

 下载:
下载:




-
Sepsis is a life-threatening organic dysfunction caused by an unregulated host response to infection. Each year the mortality of global sepsis-related cases (50 million) was approximately 22% (11 million); more than 20 million cases were among children under the age of age 5[1], nearly 4 million deaths (20%) across the world per year occurred due to neonatal sepsis infections[2]. Gram-negative infection is the common etiology of sepsis[3], primarily caused by Escherichia coli, Haemophilus influenzae, Klebsiella spp., and Salmonella spp. The zoonotic potential of Escherichia coli or Salmonella spp. from chicken-sourced food products was important to define for public health purposes. Salmonella enterica Typhimurium and Salmonella enterica Enteritidis are the leading serovars responsible for human and animal salmonellosis globally, with the majority of human cases originating from foodborne outbreaks[4].
Myocardial dysfunction is a serious consequence of sepsis and contributes to high mortality. Endotoxin or lipopolysaccharide (LPS) from Gram-negative bacteria causes septic myocardial dysfunction with severe inflammation responses[5] and complications, whereas the therapeutic approaches for this condition are limited. Quercetin, an active substance in various herbal medicines, exerts preventive and therapeutic effects in cardiovascular diseases. One study found that quercetin blocked the reduction of superoxide dismutase and catalase enzyme levels of rat hearts after LPS induction[6]. Another study indicated that quercetin suppressed the production of proinflammatory cytokines tumor necrosis factor alpha (TNFα) and interleukin-1β (IL-1β), and it also improved the cardiac function of LPS-induced mice[7]. Therefore, quercetin had antioxidation and anti-inflammatory effects in rodents. The LPS was detected by one of pattern recognition receptors such as toll like receptor 4 (TLR4), and activated the release of inflammatory factors, including TNFα, IL-1β, and interleukin-6 (IL-6), causing inflammatory cell infiltration[8]. One study indicated that inflammatory mediators nuclear factor kappa B subunit 1 (NF-κB1) and TNFα increased after LPS from S. Typhimurium induction in the myocardiocytes of chicken embryos[9]. However, the cardioprotective mechanism of quercetin on cardiac inflammation after LPS induction in chicken embryos remains unclear. Tight junctions are essential cellular barriers that separate extracellular compartments and allow passive transport of ions, solutes, and water in a regulated manner. Tight junction complex consists of occludin, Claudins, ZO-1(Zonula occludens-1), and subjacent adherens junctions[10]. Head-to-head interactions between Claudins, occludin, and ZO-1 on adjacent cells form the basis for paracellular channels that restrict permeability. Nevertheless, the protective effects of quercetin to tight junction complex after LPS induction remain elusive.
Autophagy, a cellular housekeeping and evolutionarily conserved process in all eukaryotic cells, is essential to maintain tissue homeostasis by degrading damaged organelles and aggregated proteins via lysosomal-mediated systems, particularly in long-lived cells such as myocardiocytes[11] and neuronal cells[12]. Autophagy occurs in physical and pathological conditions such as starvation, energy deficiency, hypoxia, hyperpyrexia, exercise, infection and endoplasmic reticulum stress[13]. There are three main types of autophagy: macroautophagy, microphagy, and chaperone-mediated autophagy. Autophagy plays a crucial pole in homeostasis and energy balance. Peroxisome proliferator-activated receptor Autophagy is tightly regulated by posttranscriptional signaling mechanisms involving mTOR1, AMPK, sirtuins, anti-autophagy protein (Bcl-2), and transcriptional regulation by factors such as peroxisome proliferator-activated receptor alpha (PPARα), NF-κB1. PPARs is the main executor of fatty acid β-oxidation. Apolipoprotein 4 (APOA4) is involved in absorption of the fatty acid, fat metabolism, and the glucose homeostasis, promoted the glucose absorption for adipose cells, and resisted inflammation and arteriosclerosis[14]. Sodium glucose co-transporter1 (SGLT1) play a crucial role in the metabolism of glucose. Nevertheless, there is no report on cardiac PPARα, APOA4, and SGLT1 after LPS induction in chicken embryos, it will discuss quercetin ameliorate the energy deficiency associated expressions of PPARα, APOA4, and SGLT1 in the present study.
Autophagy is involved in initiation, elongation, closure, and maturation. Elongation, formation of a double membrane structure, was comprised of an autophagy related gene 5(ATG5)-12-6 complex, LC3-I (Microtubule associated protein 1 light chain 3), and LC3-II. LC3-I is converted to LC3-II via proteolytic cleavage of its C-terminus by ATG4. ATG5 is covalently bound to ATG12 in the presence of ATG10. The products released by autophagy are monosaccharide, disaccharide, amino acids, and free fatty acids which are utilized for other cellular processes. Activation of autophagy at appropriate levels is essential for maintaining cardiac function in physiological conditions. Insufficient autophagy triggers accumulation of toxic materials, organelles dysfunction, and eventually heart failure. Autophagy augmentation and its abnormal modulation by inflammation may lead to cardiovascular functional decline and heart failure[15]. Some phytochemicals are as autophagy-related modulators. Previous studies indicated that kaempferol[16] and berberine[17] could affect and modulate autophagy; Nevertheless, the protective mechanism of quercetin modulating autophagy in the heart inflammation after LPS induction is unclear.
Apoptosis (from the Greek meaning “falling off”) is genetically programmed cellular suicide and involves the coordinated dismantling of intracellular components designed to prevent inflammation and limit damage to the surrounding environment, leading to the cell’s disintegration into apoptotic bodies[18]. Apoptosis is a coordinated and self-killing process. There are two major apoptotic pathways: the extrinsic death receptor pathway and intrinsic mitochondrial pathway[19]. When the apoptosis signaling pathway is activated, cells begin to shrink. Plasma membrane blebbing occurs and tightly encloses the packed organelles to form apoptotic bodies. Cytoskeletal and nuclear proteins are degraded and cleaved by caspases during the execution phase of apoptosis[20]. Caspase 3 (CASP3) plays an essential role in apoptosis-related signaling pathways. Apoptosis is induced by death receptors, mitochondria, and endoplasmic reticulum signaling pathways. The extrinsic signaling pathway is involved in death receptors and ligands, including the Fas cell surface death receptor (Fas) and the Fas ligand, and TNFα and TNF receptor 1. They transmit death signals from the cell surface to intracellular pathway through the Fas-associated death domain and the unubiquitinated receptor interacting serine/threonine kinase 1 (RIPK1) and receptor interacting serine/threonine kinase 3 (RIPK3), which activate the procaspase 8 and form a death-inducing signal complex. The intrinsic signaling pathway initiates from mitochondria and trigger apoptosis. LPS can promote the opening of mitochondrial permeability transition pores, impede the mitochondrial transmembrane potential, and activate CASP3. B cell CLL/lymphoma 2 (Bcl-2) controls permeability of the mitochondria membrane and then regulates the release of cytochrome C. Cytochrome C binds and activates procaspase 9, which in turn promotes CASP9, CASP3 and CASP7 activation[21].
Pyroptosis is a unique form of proinflammatory programmed cell death in sepsis[22]. Pyroptosis rapidly and irreversibly forms membrane pores to allow ions to enter the cell, resulting in osmotic swelling and ruptures, and DNA cleavage and fragmentation. The signaling pathways of pyroptosis are mainly divided into canonical pathways mediated by caspase-1 and noncanonical pathways mediated by caspase-11[23]. Inflammasome activation promotes the maturation and release of IL-1β and IL-18, cleavage of gasdermin D, and development of pyroptosis[24]. CASP1 converts pro-IL-1β to IL-1β. Inflammasome initiates when LPS activates pattern recognition receptors TLR4 and NOD-like receptors like NLRP3. TLR4 activates NF-κB1 and myeloid differentiation primary response 88 (MYD88) expressions and induces NLRP3 and pro-IL-1β release. One study indicated that vitamin C alleviates LPS-induced myocardial injury by inhibiting pyroptosis via the ROS-AKT/mTOR signalling pathway[25]. Another study suggested that irisin suppressed inflammation, apoptosis, and pyroptosis by blocking the TLR4 and NLRP3 inflammasome signalings to mitigate myocardial dysfunction in sepsis[26]. However, quercetin attenuates programmed cell death in cardiac inflammation after LPS induction remains elusive.
Evidence indicate that the interplay between the autophagy and apoptosis pathways is critical in the pathogenesis of diabetic cardiomyopathy[27] and coronary heart disease[21]. However, the complex interplay of cardiac inflammation, autophagy, pyroptosis, and apoptosis after LPS induction in the chicken embryos model remains unclear. In the present study, LPS was used to establish the septic cardiac dysfunction model of chicken embryos. In addition, the protective effects of quercetin on inflammatory response, autophagy, programmed cell death, and myocardocytes permeability were studied.
-
LPS from Salmonella enterica serotype Typhimurium (S. Typhimurium, product number: L7261, Sigma-Aldrich Trading Co. Ltd., Shanghai, China) was dissolved in a phosphate-buffered solution (PBS) at 2.5 μg/mL (500 ng/egg). Quercetin (Product number: Q4591, Sigma-Aldrich Trading Co. Ltd., Shanghai, China) was dissolved in 100% ethanol at 50, 100, or 200 μmol/L (10, 20, or 40 nmol/egg).
Specific pathogen-free embryos (weight 56.76 ± 3.32 g) were provided by a chicken breeder (Jinan SAIS Poultry Co. Ltd., Jinan, Shandong, China). The fertilized eggs were individually weighed and divided into ten groups, each group consisting of 4 replicates with 3 eggs per replicate. There were untreated or injected with 0.2 mL/egg of PBS vehicle, LPS (500 ng/egg; 0.2 mL/egg), PBS + ethanol vehicle (0.2 mL each per egg), quercetin + LPS (10, 20, or 40 nmol + LPS 500 ng/egg), and quercetin (10, 20, or 40 nmol/egg) groups. The incubation, weighting, examination, injection of the treatment solution, and taken samples was the same as our previous study[28].
-
The protocols of histopathology were the same as our previous study[28].
-
The protocols of RT-qPCR were the same as our previous study[28]. A total of thirty-one genes were selected to show the cardiac inflammatory factors, autophagy, apoptosis, and pyroptosis (Table 1). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the housekeeping gene. The relative levels of target mRNA expression were calculated using the 2−∆∆Ct method.
