for another two 48-h periods and followed by culturing with pure DMEM-F12 media until days 8–10. Adipocyte-macrophage co-cultures and polarized M1 macrophages were washed with PBS three times to remove LPS and IFNγ. Cells were then cultured in fresh media for another 24 h, and the culture media were collected as macrophage-conditioned media. Mature adipocytes were cultured in this conditioned media for 24 h, then cells were harvested for RNA extraction and purification. Six mice were used for each genotype, and experiments were conducted in triplicate.
For IL-6 measurement, cell media were collected and centrifuged at 300 g for 10 min to remove impurities. Media were diluted five times before carrying out ELISA using the Mouse IL-6 ELISA Kit (Sigma-Aldrich), followed by the Sandwich Assay Procedure (
https://www.sigmaaldrich.com/content/dam/sigma-aldrich/docs/Sigma/Bulletin/2/rab0520bul.pdf ). For the detailed procedure, refer to the Technical Bulletin. The intensity of the color was measured at 450 nm. Cells originated from at least six mice per genotype, and experiments were performed in triplicate.When performing q-PCR, total RNA was extracted from adipocytes and reverse-transcribed using the SuperScript® III First-Strand Synthesis kit from ThermoFisher. q-PCR reactions were performed for GLUT4, TNFα, IL-6, IL-10, and iNOS using SYBR green chemistry. Gene expression was standardized to that of cyclophilin (CYC), and the results were expressed as a ratio relative to a control. RNAs were extracted from cells from at least six mice of each genotype, and experiments were performed in triplicate. The details of q-PCR primers are shown in Supplementary Table S1 (available in www.besjournal.com).
Table S1. Quantitative real-time RT-PCR primers
Gene name Forward primer (5’-3’) Reverse Primer (5’-3’) Species GLUT4 CTT CAT CAT TGG CAT GGG TTT AGG ACC GCA AAT AGA AGG AAG A Mouse TNFα CATCTTCTCAAAATT CGAGTGACAA TGG GAG TAG ACA AGG TAC AAC CC Mouse IL-6 CTCTGGGAAATCGTGGAAA CCAGTTTGGTAGCATCCAT Mouse IL-10 GCTCTTACTGACTGGCATGAG CGCAGCTCTAGGAGCATGTG Mouse iNOS CCTGGTACGGGCATTGCT GCTCATGCGGCCTCCTTT Mouse CYC ATTTCTTTTGACTTGCGG AGACTTGAAGGGGAA Mouse Data are presented as mean values ± SEM. All experiments were repeated at least three times. Statistical analysis was performed with an unpaired t-test or two-way ANOVA. A P value < 0.05 was considered to indicate a statistically significant result.
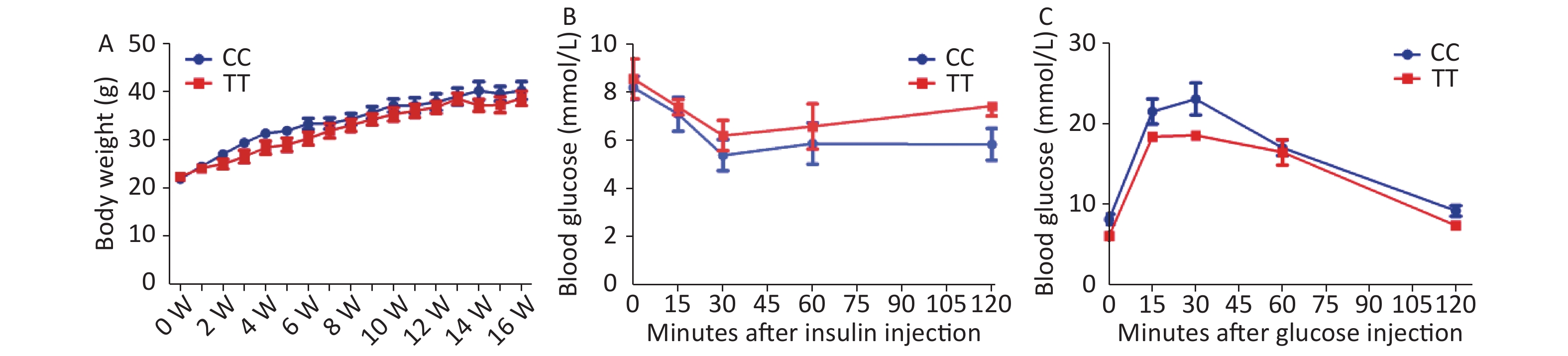
First, we found that the PTPN22619 Arg>Trp polymorphism does not influence obesity development in vivo. PTPN22 619Arg (CC) or PTPN22 619Trp (TT) male mice with the C57/BL6 background were raised individually in separate cages. Mice were placed on a HFD at 6 weeks of age and weighed weekly. As shown in Supplementary Figure S1A available in www.besjournal.com, both CC and TT mice experienced a great increase in body weight after HFD feeding, but little difference was seen between the two genotypes. In week 15, an insulin tolerance test (ITT) was performed after the mice were fasted for 8 hours (Supplementary Figure S1B). The mice were given 1 week to recover, then a glucose tolerance test (GTT) was conducted after 18 h of fasting (Supplementary Figure S1C). The blood glucose levels of both genotypes changed according to insulin or glucose injection, but no significant differences were seen. With our sample size and the observed standard deviations, we had 80% power to detect a difference of 1.5 g body weight and 1 and 0.8 mmol/L, respectively, at a significance level of 0.05, for the OGTT and ITT. These results show that the PTPN22 619 Arg>Trp polymorphism has no effect on HFD-induced obesity and related metabolic disorder in vivo. However, our in vivo findings are concordant with a series of human studies. A recent study investigated the PTPN22 1858C/T polymorphism in the susceptibility of adolescents to obesity and found no association between the two[8].