Table 1. Primer used in real-time quantitative polymerase chain reaction
Genes name Primer sequence (5’–3’) Gene bank ID APOA4 F:GAGGTACTTCACTGAGCTGGG R:TGAGCTGCTTGGTGATCTCAG NM_204938 AMPKα1 F:CGGCAGATAAACAGAAGCACGAG; R:CGATTCAGGATCTTCACTGCAAC NM_001039603 AMPKα2 F:CATGGACGTGTTGAAGAGGCAG; R:TTCTCTGGTTTCAGGTCCCTGTGG NM_001039605 ATG5 F:CCCATCCCTGGTCCGTAAC; R:CGGCGGCGTATACGAAGTA NM_001006409 ATG7 F:TGCAGTTTGCTCCCTTCAGT R:TGGGAAACCTGATGGATCGC NM_001396468 Bcl-2 F: TGGCTGCTTTACTCTTGGGG; R:TATCTCGCGGTTGTCGTAGC NM_205339 Beclin-1 F:CCGCTGAAGCTCGATACCTC R:TTCTGGCTGGTGGGATGAAC NM_001006332.1 CASP1 F:CACTTCCACTTCGGATGGCT; R:CCACGAGACAGTATCAGGCG XM_015295935 CASP3 F:ACCGAGATACCGGACTGTCA; R:GCCATGGCTTAGCAACACAC NM_204725 CASP12 F:AATAGTGGGCATCTGGGTCA; R:CGGTGTGATTTAGACCCGTAAGAC [97] Claudin 1 F:CTGGGTCTGGTTGGTGTGTT; R:CGAGCCACTCTGTTGCCATA NM_001013611 Fas F:GTCAGTGCTGCACGAAATGT; R:AACCTCCAAACCGAGTGCTT NM_001199487 Drp1 F: GGCAGTCACAGCAGCTAACA; R:GCATCCATGAGATCCAGCTT NM_001079722 GAPDH F: GAGAAACCAGCCAAGTATGATG; R: CACAGGAGACAACCTGGTCC NM_204305 IFNγ F:CTGACAAGTCAAAGCCGCAC; R:CTTCACGCCATCAGGAAGGT NM_205149 IL-1β F:GCTCAACATTGCGCTGTACC; R:AGGCGGTAGAAGATGAAGCG FJ537850 IL-6 F:ACGAGGAGAAATGCCTGACG; R:CTTCAGATTGGCGAGGAGGG NM_204628 IL-8 F:TGCCAGTGCATTAGCACTCA; R:TTGGCGTCAGCTTCACATCT HM179639 IL-10 F:TGCGAGAAGAGGAGCAAAGC R:AACTCCCCCATGGCTTTGTAG AJ621254 LC3B F:CTTCTTCCTCCTGGTGAACG; R:GCACTCCGAAAGTCTCCTGA NM_001031461 MMP9 F:ACACAGACTCTATGCTGCCTG; R:GAGAGTAGGGCGGGGAAAAT NM_204667 MMP3 F:ATCAGGCTCTACAGTGGTG; R:ATGGGATACATCAAGGCAC XM_025152201 MYD88 F:TTAGTCTTTCCCCAGGGGCT; R:GCCAGTCTTGTCCAGAACCA NM_001030962 NF-κB1 F:TCAACGCAGGACCTAAAGACAT; R:GCAGATAGCCAAGTTCAGGATG NM_001396396 Occludin F:TACATCATGGGCGTCAACCC R:CCAGATCTTACTGCGCGTCT NM_205128 PPARα F:AACGGAGTTCCAATCGCTGG R:GGATGCTGGTGAAAGGGTGT XM_025150258 RIPK1 F:GATCCATTTGCGAAGCTGCC R:CTTAGGCTAATGGCGCTGGT NM_204402 SGLT1 F:GGCCAGGGCTTACTATTGGT R:AGCTTCAGGTATCCGCACAT AJ236903 TLR4 F:GGCTCAACCTCACGTTGGTA; R:AGTCCGTTCTGAAATGCCGT KP410249 ZO-1 F:TATGAAGATCGTGCGCCTCC; R:GAGGTCTGCCATCGTAGCTC XM_015278977 TNFα F:CCCATCCCTGGTCCGTAAC; R:CGGCGGCGTATACGAAGTA MF000729 Note. APOA4: apolipoprotein 4; AMPKα1: Protein kinase AMP-activated catalytic subunit alpha 1; AMPKα2: Protein kinase AMP-activated catalytic subunit alpha 2; ATG5: Autophagy related gene 5; ATG7: Autophagy related gene 7; Bcl-2: B cell CLL/lymphoma 2; CASP1: Caspase 1; CASP3: Caspase 3; CASP12: Caspase 12; Drp1: dynamin 1 like; GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; Fas: Fas cell surface death receptor; IFNγ: Interferon gamma; IL-8: Interleukin-8; IL-1β: Interleukin-1β; IL-6: Interleukin-6; IL-10: Interleukin-10; LC3B: Microtubule associated protein 1 light chain 3 beta; MMP3: Matrix metallopeptidase 3; MMP9: Matrix metallopeptidase 9; MYD88: Myeloid differentiation primary response 88; NF-κB1: Nuclear factor kappa B subunit 1; RIPK1: receptor interacting serine/threonine kinase 1; PPARα: Peroxisome proliferator-activated receptor alpha; SGLT1: Sodium glucose co-transporter1; TLR4: Toll like receptor 4; ZO-1: Zonula occludens-1; TNFα: Tumor necrosis factor alpha. -
Immunohistochemical investigations were carried out using indirect method of peroxidase with a primary antibody specific for CASP1 (caspase 1, anti-CASP1, GB11383, Servicebio, Wuhan servicebio technology CO., LTD), CASP3 (caspase 3, anti-CASP3, GB11532, Servicebio), AMPKα2 (protein kinase AMP-activated catalytic subunit alpha 2, anti-AMPKα2, GB113685, Servicebio), Claudin 1 (anti-Claudin 1, GB113685, Servicebio), LC3-II/I (anti-LC3-II/I, GB11124, Servicebio), MMP9 (matrix metallopeptidase 9, anti-MMP9, GB11132, Servicebio), and TLR4 (anti-TLR4, GB11519, Servicebio). The protocols of immunohistochemiscal investigation were the same as our previous study[28].
-
The protocols of Western blotting analysis were the same as our previous study[28]. Specific antibodies were used to detect the following proteins: TLR4 (1:3000, GB11519, Servicebio), Interferon gamma (IFNγ, 1:3000, GB11107-1, Servicebio), MMP9 (1:3000, GB12132-1, Servicebio), MMP3 (matrix metallopeptidase 3, 1:3000, GB11131, Servicebio), LC3-II/I(1:3000, Servicebio) CASP1 (1:3000, Servicebio), CASP3 (1:3000, GB11383, Servicebio), AMPKα1(Protein kinase AMP- activated catalytic subunit alpha 1, 1:3000, 66536-1-IG, Proteintech, Wuhan, Hubei, China), GAPDH (1:2000, GB15002, Servicebio), and the secondary IgG HRP conjugated-antibody (1:5000; Servicebio). The expression levels of the target proteins were normalized to GAPDH.
-
All data were statistically analyzed using the paired t test and ANOVA method by the SPSS software (version 16.0, SPSS Inc., Chicago, Illinois, USA), and were presented as mean ± standard deviation. Differences at P < 0.01 were indicated as significant.
-
Based on the histopathological changes, effects of quercetin on LPS-induced cardiac inflammation were presented in the chicken embryos. There was no significant change and no inflammatory response in the control group, the PBS group, the (PBS + ethanol) group, and the quercetin group (Figure 1A–F). They all showed normal morphology. Nevertheless, the obvious inflammatory change included myocardial fiber swelling and ruptures, and there was inflammatory cell infiltration (heterophils accumulated) in the hearts of the LPS group (Figure 1G). No inflammatory cell infiltration was presented in the (LPS + Q) group (Figure 1H–J).
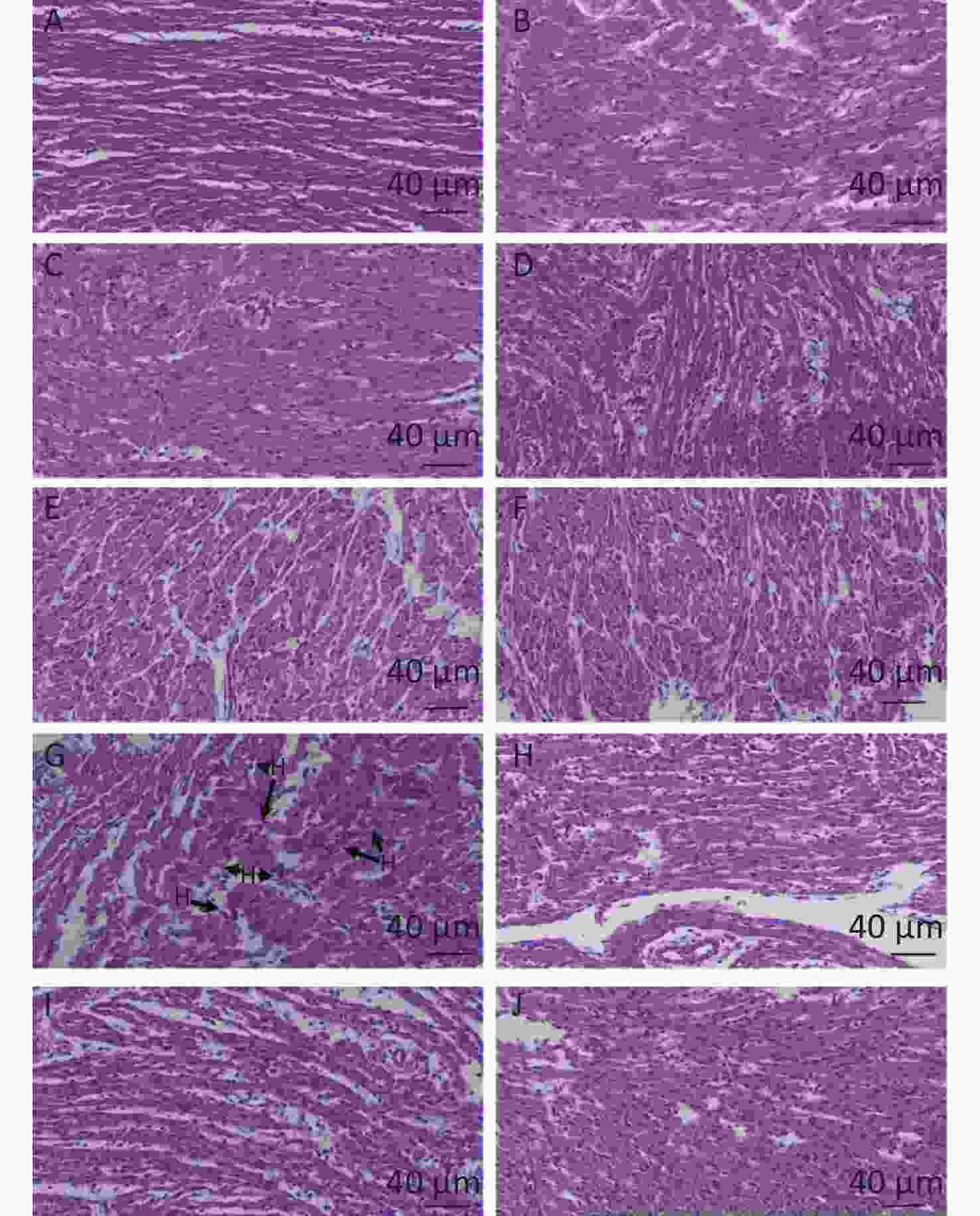
Figure 1. The histopathologic changes of the hearts induced by LPS and quercetin in chicken embryos A, control group; B, PBS group; C, PBS + ethanol group; D–F, 10 nmol, 20 nmol or 40 nmol quercetin; G, treatment group (500 ng/egg); H–J, 500 ng LPS +(10 nmol, 20 nmol, or 40 nmol). Scale bar: 40 μm. HB: Heterophilc banded cells, arrow.
-
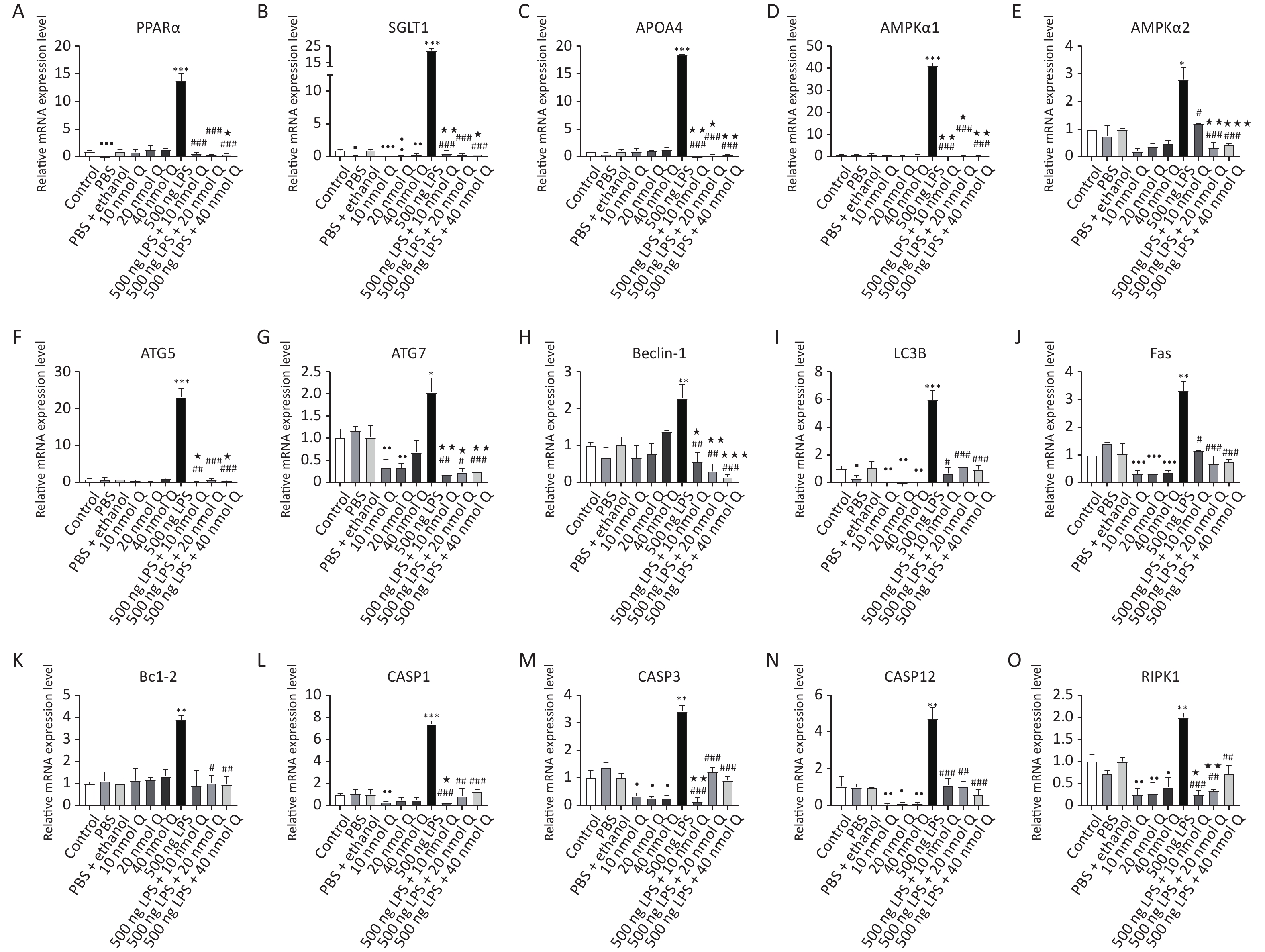
There was no different mRNA expression in inflammatory-related factors when compared the PBS group or the (PBS + ethanol) group with the control group, respectively. Quercetin significantly downregulated the mRNA expression of TNFα, IL-6, IL-8, p38, and MMP9 when compared with the (PBS + ethanol) group at three dosages (P < 0.05, P < 0.01, or P < 0.001). Quercetin with 10 nmol and 20 nmol significantly downregulated the mRNA expression of TLR4 when compared with that of the (PBS + ethanol) group (P < 0.001). Quercetin with 20 nmol significantly downregulated both the mRNA expression of IFNγ and IL-1β when compared with that of the (PBS + ethanol) group (P < 0.05). Quercetin with 20 nmol and 40 nmol significantly downregulated the mRNA expression of MMP3 when compared with that of the (PBS + ethanol) group (P < 0.05). Quercetin significantly upregulated the mRNA expression of IL-10 when compared with that of (PBS + ethanol) group, whether it was 10 nmol, 20 nmol, or 40 nmol (P < 0.05, P < 0.01, or P < 0.001). (Supplementary Figure S1A–K, available in www.besjournal.com).
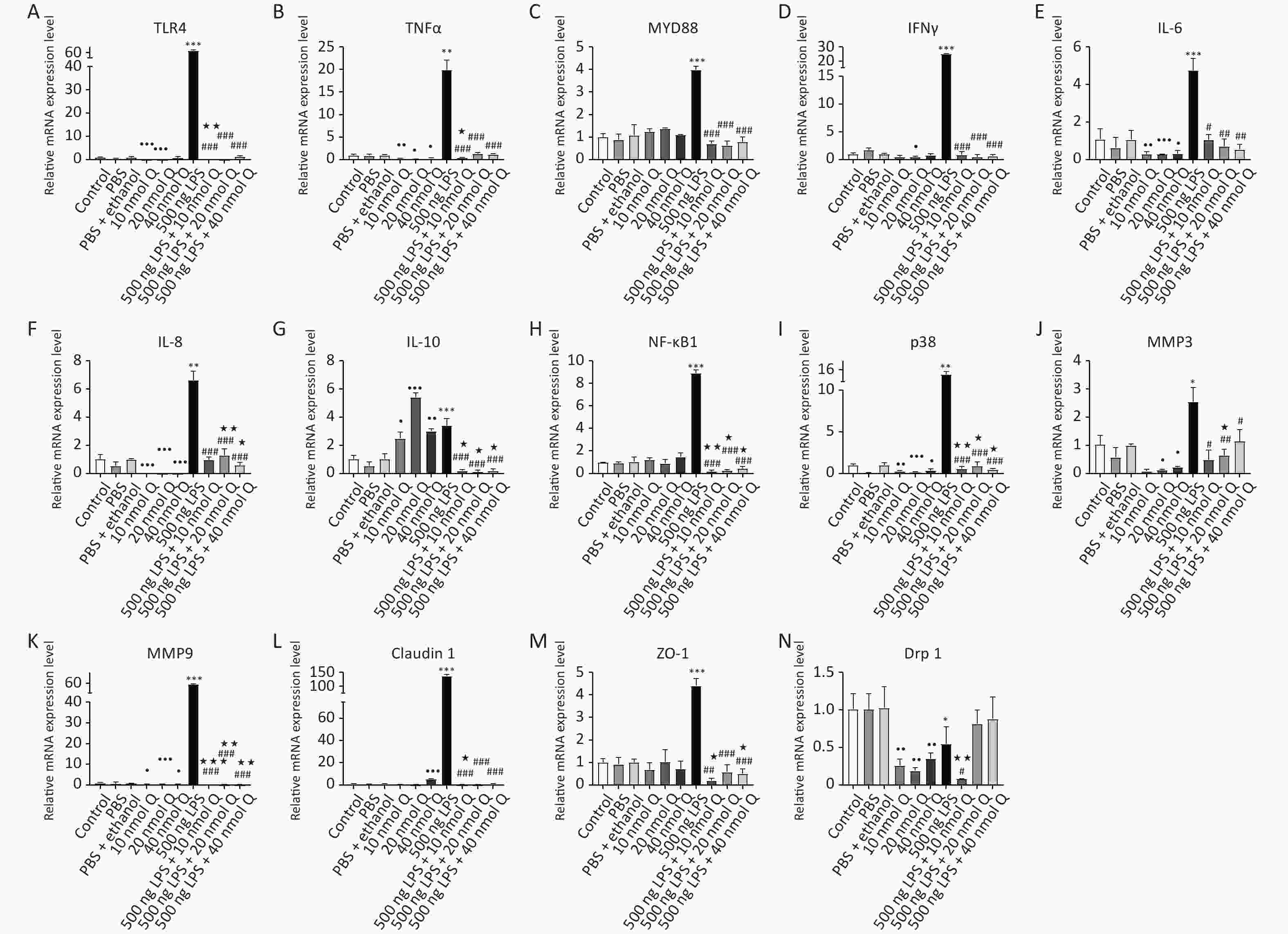
Figure S1. Quercetin alleviates the mRNA expressions of inflammatory associated factors induced by LPS in the hearts of chicken embryos. Data are presented as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; #Indicates the significant differences between the LPS group and the (LPS + Q) group; ★Indicates the significant differences between the (PBS + ethanol) group and the (LPS + Q) group; ●Indicates the significant differences between the (PBS + ethanol) group and the Q (quercetin) group.
LPS induced pattern recognition receptor TLR4, inflammatory cytokines including IFNγ, IL-1β, IL-6, IL-10, TNFα, and MYD88 mRNA expression increase by 68.6 folds, 25.0 folds, 204.6 folds, 4.8 folds, 3.4 folds, 19.9 folds, and 4.0 folds when compared with that of PBS group, respectively (P < 0.05, P < 0.01, or P < 0.001); as well as chemokines IL-8, mitogen activated protein kinases p38, and nuclear factor NF-κB1 mRNA expression increase by 6.6 folds, 13.8 folds, and 8.9 folds, respectively (P < 0.01 or P < 0.001). Quercetin significantly blocked the LPS-induced expression of these inflammation-related factors at all doses (P < 0.05, P < 0.01, or P < 0.001). Quercetin also suppressed the upregulation of matrix metallopeptidases including MMP3 and MMP9 (increased by 2.5 folds and 58.2 folds, respectively) by LPS (P < 0.05 or P < 0.001), and reduced the mRNA expression below the vehicle controls (P < 0.05, P < 0.01, or P < 0.001).