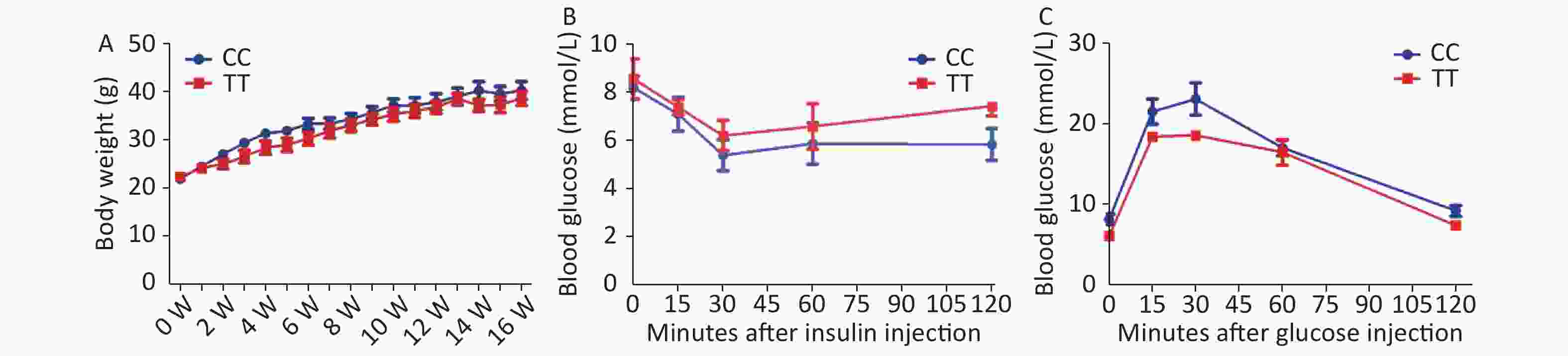
Figure S1. IP-GTT, IP-ITT and body weight in PTPN22619Arg or PTPN22 619Trp mice.
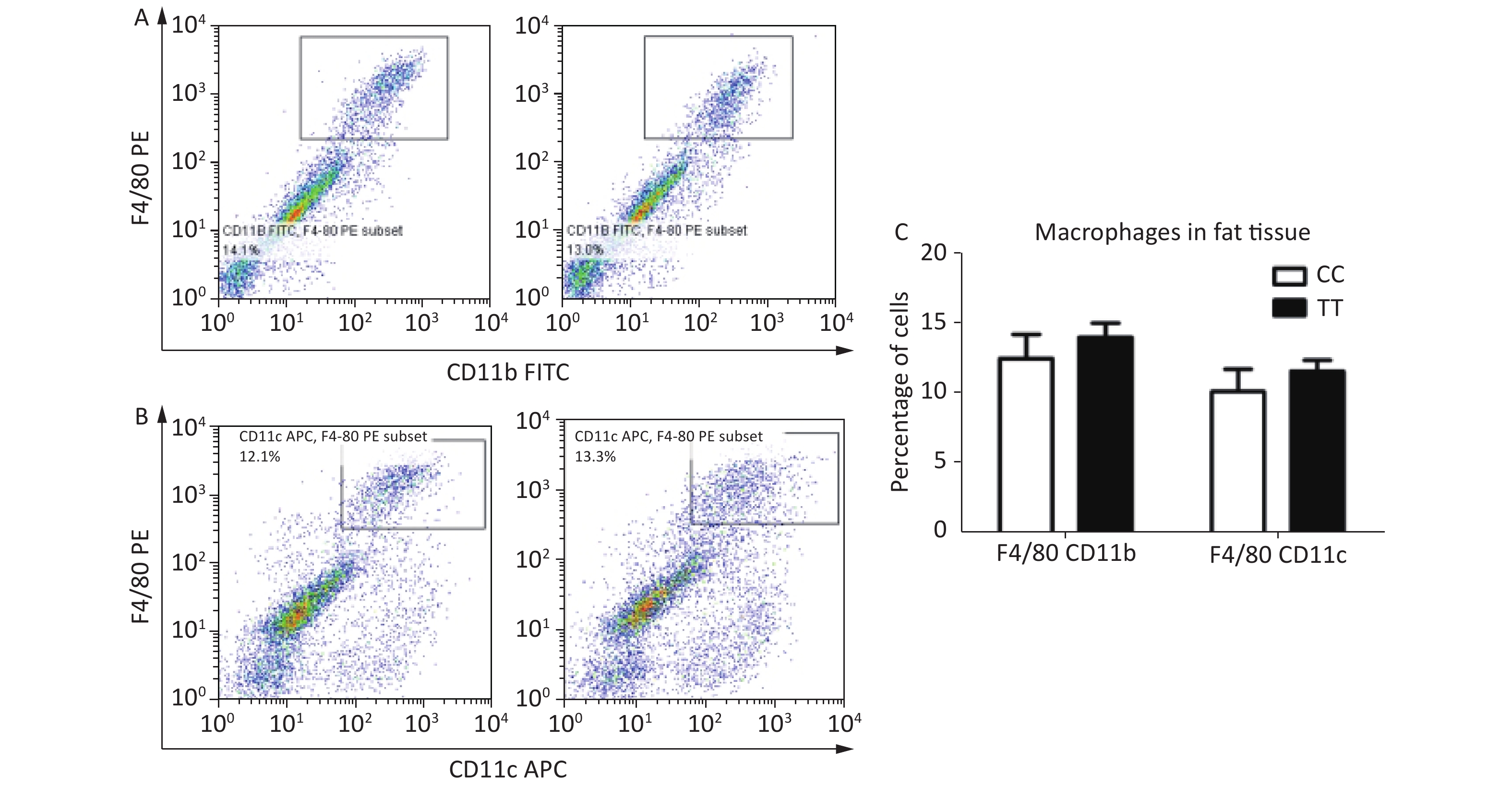
Second, cell populations of ATMs are not affected by the PTPN22 619 Arg>Trp polymorphism. In mice, ATMs express the antigens F4/80 and CD11b; CD11c+ macrophages are seen as proinflammatory M1 macrophages, which largely increase in obesity[9-10]. To identify the macrophage population in adipose tissue, mice with the CC or TT genotype were sacrificed in week 16 after HFD feeding, and SVF cells from epididymal fat pads were stained with the F4/80, CD11b, and CD11c antibodies for flow cytometry. Total ATMs, represented by F4/80+ CD11b+, were gated as Figure 1A. F4/80+ CD11c+ were gated as the population of M1 macrophages (Figure 1B). The percentages of both the F4/80+ CD11b+ and the F4/80+ CD11c+ population were quantified and analyzed, but no significant differences were found between CC and TT (Figure 1C). This means that in diet-induced obese mice, though there is macrophage infiltration and an increased number of proinflammatory M1 macrophages, the 619 Arg>Trp polymorphism of PTPN22 does not influence these processes.