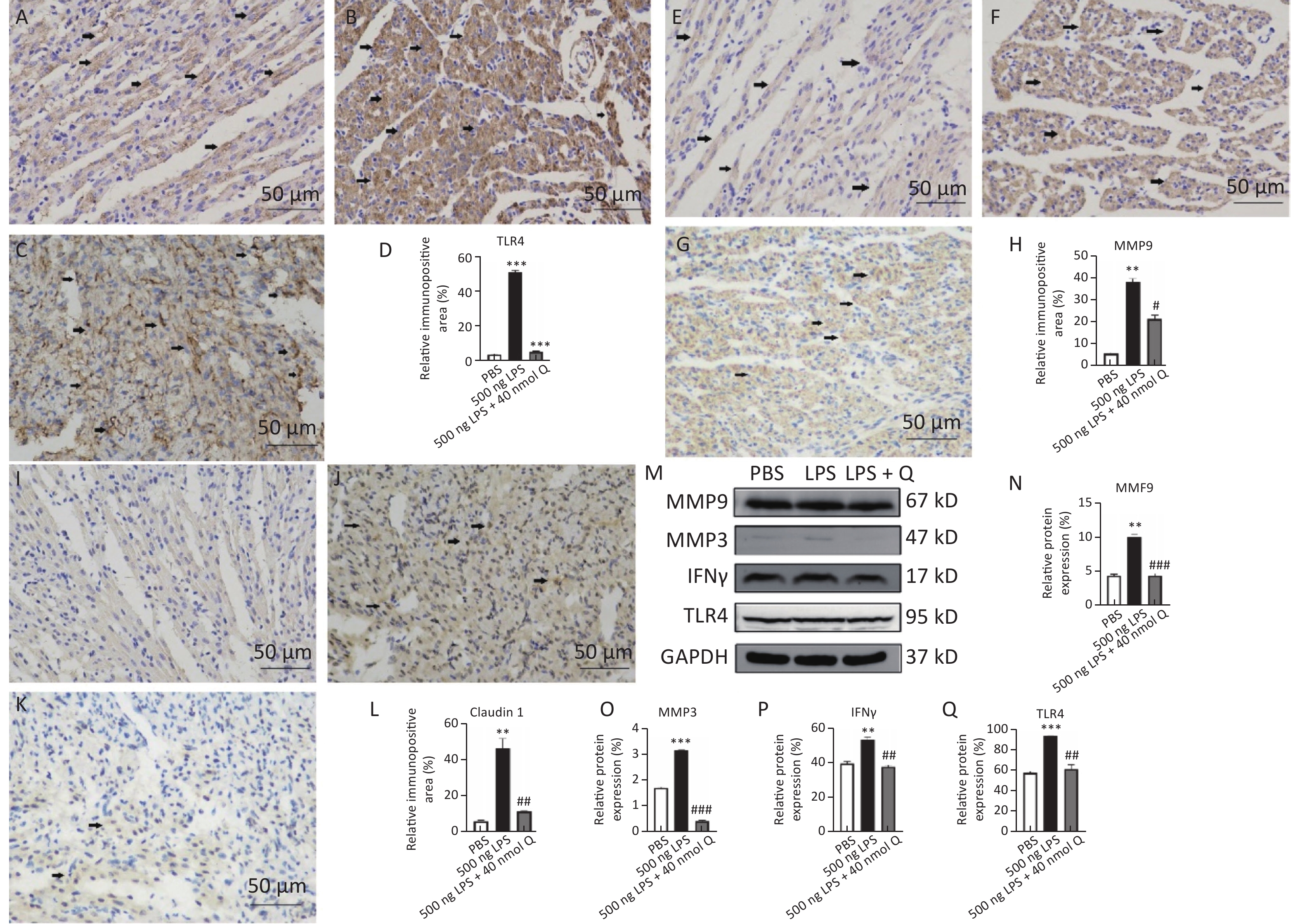
The immunopositivity of TLR4 and MMP9 in the cytoplasm of myocardiocytes significantly increased after LPS induction when compared with that of the PBS group (P < 0.01 or P < 0.001), whereas the immunopositivity to TLR4 and MMP9 in the treatment group decreased when compared with that of the LPS group (P < 0.05 or P < 0.001) (Figure 2). The protein expressions of TLR4, IFNγ, MMP3, and MMP9 significantly increased after LPS induction when compared with that of the PBS group (P < 0.01 or P < 0.001), whereas quercetin could significantly decrease these expressions compared with the LPS group (P < 0.01 or P < 0.001). These findings revealed quercetin attenuated LPS-induced cardiac inflammation.
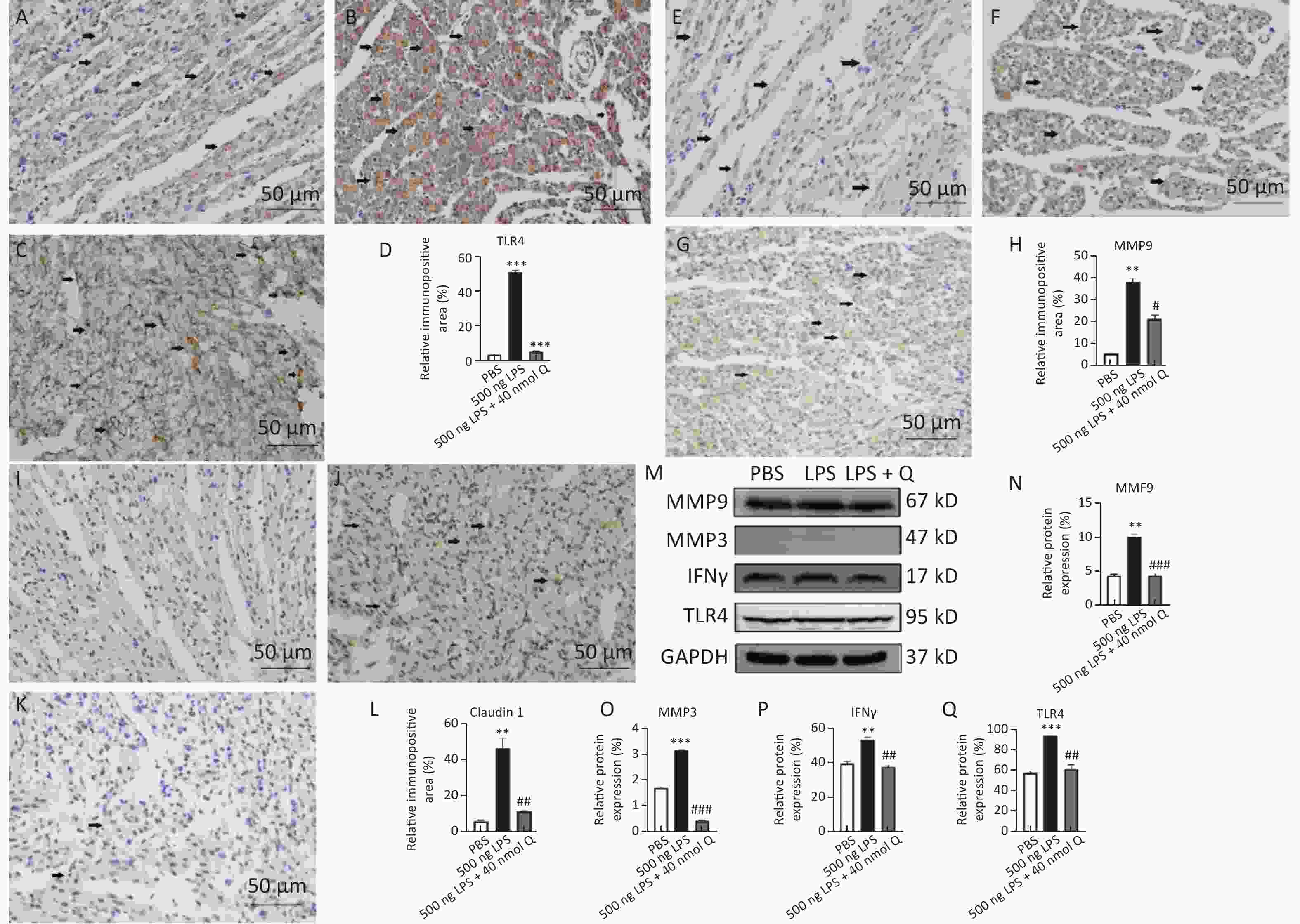
Figure 2. Quercetin alleviates the protein expression of TLR4, MMP9, Claudin 1, IFNγ, and MMP3 induced by LPS in the hearts of chicken embryos by immunohistochemistry and Western blotting. A, E, I: PBS group of TLR4, MMP9, Claudin 1, respectively; B, F, J: LPS group (500 ng LPS/egg) of TLR4, MMP9, Claudin 1, respectively; C, G, K, treatment group (500 ng LPS/egg + 40 nmol quercetin/egg) of TLR4, MMP9, Claudin 1, respectively; D, H, L, relative immunopositive area of TLR4, MMP9, Claudin 1, respectively. Immunopositivity to TLR4, MMP9, Claudin 1 (arrow, brown to yellow). Scale bar: 50 μm. M, the protein expression of TLR4, IFNγ, MMP3 and MMP9; N, relative protein expression of MMP9; O, relative protein expression of MMP3; P, relative protein expression of IFNγ; Q, relative protein expression of TLR4. Data were expressed as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; **P < 0.01; ***P < 0.001. #indicates the significant differences between the LPS group and the (LPS + Q) group; Q, Quercetin; #P < 0.05; ##P < 0.01; ###P < 0.001.
-
Zonula occludens-1 (ZO-1) and Claudin 1 are major components of cardiac tight junction tight proteins. LPS significantly increased the mRNA expression of Claudin 1 and ZO-1 by 137.3 folds and 4.4 folds (P < 0.001), respectively (
Supplementary Figure S1L–M ). Quercetin significantly decreased Claudin 1 and ZO-1 expression when compared with vehicle groups after LPS induction at three doses (10–40 nmol/egg) (P < 0.01 or P < 0.001). The immunopositivity of Claudin 1 in the cytoplasms of myocardiocytes significantly increased after LPS induction when compared with that of the PBS group (P < 0.01), whereas the immunopositivity to Claudin 1 in the treatment group significantly decreased as compared with that of LPS group (P < 0.01) (Figure 2). -
There was reduced mRNA expression in peroxisome proliferator-activated receptor alpha (PPARα), SGLT1 and microtubule associated protein 1 light chain 3 beta (LC3B) when compared the PBS group with the control group, respectively (P < 0.05 or P < 0.001). Other autophagy-related genes had no significant changes as compared the PBS group with the control group (Supplementary Figure S2A–I, available in www.besjournal.com).
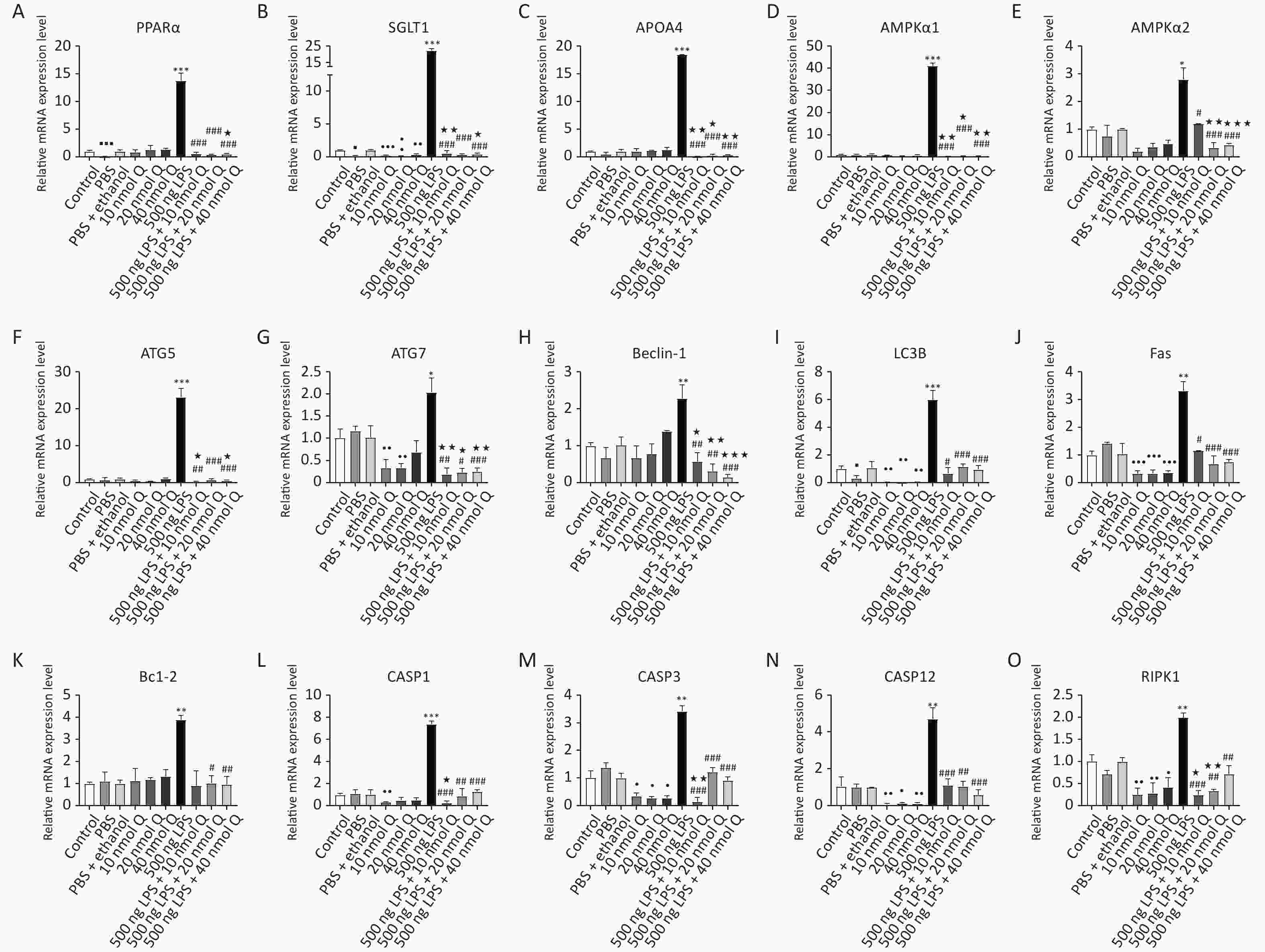
Figure S2. Qercetin ameliorates LPS-induced cardiac autophagy and programmed cell death in chicken embryos. The abbreviation and annotation is the same as Table 1 and Figure 2 Data are presented as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; #Indicates the significant differences between the LPS group and the (LPS + Q) group; ★Indicates the significant differences between the (PBS + ethanol) group and the (LPS + Q) group; ●Indicates the significant differences between the (PBS + ethanol) group and the Q (quercetin) group.
Quercetin significantly downregulated the mRNA expression of SGLT1 and LC3B when compared with the (PBS + ethanol) group at three dosages (P < 0.05, P < 0.01, or P < 0.001). Quercetin with 10 nmol or 20 nmol significantly downregulated the mRNA expression of ATG7 when compared with that of the (PBS + ethanol) group (P < 0.01).
Autophagy signaling pathway was significantly activated by LPS, as indicated by markedly increased mRNA expression of energy-deficient associated genes including PPARα, SGLT1, APOA4, AMPKα1, and AMPKα2 by 13.8 folds, 22.5 folds, 18.5 folds, 41.1 folds, and 2.8 folds. Autophagy-related genes (ATG) including ATG5, ATG7, Beclin-1, and LC3B mRNA expression were upregulated by 23.2 folds, 2.0 folds, 2.3 folds, and 6.0 folds, respectively (P < 0.05, P < 0.01, or P < 0.001). Quercetin downregulated these gene expressions at three doses (P < 0.05, P < 0.01, or P < 0.001) (10–40 nmol/egg).
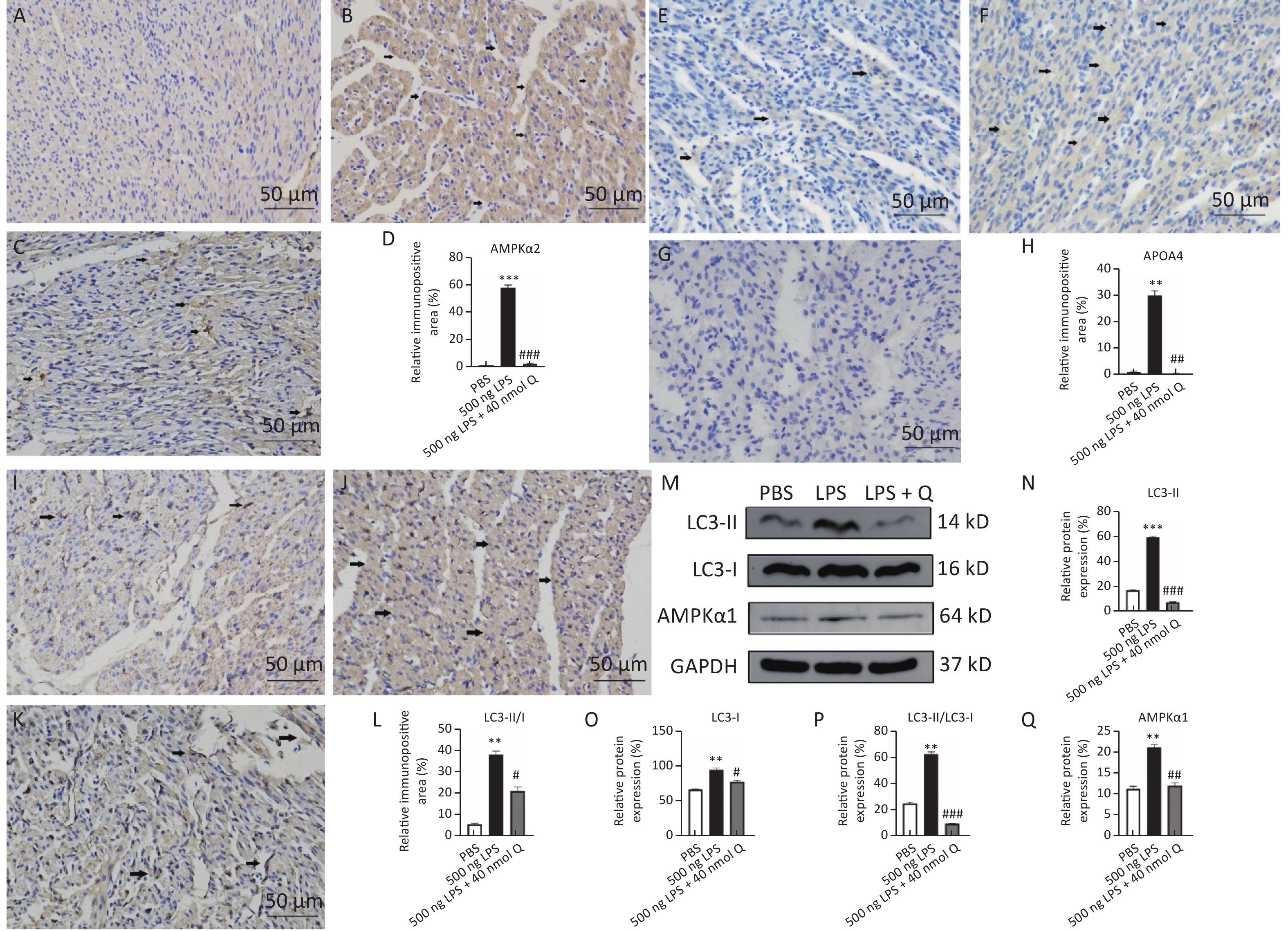
The immunopositivity of APOA4, AMPKα2, LC3-I, and LC3-II in the cytoplasm of myocardiocytes significantly increased after LPS induction when compared with that of the PBS group (P < 0.01 or P < 0.001), whereas the immunopositivity to APOA4, AMPKα2, LC3-I, and LC3-II in the treatment group significantly decreased when compared with that of the LPS group (P < 0.05 or P < 0.001) (Figure 3). The protein expressions of AMPKα1, LC3-I, LC3-II, and ratio of LC3-II/LC3-I significantly increased after LPS induction as compared with that of the PBS group (P < 0.01 or P < 0.001), whereas quercetin decreased these expressions when compared with the LPS group (P < 0.05, P < 0.01, or P < 0.001) (Figure 3). These findings clearly demonstrated that quercetin ameliorated LPS-induced cardiac inflammation through modulating autophagy.
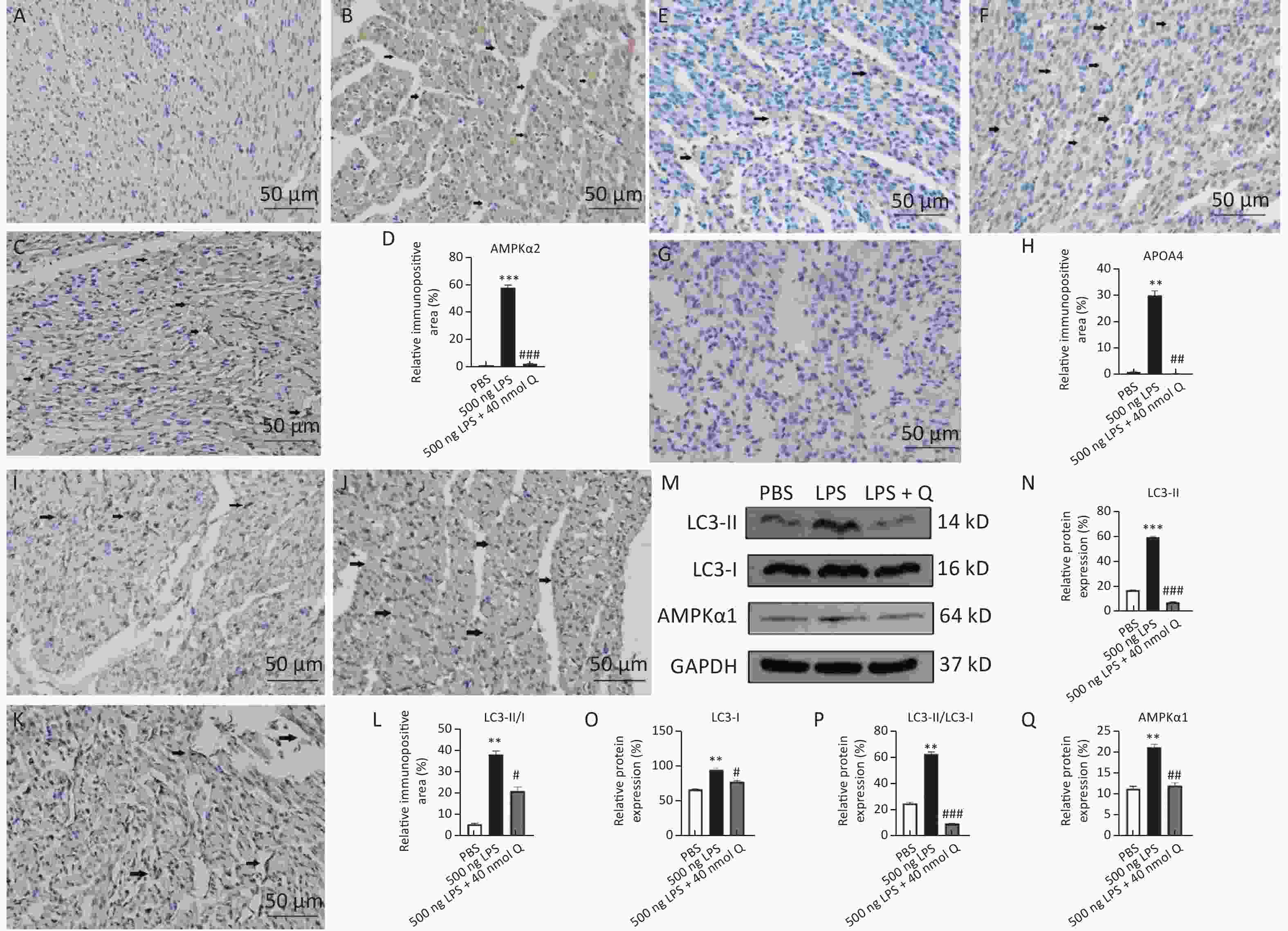
Figure 3. Quercetin alleviates the protein expression of AMPKα1, AMPKα2, APOA4, LC3-II, and LC3-I induced by LPS in the hearts of chicken embryos by immunohistochemistry and Western blotting. A, E, I: PBS group of AMPKα2, APOA4, and LC3-II/I, respectively; B, F, J: LPS group (500 ng LPS/egg) of AMPKα2, APOA4, and LC3-II/I, respectively; C, G, K, treatment group (500 ng LPS/egg + 40 nmol quercetin/egg) of AMPKα2, APOA4, and LC3-II/I, respectively; D, H, L, relative immunopositive area of AMPKα2, APOA4, and LC3-II/I, respectively. Immunopositivity to AMPKα2, APOA4, and LC3-II/I (arrow, brown to yellow). Scale bar: 50 μm. M, the protein expression of AMPKα1, LC3-I, and LC3-II; N, relative protein expression of LC3-II; O, relative protein expression of LC3-I; P, ratio of relative protein expression of LC3-II/LC3-I; Q, relative protein expression of AMPKα1. Data were expressed as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; **P < 0.01; ***P < 0.001. #Indicates the significant differences between the LPS group and the (LPS + Q) group; Q, Quercetin; #P < 0.05; ##P < 0.01; ###P < 0.001.