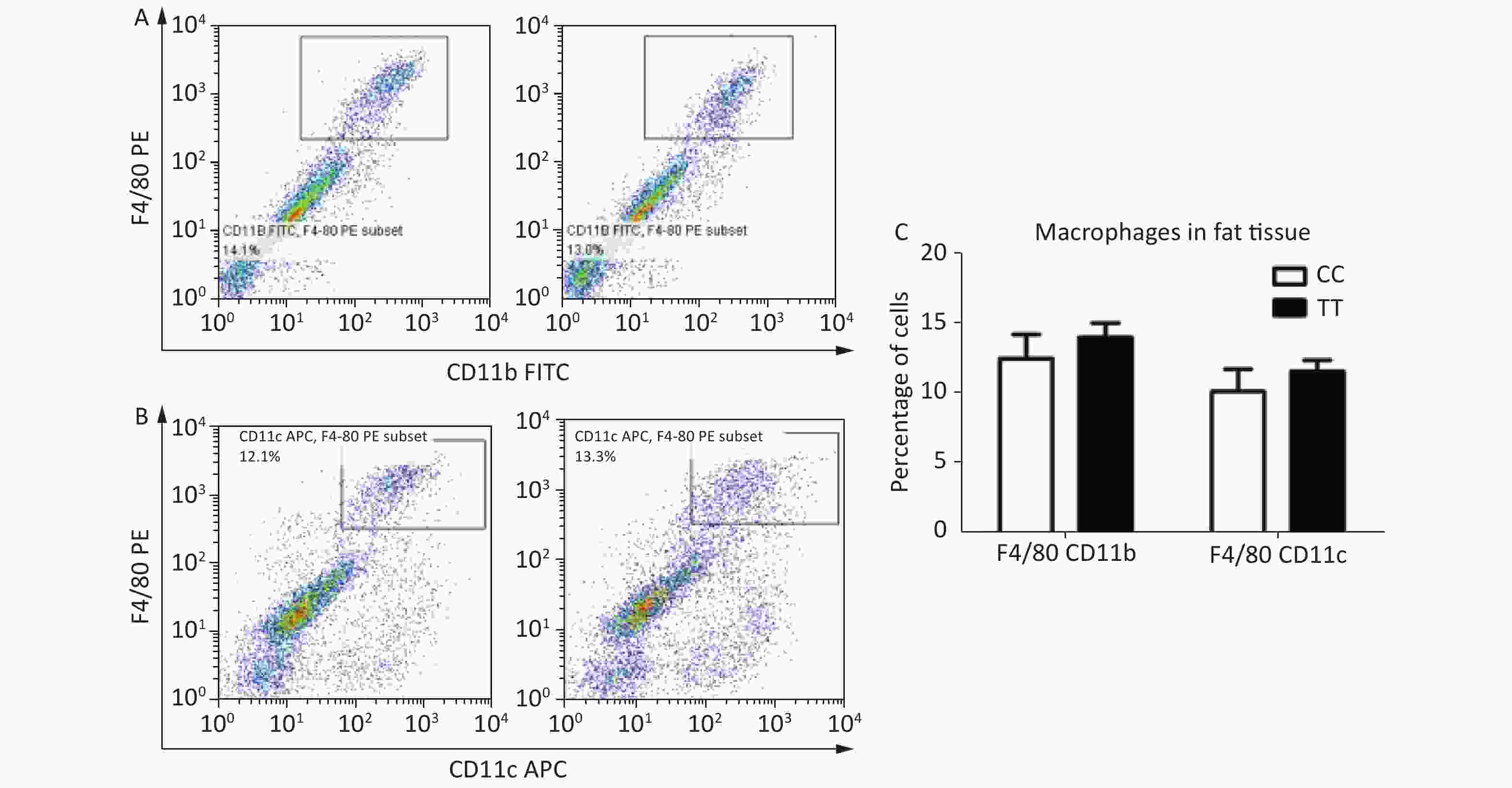
Figure 1. Macrophage populations in adipose tissue.
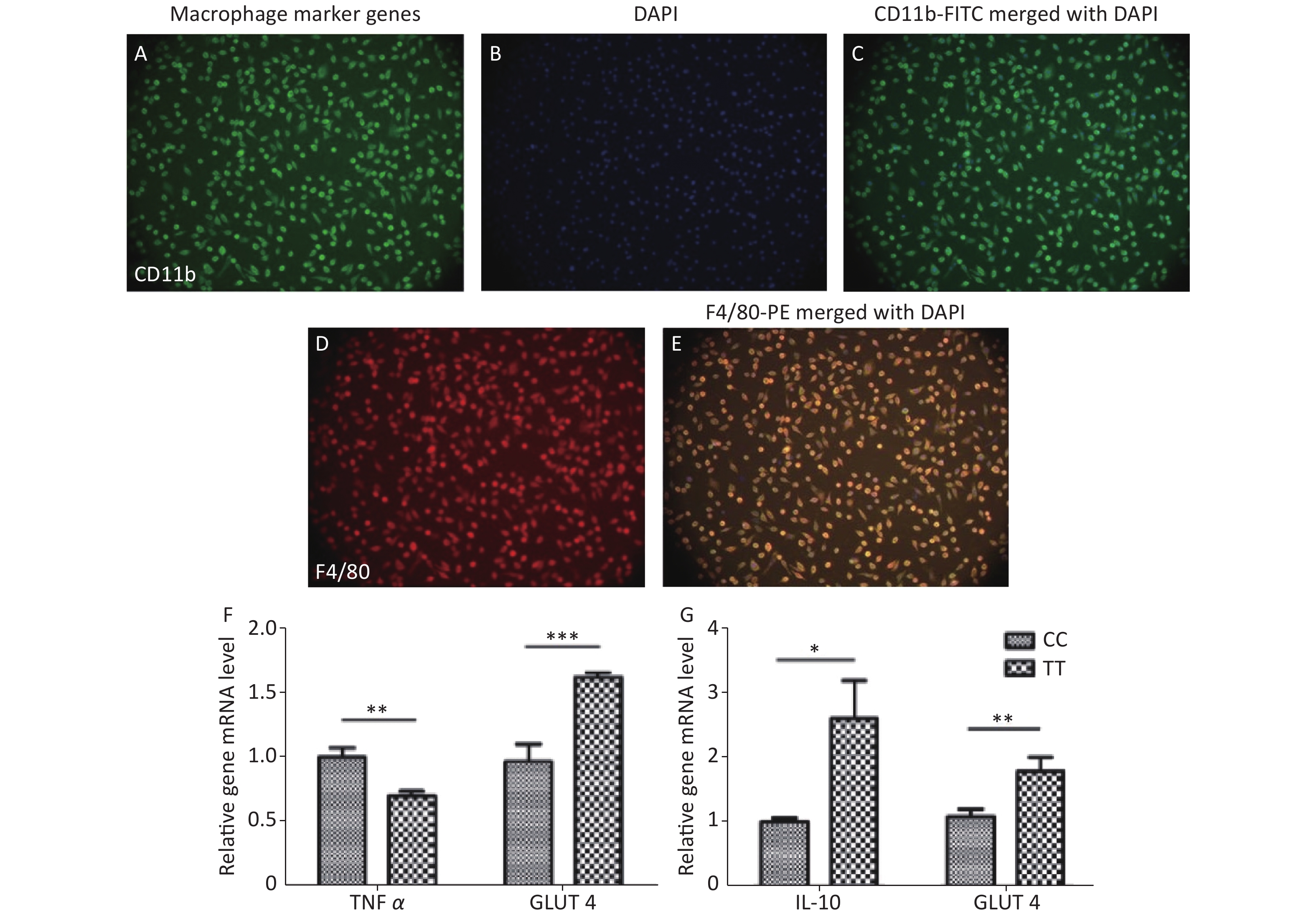
Third, adipocyte homeostasis was improved after co-culturing with PTPN22 619 Trp macrophages. In obese individuals, macrophages are recruited to the adipose tissue (AT), ultimately leading to the remodeling of AT by indirect/secreted and direct/cell contact-mediated factors. As introduced previously, PTPN22 is reported to play an important role in macrophage polarization. Therefore, we investigated whether the PTPN22 619 Arg>Trp polymorphism in macrophages could modulate adipocyte homeostasis. Peritoneal macrophages were prepared and purified by attachment. As shown in Figure 2A–E, more than 95% of the total cells are macrophages, which are CD11b and F4/80 positive. Mature adipocytes were induced from CC-genotyped