-
There was no different mRNA expression in programmed cell death-associated genes when compared the PBS group or the (PBS + ethanol) group with control group, respectively. Quercetin significantly downregulated the mRNA expression of Fas, CASP12, CASP3, RIPK1 (Supplementary Figure S2J–O), and dynamin 1 like (Drp1, Supplementary Figure S1N) when compared with the (PBS + ethanol) group at three dosages (P < 0.05, P < 0.01, or P < 0.001). Quercetin with 10 nmol significantly downregulated the mRNA expression of CASP1 when compared with that of the (PBS + ethanol) group (P < 0.01).
The mRNA expression of cardiac death receptors and ligands (TNFα and Fas), pyroptosis-associated gene caspases (CASP1 and CASP12), apoptosis-associated gene (CASP3 and Bcl-2), and necroptosis-associated gene (RIPK1) were upregulated by 19.9 folds, 3.3 folds, 7.4 folds, 4.7 folds, 3.4 folds, and 2.0 folds after LPS induction (P < 0.05, P < 0.01, or P < 0.001), respectively. Quercetin treatment significantly prevented programmed cell death factors increases after LPS induction. The mRNA expression of mitochondria fission factor Drp1 were downregulated by 0.6 folds after LPS induction (P < 0.05), while quercetin could upregulate the mRNA expression of Drp1 without a statistical difference.
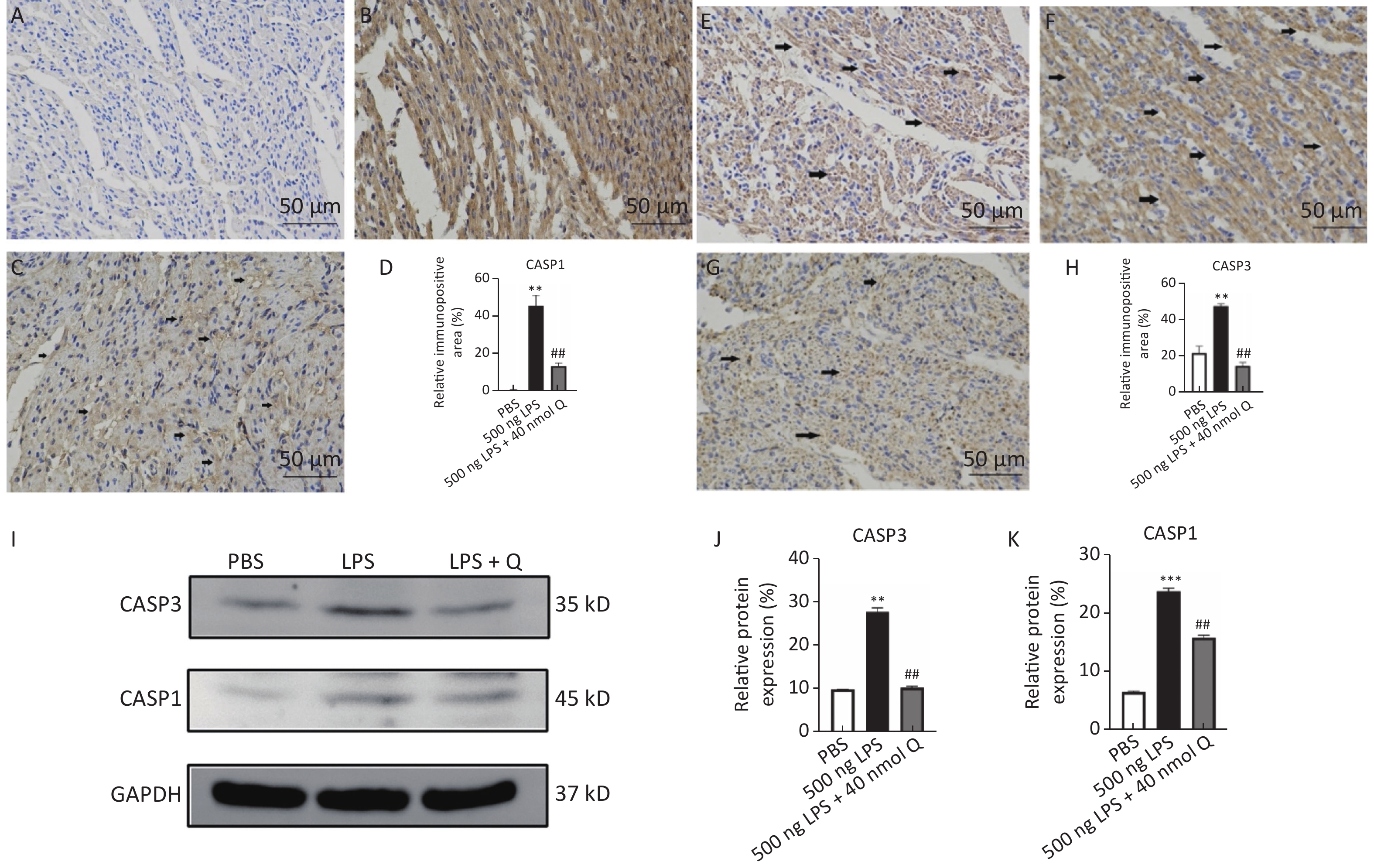
The immunopositivity of CASP1 and CASP3 in the cytoplasm of myocardiocytes was significantly enhanced after LPS induction when compared with that of the PBS group (P < 0.01), whereas quercetin significantly reduced the immunopositivity to CASP1 and CASP3 in the treatment group when compared with that of the LPS group (Figure 4A–H). The protein expressions of CASP1 and CASP3 significantly increased after LPS induction as compared with that of the PBS group (P < 0.01 or P < 0.001), whereas quercetin could significantly decrease these expressions when compared with that of the LPS group (P < 0.01) (Figure 4I–J). Our evidence demonstrated that quercetin ameliorated LPS-induced cardiac inflammation by mean of the modulation of programmed cell death.
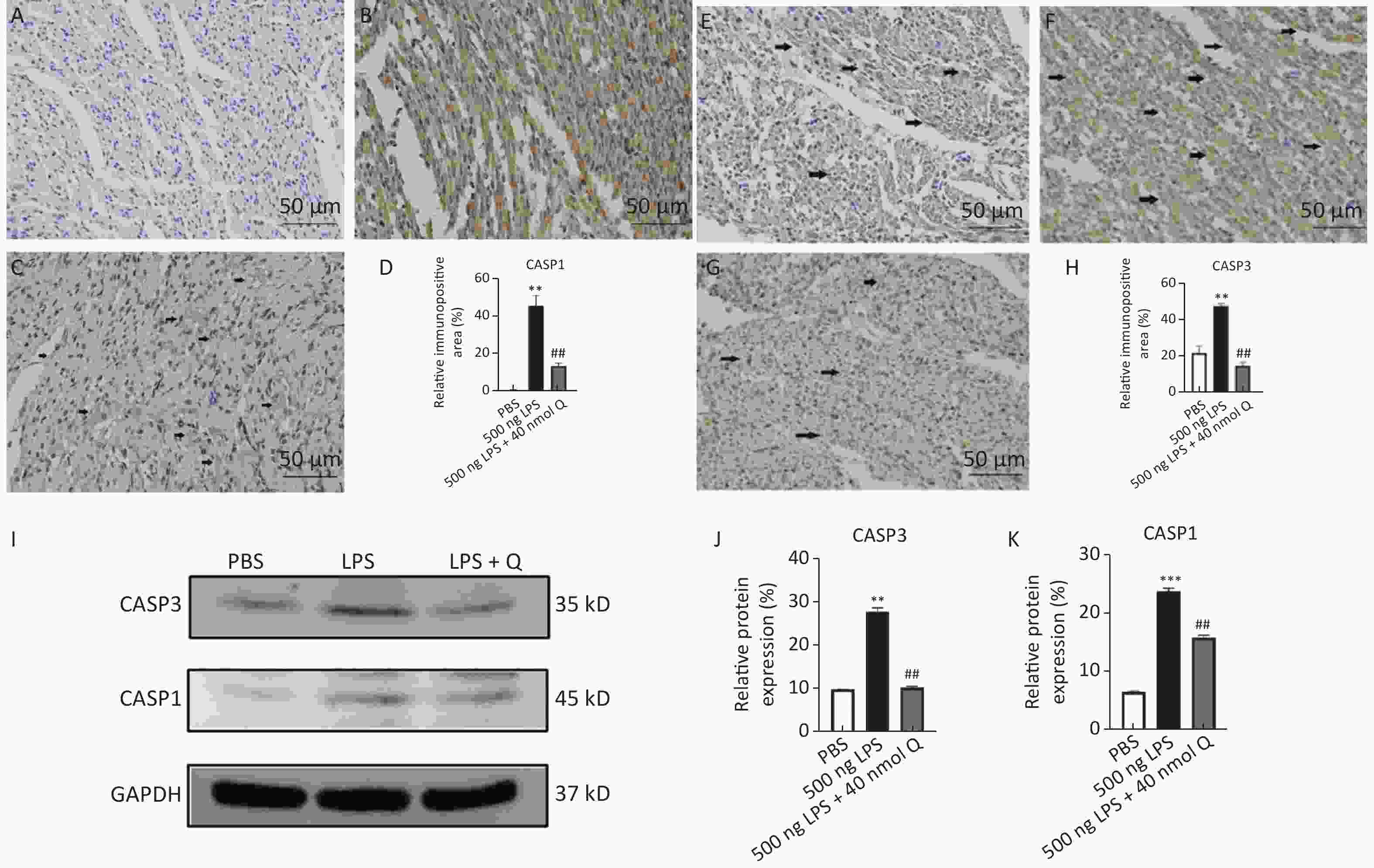
Figure 4. Quercetin alleviates the protein expression of CASP1 and CASP3 induced by LPS in the hearts of chicken embryos by immunohistochemistry and Western blotting. A, E: PBS group of CASP1 and CASP3, respectively; B, F: LPS group (500 ng LPS/egg) of CASP1 and CASP3, respectively; C, G, treatment group (500 ng LPS/egg + 40 nmol quercetin/egg) of CASP1 and CASP3, respectively; D, H, relative immunopositive area of CASP1 and CASP3, respectively. Immunopositivity to CASP1 and CASP3 (arrow, brown to yellow). Scale bar: 50 μm. I, the protein expression of CASP1 and CASP3; J, relative protein expression of CASP1; K, relative protein expression of CASP3. Data were expressed as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; **P < 0.01; ***P < 0.001. #Indicates the significant differences between the LPS group and the (LPS + Q) group; Q, Quercetin; #P < 0.05; ##P < 0.01.
-
Quercetin is a common flavonoid in vegetables, fruits, and traditional Chinese herbal medicines, including Manuka honey[29] and strawberry[30]. It has antioxidant[31], anti-inflammatory[32], hepatoprotective[33], and anti-cancer activity with respect to cancers such as blood, prostate and lung cancer[34]. The present study showed quercetin could significantly downregulate the mRNA expression of TNFα, IL-6, IL-8, p38, and MMP9 when compared with the (PBS + ethanol) group at three dosages in the hearts of chicken embryos, while significantly upregulating the mRNA expression of IL-10. It suggested that the quercetin has anti-inflammatory effect. In addition, quercetin with 10 nmol or 20 nmol significantly downregulated the mRNA expression of SGLT1, LC3B and ATG7 when compared with that of the (PBS + ethanol) group, and it revealed that quercetin reduced the mRNA expression of autophagy in the hearts of chicken embryos. Autophagy plays an important role in both pro-survival and pro-death functions in cancers. The targeting autophagy has been under consideration for prostate cancer therapy. Autophagy can affect the response of prostate cancer cells to therapy including chemotherapy and radiotherapy, nanoparticle[35], and phytochemicals[36]. Quercetin significantly downregulated the mRNA expression of Fas, CASP12, CASP3, RIPK1, and Drp1 when compared with the (PBS + ethanol) group at three doses, and it suggested that the quercetin could inhibit the cardiac pyroptosis, apoptosis, and necroptosis in chicken embryos.
The heart presented myocardial-fiber rupture and inflammatory cell infiltration after LPS induction in this study; LPS induction recruited banded heterophils and segmented heterophils in the hearts of chicken embryos, similar to a previous study[37]. However, quercetin can improve the inflammatory response in the hearts of chicken embryos.
Quercetin alleviates LPS-induced heart inflammation by the TLR4/MYD88/NF-κB signaling pathway. TLR4 gene expression was high in the heart after LPS induction[38], similar to our results. TLR4 could sense and recognize LPS, an evolutionally conserved process. Smooth LPS needs CD14 to activate the TLR4/MYD88/NF-κB signaling pathway[39], and the protein expression of TLR4 was associated with CD14 after LPS induction in human primary intestinal epithelial cells. Chicken heterophils play an important pole in innate immune response initiation and TLR4 recognition after Salmonella infection or smooth LPS induction from Salmonella spp.[40]; it stimulated an increased degranulation. There is CD14 in the heart of chicken embryo[41]; therefore, the conjugation of LPS with LPS-binding protein activated CD14 and initiated the TLR4/MYD88/NF-κB signaling pathway. The signal transduction in MyD88-dependent pathway proceeds by Toll/IL-1R domain-containing adaptor protein (TIRAP) recruitment to the Toll/IL-1R domain (TIR)-TIR part of TLR4 dimers, and the binding occurs via TIRAP-TIR-domain. Upon binding to TIRAP-TLR4 in vivo, MyD88 oligomerizes further and recruits the interleukin-1 receptor-associated serine/threonine kinases (IRAK) via the death domains (DD), forming myddosome[42]. TLR4 binds to certain cytokine receptors MYD88 to complete the pro-inflammatory cytokine signaling cascade. The protein and mRNA expression of TLR4 increased, and promoted the mRNA expression of MYD88, stimulated the inflammatory factors increase (IFNγ, IL-1β, IL-8, IL-6, IL-10, p38), and led to activation of the mRNA expression NF-κB1. TLR4 deletion mice increases basal energy expenditure and attenuates heart apoptosis[43]. Peripheral monocytes collected from different breeds of chicken showed that different genes expression of TLR4 and MYD88 when stimulated with LPS derived from S. Typhimurium[44]. MYD88 is widely expressed in a variety of cells in the cardiovascular system. Stressed cells and senescent cells released damage-associated molecular patterns (DAMP); Heterophils, monocytes, and macrophages first recognized DAMP through TLR4 to rapidly initiate immune responses and produce pro-inflammatory factors. One study showed that left ventricular function decreased in mice stimulated with LPS from E. coli, while left ventricular function did not decrease in TLR4-knockout mice[43]. This suggests that the TLR4 gene plays an important role in the heart after LPS stimulation. One study showed that protein expressions of TLR4 and MYD88 in the heart were upregulated after LPS induction in vivo and in vitro[45]. Our study found that mRNA and protein expressions of cardiac TLR4 were upregulated after LPS induction, whereas quercetin blocked the TLR4 expressions with three doses.
The infections from Gram-negative bacteria caused endotoxemia, resulting in multiple organ failure, including heart failure and kidney failure. LPS recruits chicken heterophils to the heart, initiating an inflammatory response. One study found that IL-6 deficient mice exhibited impaired heart rhythm and increased cardiac oxidative stress after LPS stimulation[46]. The mRNA expressions of IL-8 increased five-fold in chicken heterophils after Salmonella enterica subspecies enterica serovar Enteritidis infection[47], similar to our results. A previous study indicated that the expressions of IL-6 and TNFα were upregulated after LPS induction in rat primary myocardial fibroblasts (H9C2)[48], consistent with our results. IL-1β and TNFα are crucial proinflammatory cytokines, as they have powerful pro-inflammatory activities and promote the secretion of a variety of pro-inflammatory mediators. Our study found that the mRNA expression of cardiac inflammatory factors (IL-1β, IL-6, IL-8, and NF-κB1); meanwhile, mRNA and protein expressions of IFNγ were significantly upregulated after LPS induction in chicken embryos, while quercetin alleviated cardiac inflammatory responses and reduced the gene expressions of LPS-induced inflammatory factors. In addition, it was found that quercetin significantly downregulated the mRNA expression of TLR4, TNFα, IL-6, IL-8, p38, MMP3, and MMP9 when compared with the (PBS + ethanol) group. However, quercetin upregulated the mRNA expression of IL-10 (anti-inflammatory cytokine). It indicated that the quercetin could balance the mRNA expression of inflammation-associated factors.
Matrix metalloproteinases (MMPs), a kind of proteolytic enzymes of extracellular matrix, are a zinc-dependent endopeptidase family mainly responsible for tissue remodeling and regeneration in variable physiological and pathological processes. MMPs could degrade extracellular matrix proteins, glycoproteins, membrane receptors, cytokines, and growth factors. MMP3 was involved in initiating an early and lethal cytokine response to S. Typhimurium infection to mice[49]. The remodeling of the extracellular matrix is closely correlated with cell proliferation, differentiation, adhesion, migration, and apoptosis. After intraperitoneal injection of LPS in rats, plasma MMP2 levels and MMP9 levels were upregulated[50]. One study indicated that rutin, a quercetin glycoside[51], mitigated the expressions of fibrosis-related genes (MMP2 and MMP9) in the hearts to prevent LPS-induced cardiac fibrosis[52]. Cellular fibrosis (upregulation of MMP3 and MMP9) was induced by angiotensin II in H9C2 cells, and CASP3 significantly increased. MMP3-null myocardial fibroblast cell significantly decreased the protein expressions of MMP3, MMP9, and Bcl-2[53]. We found that the mRNA and protein expressions of MMP3 and MMP9 in the hearts increased after LPS induction in chicken embryos, it suggested that extracellular matrix degradation initiated after LPS induction in the hearts; meanwhile, quercetin can alleviate the mRNA and protein expression of MMP3 and MMP9 after LPS induction.
Quercetin alleviates LPS-induced mRNA and protein expression of Claudin 1 in the heart of chicken embryos. In vertebrates, there are more than 20 members of the Claudin family responsible for the permeability of tight junctions, divided into different groups based on their molecular weight and ion selectivity[54]. One study indicated that Claudin 1 was expressed in 3- to 8-days chick embryos[55]. The chick Claudin-1 clone shared 73% and 70% identity with the human and mouse Claudin-1 homologues, respectively. At HH (Hamburger and Hamilton) stage 4 and stage 10 embryos, Claudin-1 protein was primarily expressed in the surface of ectoderm, while ZO-1 and Occludin were more broadly expressed throughout the section at HH stage 5[56].
We found that the higher expression of Claudin 1 gene in myocardium of chicken embryos after LPS induction, reduced the permeability of myocardial cells. However, quercetin reduced the gene and protein expressions and restored it to normal. Tight junction gene expression is regulated at both transcriptional and post-transcriptional levels. ZO-1, an intracellular scaffolding protein, is located in the intercalated disks of the human heart[57], and regulated signal transduction, transcription and cellular communication in the heart[58]. Myocardiocytes expressions of ZO-1 are essential for normal atrioventricular conduction; global deletion of ZO-1 in the mice is lethal at embryonic day 11.5[59]. We found quercetin inhibited the ZO-1 expression induced by LPS in the hearts of chicken embryos.
The heart is an organ with high energy demand, which makes it particularly sensitive to mitochondrial dysfunction. Mitochondria are ubiquitous organelles forming large tubular structures that spread throughout the cytoplasm, and are the principal producers of cellular ATP. They constitute 30% of myocardial mass and are associated with various cellular activities critical to homeostasis, including mitochondrial biosynthesis, dynamic fission and fusion, and mitophagy. Mitochondrial fusion and fission are evolutionarily conserved mechanisms that promote mitochondrial health via the exchange of mitochondrial proteins, lipids, and genomes. The mitochondrial fusion and fission cycle in adult mice hearts occurs approximately every 16 days[60]. The key component of the mitochondrial fission process is segregation and further asymmetric separation of a mitochondrial body, yielding healthy and impaired organelles with subsequent decomposition and removal of impaired elements through autophagy. Mitochondrial stress in myocardiocytes which induced by LPS manifested that superoxide overproduction, ATP deprivation, and apoptosis activation[61]. A previous study found that oxidative stress increased and mitochondrial dysfunction occured in fibroblasts after LPS from Porphyromonas gingivalis induction, it decreased the mitochondrial protein expression, mitochondrial mass, and mitochondrial membrane potential[62]. Another study indicated that the LPS of Porphyromonas gingivalis infection induced mitochondrial fragmentation, increased the mitochondrial reactive oxygen species levels, and decreased the mitochondrial membrane potential and ATP concentration in vascular endothelial cell[63]. The dynamin-related protein Drp1, a GTPase, is a major regulator of mitochondrial fission; it can activate apoptosis by promoting the opening of the mitochondrial permeability transition pore complex, and change the mitochondrial outer membrane permeability, and promote release of intermembrane proteins[64]. Drp1 deficiency in myocardiocytes caused energy shortage and heart failure, and defects of mitochondrial morphology, ultrastructure, and function of the right ventricle in the embryonic hearts of mice[65]. Another study indicated that Drp1 inhibition was beneficial to myocardial functions[66], because it preserved the mitochondrial networking and morphology. Downregulation of Drp1 gene expressions can induce mitochondrial elongation, and damaged mitochondria will accumulate and initiate apoptosis[67]. We found that the cardiac gene expression of Drp1 decreased after LPS induction and balanced the mitochondria dynamic fusion and fission, led to the increases of mitochondria dysfunction, and triggered the gene expressions of autophagy (ATG5 and LC3B).