pre-adipocytes (Supplementary Figure S2 available in www.besjournal.com). Then macrophages of both genotypes were polarized into M1 or M2 macrophages, and the conditioned media were used to culture mature adipocytes. Inflammatory factor TNFα had a reduced expression level in adipocytes co-cultured with Trp-allele M1 macrophages, compared with adipocytes cultured with wildtype macrophages, whereas adipocytes cultured with Trp-allele M2 macrophages showed higher transcription of the anti-inflammatory factor IL-10. The expression of glucose transporter GLUT4 was elevated in adipocytes co-cultured with both Trp M1 and M2 macrophages (Figure 2F–G). These data prove that the PTPN22 619 Arg>Trp polymorphism in macrophages has a positive effect on adipocyte homeostasis.
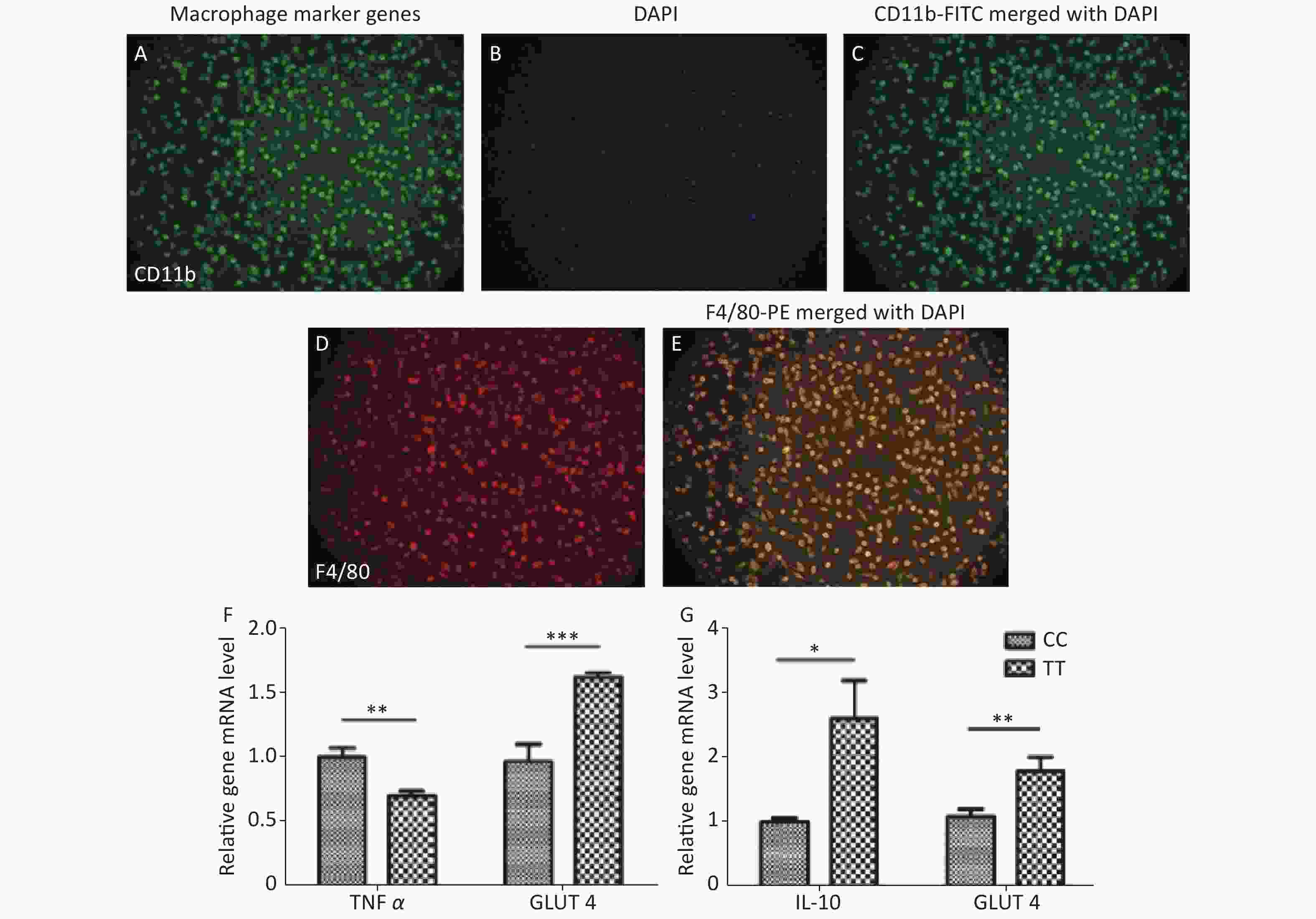
Figure 2. Macrophage and adipocyte co-culture.
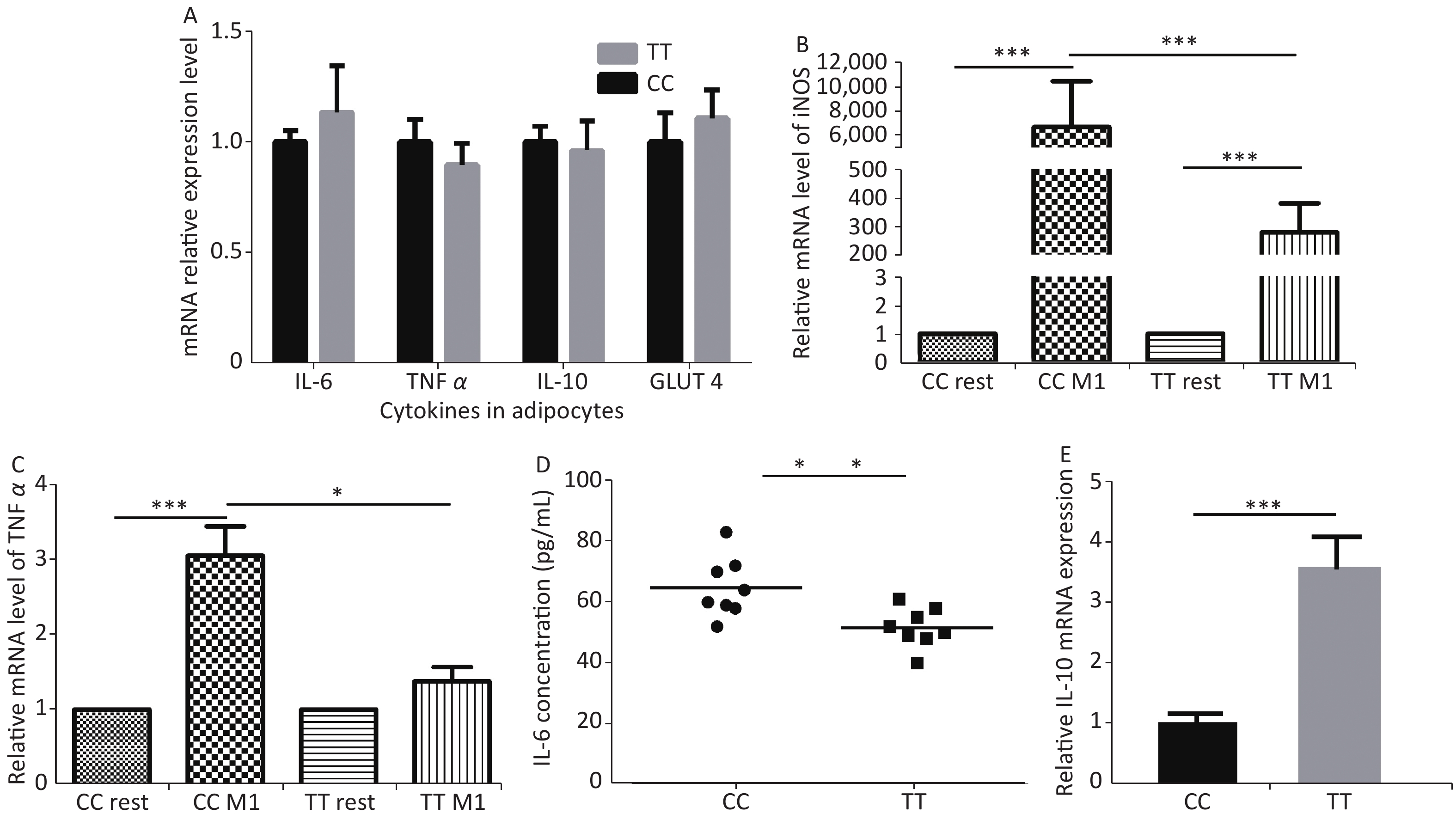
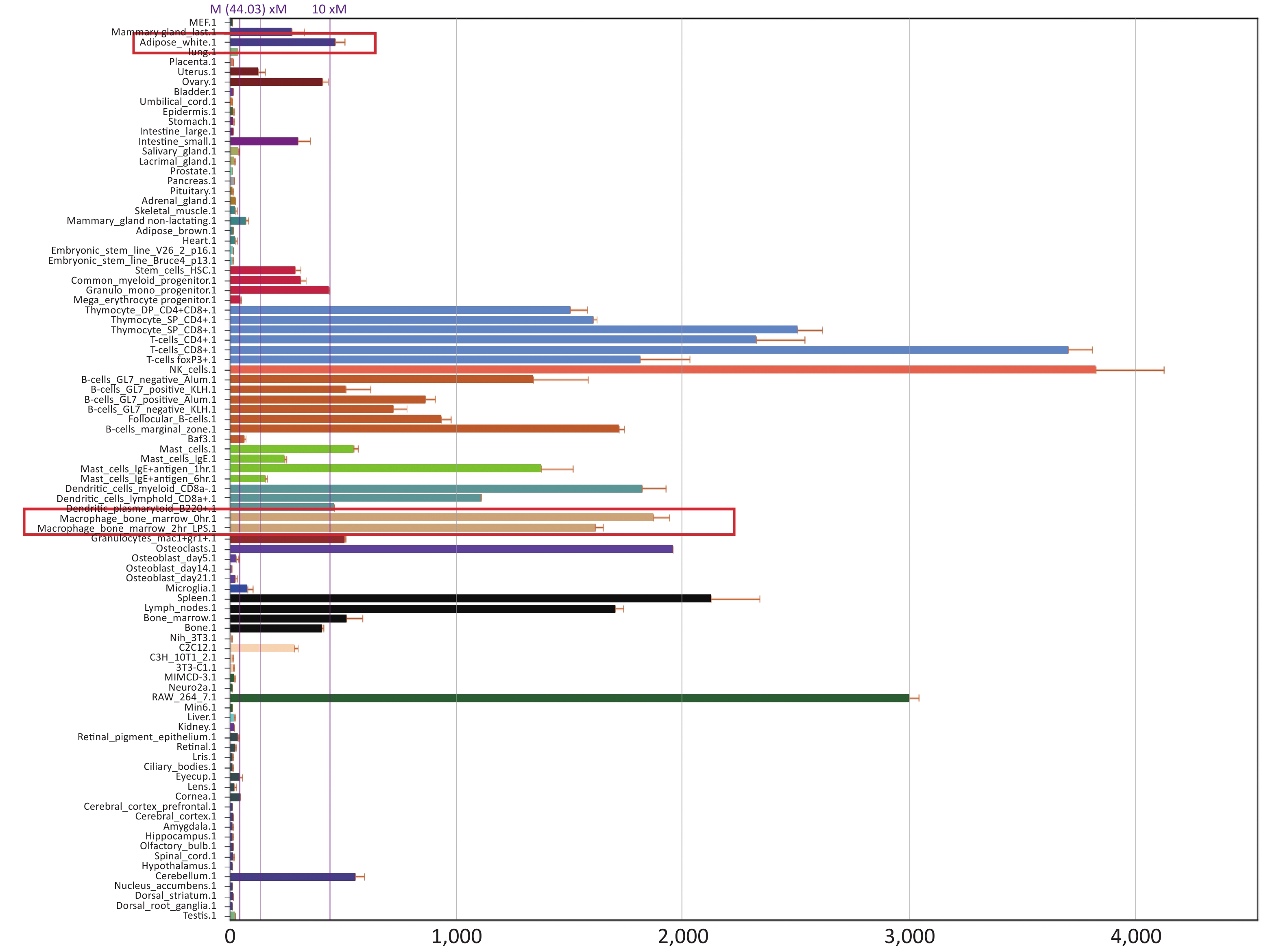
Fourth, the reason for improved adipocytes homeostasis is the altered cytokine/chemokine expression in PTPN22 619 Arg>Trp polymorphic macrophages. Human lymphoid-specific phosphatase (LYP) protein is restrictively expressed in immune cells, such as lymphocytes, dendritic cells, and macrophages (Supplementary Figure S3 available in www.besjournal.com from Bio-GPS). We presume, with the low expression in adipose tissue, this polymorphism would not alter gene expression in adipocytes. Its ability to improve adipocyte homeostasis could be due to altered gene expression in macrophages. Therefore, we examined related gene expression in both adipocytes and macrophages. Pre-adipocytes from both genotypes were collected and differentiated as previously described. Then, RT-qPCR was used to detect the expression of inflammatory and glucose transporter genes in mature adipocytes. None of these genes, including IL-6, TNFα, IL-10, and GLUT 4, were influenced by the Trp allele (Figure 3A). Then, we monitored gene expression in macrophages. Peritoneal macrophages from both genotypes were polarized into M1 macrophages or M2 macrophages as previously described. In M1 macrophages, as expected, both genotypes had elevated iNOS and TNFα expression levels compared with resting macrophages, but the Trp allele significantly reduced iNOS, TNFα, and mRNA expression (Figure 3B–C) and IL-6 secretion (Figure 3D) compared with the wild type, while strongly inducing the IL-10 mRNA level in M2 macrophages (Figure 3E). The above data show that the 619 Arg>Trp polymorphism of PTPN22 does not influence the expression of inflammatory cytokines in adipocytes, but positively prevents pro-inflammatory cytokine expression in M1 macrophages and enhances the expression of anti-inflammatory cytokines. Combined with result 3.3, these results confirmed our hypothesis that the positive effects of the Trp allele in adipocyte glucose homeostasis is through an indirect influence of modulation of macrophage cytokine expression.
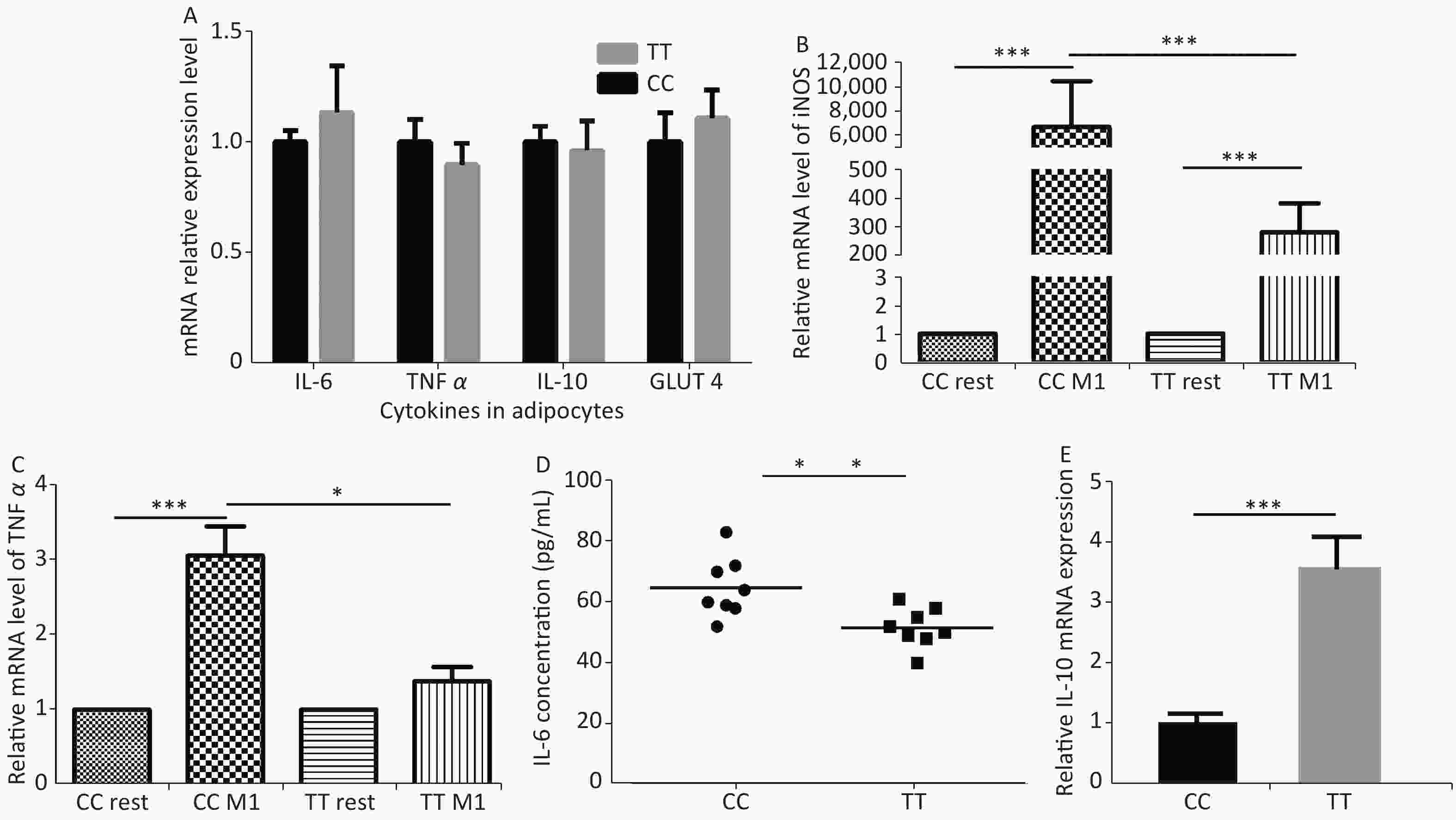
Figure 3. Cytokine expression in adipocytes and macrophages.
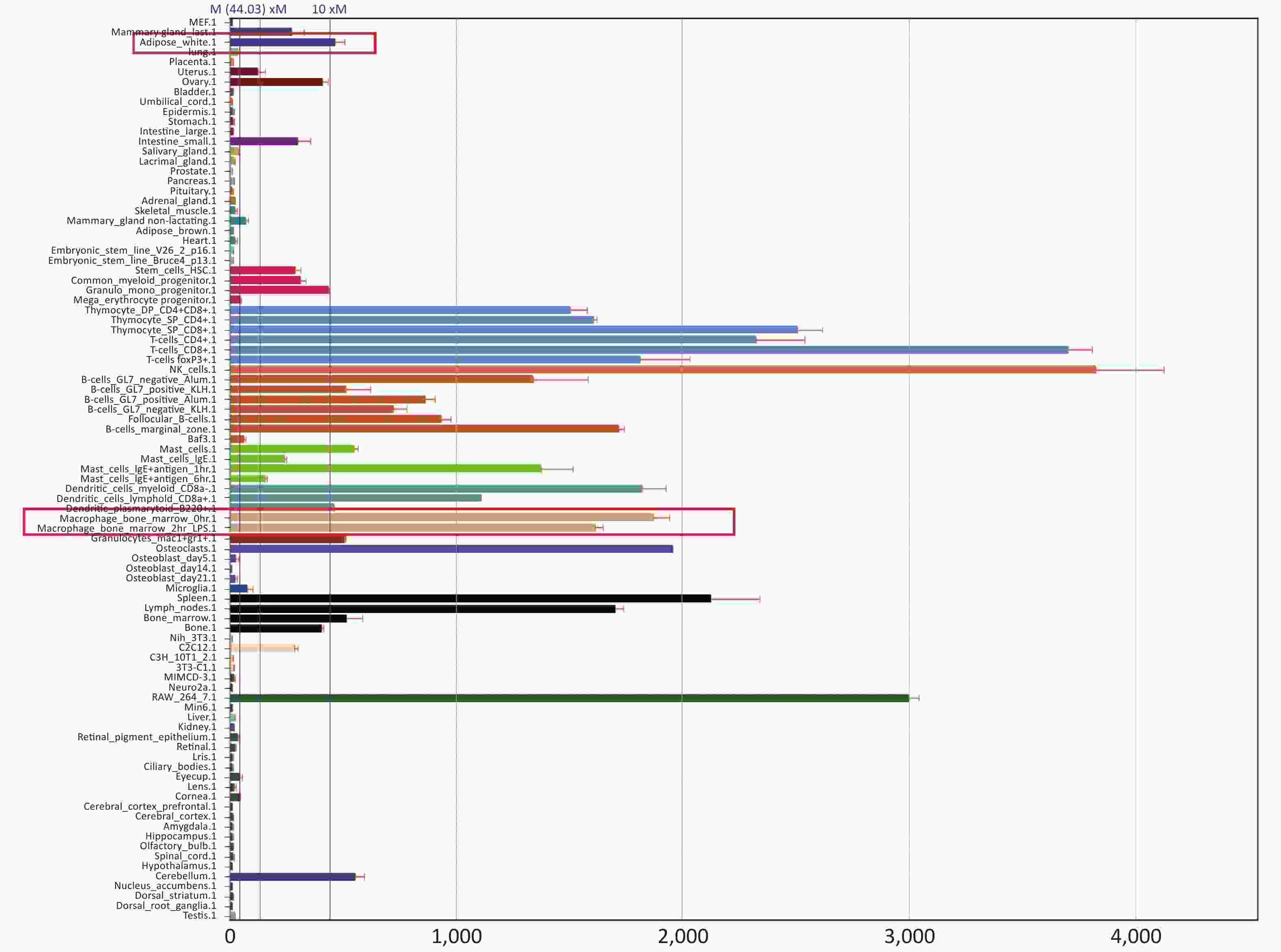
Figure S3. PTPN22 expression profile.
As a conclusion, a strong positive effect was found in adipocytes co-cultured with PTPN22 Trp/Trp macrophages because of differential cytokine expression in macrophages, though no association between the variant and obesity development was observed in vivo. This could be due to compensatory adjustment, which commonly acts to maintain health status when a slight change occurs in vivo.
Ethics Approval and Consent to Participate This work has the approval from Animal Care committee of McGill University.
Consent for Publication Not applicable.
Availability of Data and Material All data generated or analyzed during this study are included in this published article.
Competing Interests We have read and understood BMC Genetics policy on declaration of interests and declare that we have no competing interests.
Authors' Contributions Mei Hang developed the study concept, did the experiments and wrote the manuscript. SUN Chao participated in the study design. SUN Yuan Chao, NIU Yu Juan, and WU Chuan Hong participated in the manuscript writing. Constantin Polychronakos supervised this study and helped to correct the English writing. All Authors read and approved the manuscript.
Acknowledgement We greatly appreciate the assistance of stuff in Lyman Duff Animal Facility (McGill University). We also thank Rosemarie Grabs, Marylene Rousseau, Natalija Popovic and Xiaoyu Du for the technical assistance.
Figure S2. Microscopy of mature adipocyte.
doi: 10.3967/bes2021.031
-
-
S1. IP-GTT, IP-ITT and body weight in PTPN22619Arg or PTPN22 619Trp mice.
Body weight were recorded every week since starting HFD intake (A). IP-ITT (1 U/kg) and IP-GTT (glucose 2 g/kg): Male mice after feeding HFD for 15 to 16 weeks were used. Blood glucose were assessed after insulin injection (B) and glucose injection (C). n = 6. All values are means ± SE.
Figure 1. Macrophage populations in adipose tissue.
Epididymal SVF cells from both CC and TT mice were gated for F4/80+ CD11b+ as the total macrophage (A) and F4/80+ CD11c+ proinflammatory M1 macrophage (B) populations. (C) Quantification of F4/80+ CD11b+ and F4/80+ CD11c+ population. n = 6. All values are means ± SE.
Figure 2. Macrophage and adipocyte co-culture.
Peritoneal macrophages were stained with CD11b-FITC (A), DAPI (B), and F4/80-PE (D), then identified under a fluorescent microscope (20 x 100 objective), (C) and (E) show CD11b-FITC and F4/80-PE merged with DAPI, respectively. Macrophages had a purification level higher than 95%. TNFα and GLUT4 mRNA transcription level in adipocytes are shown in (F) after M1 conditioned-media-treatment. (G) IL-10 and GLUT4 mRNA transcription in adipocytes after M2 macrophage co-culturing. All data were normalized against wild type. (*P < 0.05, **P < 0.01, ***P < 0.005). n = 6. All values are means ± SE.
Figure 3. Cytokine expression in adipocytes and macrophages.
Adipocytes from both genotypes were induced to mature adipocytes and IL-6, TNFα, IL-10, and GLUT 4 mRNA levels were detected (A). iNOS and TNFα mRNA levels (B–C) and IL6 protein levels (D) from M1 macrophages and IL-10 mRNA levels from M2 macrophages (E). *P < 0.05, **P < 0.01, ***P < 0.005. n = 6. All values are means ± SE.
S3. PTPN22 expression profile.
Searching result from Bio-GPS shows PTPN22 expression profile in different tissues. Expression in adipocyte and macrophage were highlighted in red.
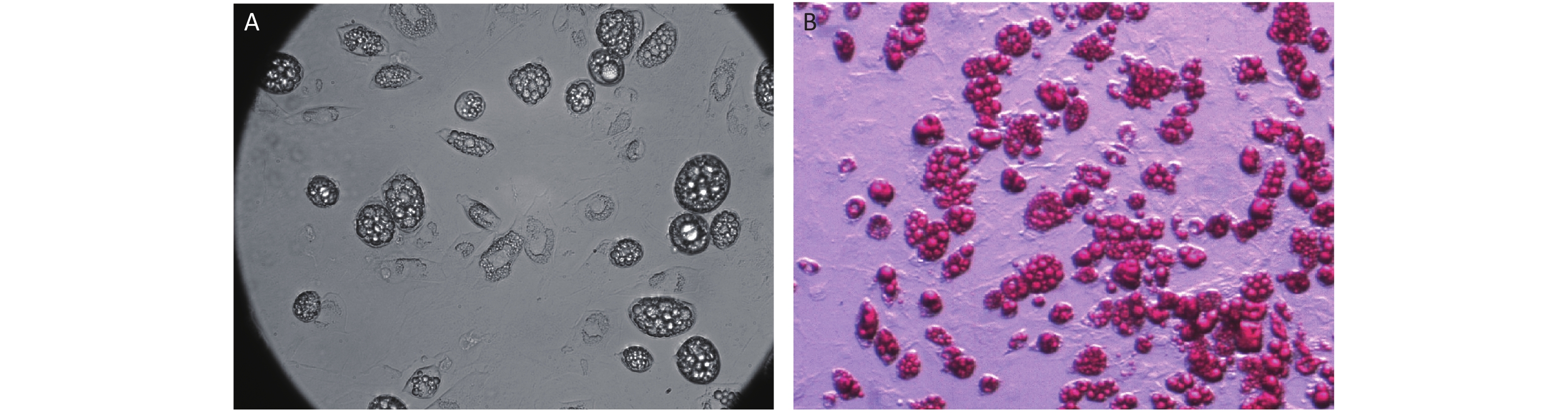
S2. Microscopy of mature adipocyte.
Adipocytes from inguinal fat pad were used as the source of preadipocytes. D10 differentiated adipocyte were directly microscoped (A) or microscoped after Oil Red O staining (B) at 40 x 100 objective.
S1. Quantitative real-time RT-PCR primers
Gene name Forward primer (5’-3’) Reverse Primer (5’-3’) Species GLUT4 CTT CAT CAT TGG CAT GGG TTT AGG ACC GCA AAT AGA AGG AAG A Mouse TNFα CATCTTCTCAAAATT CGAGTGACAA TGG GAG TAG ACA AGG TAC AAC CC Mouse IL-6 CTCTGGGAAATCGTGGAAA CCAGTTTGGTAGCATCCAT Mouse IL-10 GCTCTTACTGACTGGCATGAG CGCAGCTCTAGGAGCATGTG Mouse iNOS CCTGGTACGGGCATTGCT GCTCATGCGGCCTCCTTT Mouse CYC ATTTCTTTTGACTTGCGG AGACTTGAAGGGGAA Mouse  下载: 导出CSV
下载: 导出CSV
-
[1] Fousteri G, SN Liossis, M Battaglia. Roles of the protein tyrosine phosphatase PTPN22 in immunity and autoimmunity. Clin Immunol, 2013; 149, 556−65. doi: 10.1016/j.clim.2013.10.006 [2] Krischer JP, X Liu, K Vehik, et al. Predicting Islet Cell Autoimmunity and Type 1 Diabetes: An 8-Year TEDDY Study Progress Report. Diabetes Care, 2019. [3] Ferrante AW Jr. The immune cells in adipose tissue. Diabetes Obes Metab, 2013; 15(Suppl 3), 34−8. [4] Yu X, JP Sun, Y He, et al. Structure, inhibitor, and regulatory mechanism of Lyp, a lymphoid-specific tyrosine phosphatase implicated in autoimmune diseases. Proc Natl Acad Sci USA, 2007; 104, 19767−72. doi: 10.1073/pnas.0706233104 [5] Wang Y, I Shaked, SM Stanford, et al. The autoimmunity-associated gene PTPN22 potentiates toll-like receptor-driven, type 1 interferon-dependent immunity. Immunity, 2013; 39, 111−22. doi: 10.1016/j.immuni.2013.06.013 [6] Clarke F, HA Purvis, C Sanchez-Blanco, et al. The protein tyrosine phosphatase PTPN22 negatively regulates presentation of immune complex derived antigens. Sci Rep, 2018; 8, 12692. doi: 10.1038/s41598-018-31179-x [7] Li M, H Beauchemin, N Popovic, et al. The common, autoimmunity-predisposing 620Arg > Trp variant of PTPN22 modulates macrophage function and morphology. J Autoimmun, 2017. [8] Salinas-Santander MA, RB Leon-Cachon, AC Cepeda-Nieto, et al. Assessment of biochemical parameters and characterization of alpha -308G/A and +1858C/T gene polymorphisms in the risk of obesity in adolescents. Biomed Rep, 2016; 4, 107−111. doi: 10.3892/br.2015.534 [9] Lumeng, CN, JL Bodzin, AR Saltiel. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J Clin Invest, 2007; 117, 175−84. doi: 10.1172/JCI29881 [10] Lumeng CN, JB DelProposto, DJ Westcott, et al. Phenotypic switching of adipose tissue macrophages with obesity is generated by spatiotemporal differences in macrophage subtypes. Diabetes, 2008; 57, 3239−46. doi: 10.2337/db08-0872 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 907
- HTML全文浏览量: 396
- PDF下载量: 42
- 被引次数: 0

 Quick Links
Quick Links