Pathophysiology of cardiometabolic syndrome included hypertension with endothelial dysfunction, dysregulated lipid metabolism, oxidative and nitrosative stress, endoplasmic reticulum stress, mitochondrial damage, and inflammation. Certain cellular machineries such as autophagy, apoptosis, pyroptosis, and necroptosis were the main regulatory mechanisms for cell death and survival of cardiometabolic syndrome. Protein quality control is mainly governed by the ubiquitin proteasome system and autophagy-lysosomal pathways to degrade and remove unfold and damaged proteins. Inflammation can cause mitochondrial injures, leading to abnormal accumulation of aged or injured cell debris and organelles. Mitochondrial injures resulted in reduced energy output and a deficient cellular nutrients status. Autophagy, a conserved intracellular pathways, is an intracellular catabolic process which enables the cell to degraded unwanted cellular components within a double membrane structure formed upon fusion with the lysosome[68]. Therefore, autophagy plays a critical role in normal cellular homeostasis and development.
The heart is a critical organ with high energy demand, and mitochondrial injures occurred after LPS induction with lower ATP, ADP, and AMP levels[69]. LPS induced oxidative damage in cytosol and mitochondria[70]. SGLT1 is a sodium glucose co-transporter 1 and broadly distributed in the heart, the kidney, and the small intestine. SGLT1 is expressed in myocardial fibers. The mRNA expression of SGLT1 increased after LPS induction in the hearts of chicken embryos; it suggested that the energy deficiency promoting the glucose transporter-associated gene expression increase. However, the quercetin could suppressed the mRNA expression of SGLT1; it may result from enhancement of oxidative stress in cytosol and mitochondria. Evidence shows quercetin prevents LPS-induced oxidative stress and inflammation by modulating the NADPH oxidase 2 (NOX2)/ reactive oxygen species (ROS)/NF-kB pathway in lung epithelial cells[71].
Autophagy plays a crucial pole in lipid metabolism and degradation in mycocardiocytes and serves an important mechanism for cholesterol efflux to APOA4. APOA4, an essential gene in both glucose and lipid metabolism, is the main circulating apolipoprotein. APOA4 contributes to absorption of edible fat, triglyceride transport, and reverse cholesterol transport. The mRNA expression of APOA4 increased in rat myocardium during hypercholesterolemia model[72]. APOA4-knockout mice showed that elevated triglyceride content, enhanced expression of lipogenic enzymes, and diminished phosphorylated protein kinase (Akt) in the myocardium by inhibiting the phosphoinositide 3-kinase (PI3K-Akt) pathway in stable obese animal models[73]. A previous study indicated the quercetin postconditioning could induce cardioprotection by activating the PI3K/Akt signaling pathway in the myocardial ischemia/reperfusion injuries in rat[74]. In the present study, the cardiac mRNA and protein expressions of APOA4 increased after LPS induction; it suggested APOA4 increased fat catabolism after LPS induction. Nevertheless, quercetin decreased the expressions of APOA4; it might activate the PI3K-Akt pathway.
Autophagy can recycle and regenerate the cell components such as amino acids, lipids, and other metabolic precursors. Autophagosomes fuse with lysosomes to generate autophagolysosomes, where the cargo contents are digested to release new amino acids and free fatty acids. Because the fatty acids can produce much more ATP (the ratio of a single molecule of palmitate to glucose producing ATP is nearly 104:31). Therefore, it promoted lipid peoxidation in energy insufficiency of myocardiocytes after LPS induction. LPS-induced sepsis or endotoxemia caused energy deficiency by inhibiting the oxidation of long-chain fatty acids and glucose in the myocardium. Evidence indicates that the total fatty acid level and the non-esterified fatty acid level of the heart decreased after LPS induction[75]. The level of short- and medium-chain acylcarnitines, lactate, and alanine in myocardium increased after LPS induction in pigs, and decreased the oxidation of short and medium-chain fatty acid[76]. LPS can trigger cardiac energy deficiency with hypoglycemia and hyperlactacidemia and increase AMPK mRNA expression, as well as promote the increase of ATG5, ATG7, and LC3B. Septic shock causes hypoglycemia and oxidation of long-chain fatty acids, resulting in myocardial energy deficiency. When the heart is induced by LPS, the ATP level of the heart decreases, inducing autophagy to meet the energy needs and protein synthesis[77].
Autophagy is tightly regulated by posttranscriptional signaling mechanisms including AMPK and the anti-autophagy protein (Bcl-2), and transcriptional regulation by factors such as PPARα in the hearts. Fatty acid oxidation is transcriptionally regulated by PPARα in the hearts and accounts for 70% of cardiac ATP content in normal conditions. Reduced PPARα expression leads to fatty acid oxidation decrease and myocardial energy deficiency in mitochondria during sepsis and heart failure [78]. PPARα null mice indicated that the autophagy enhancement (the protein expressions of Atg5, Atg7, Beclin-1, and LC3B were upregulated)[79]. It was found that the mRNA expressions of PPARα and SGLT1 upregulated after LPS-induced energy deficiency, and the mRNA and protein expressions of APOA4 increased; whereas quercetin could balance gene and protein expressions to improve the energy status in mitochondria.
AMPK is a conserved cell energy sense gene that maintains energy balance, cells growth, protein synthesis, apoptosis, and autophagy. Multiple members of mechanistic target of rapamycin kinase (MTOR) pathways can regulate the initiation of autophagy, including AMPK and MTOR complex 1(MTORC1). AMPK is an upstream regulator of the MTOR. When energy deficiency, nutrient deprivation, and inflammation occur, the energy sensitive AMPKα1 and AMPKα2 can be activated; While the MTORC1 is activated, autophagy is inhibited in nutrient rich conditions[80]. In the heart, AMPK is activated in ischemia, hypoglycemia, and energy deficiency; glucose absorption and glycolysis increased. Apoptosis and autophagy are very complex processes in vivo, and the expressions of apoptosis and autophagy are regulated by reciprocal feedback[81]. Our findings showed that cardiac energy deficiency after LPS induction activated AMPKα1 and AMPKα2 mRNA and protein expressions, and promoted autophagy and pyroptosis. Bcl-2 is both an anti-apoptotic gene and a pro-apoptotic gene. One study indicated that the gene expression of Bcl-2 was very high in the hearts of fetal rats, and decreased within one to five days postpartum, and then gradually increased at 11–21 days old[82]. Another study found that the ovaries and testes of chicken embryos expressed Bcl-2 and CASP3 proteins from embryonic day 12[83]. In the present study, the expression of anti-apoptotic gene Bcl-2 was upregulated after LPS induction, while quercetin blocked upregulation of apoptosis (the mRNA and protein expressions of CASP3 decreased).
Beclin-1, a marker and regulator of autophagy, plays a central role in the initiation of autophagy that ubiquitously expressed. The phosphorylation of Beclin-1 is post-translationally modified by AMPK and ULK. Beclin-1 is taken part in regulating PI3K complexes activities. LPS decreased both the total mass and the function of mitochondria-associated membranes, and these conditions became worse in Beclin-1 knockout mice[84]. Cardiac-specific overexpression of Beclin-1 promoted autophagy; suppressed mammalian target of rapamycin signaling; improved cardiac function; and alleviated inflammation and fibrosis after LPS induction[85]. Inhibition of autophagy with genetic deletion of Beclin-1 or Atg7 also suppressed apoptosis, denoting a close association between autophagy and apoptosis[86]. In this report, the mRNA expressions of Beclin-1 upregulated after LPS induction in the hearts of chicken embryos, and initiated the autophagy, while quercetin could significantly downregulated the Beclin-1 expression.
Autophagy is a survival mechanism that initiates cell apoptosis. Autophagy refers to the process involving the decomposition of intracellular components via lysosomes. ATG5, LC3-I, and LC3-II participate in the formation of autophagosome. There were two types of ubiquitin-like binding reaction systems, namely ATG5-ATG7-ATG12 binding reaction systems and LC3-I/LC3-II binding reaction systems. Deletion of ATG genes (ATG3, ATG5, ATG6, ATG7, ATG9, and ATG16L) resulted in the death of all developing mice[87]. ATG5, ATG16, and ATG12 form a complex that catalyzes LC3-I to LC3-II on the phagocytic membrane. LC3-I, a ubiquitin-like protein, exists in the cytoplasm, and conjugates to phosphatidylinositol to form LC3-II. LC3-II is the only protein marker of autophagosome maturation. TNFα and TNFα receptors are expressed in all cells of the body. The death receptor Fas and ligand TNFα of the heart were upregulated after LPS induction in the present study. In one study, immortal chicken embryo fibroblast DF-1 cells with ATG5-knockout inhibited cell proliferation and promoted apoptosis (the mRNA expression of CASP3 increased)[88]. ATG5-null mice caused cardiomyopathy, mitochondrial collapse; increased left ventricular volume; decreased cardiac function; and promoted animals death[89]. Our results showed that the gene and protein expressions of cardiac autophagy (LC3-I and LC3-II) was upregulated after LPS induction. Quercetin significantly inhibited the expressions in the chicken hearts after LPS induction. These results suggested that quercetin improved LPS-induced energy deficiency and reduced the activation of cardiac autophagy pathway.
Pyroptosis is an inflammatory programmed cell death with CASP1-dependent and CASP11-dependent pathways. CASP1 and CASP12 are inflammatory caspases, which are only found in vertebrate. CASP1 can activate and proteolytic cleave pro-IL-1β and pro-IL-18 to IL-1β and IL-18. IL-18 are IFNγ-inducing cytokines. Endogenous CASP1-cleaved IL-18 significantly contributes to induction of IFN-gamma by IL-12. CASP1 and CASP12 are critical components for the prognosis of cardiovascular diseases and inflammasome[90]. CASP12 is ubiquitously expressed and localized in the endoplasmic reticulum (ER) and activated by ER stress, including disruption of ER calcium homeostasis and accumulation of excess proteins in ER, leading to apoptosis[91]. Caspase-12-deficient mice are partially resistant to apoptosis induced by ER stress but sensitive to other apoptotic stimulation[92]. Caspase-12 mRNA expression is upregulated to a similar extent in wildtype and caspases-1/11 deficient cells upon treatment with LPS in vitro[93]. Inflammasome is closely associated with pyroptosis. Inflammasome initiates when LPS activates TLR4 and NOD-like receptors such as NLRP3. TLR4 activate NF-κB1 and MYD88 expressions and induce NLRP3 and pro-IL-1β release. A previous study indicated that Glaesserella parasuis LPS-mediated inflammation leads to pyroptosis in porcine alveolar macrophage[94], similar to our results. In the present study, the mRNA and protein expressions of TLR4 and CASP1 increased after LPS induction; meanwhile, the mRNA expressions of CASP12, NF-κB1, and MYD88 were upregulated after LPS induction in the hearts of chicken embryos. It indicted that the CASP1 dependent pyroptosis pathway occurred after LPS induction in the hearts of chicken embryos; however, quercetin could ameliorate the pyroptosis in the hearts of chicken embryos.
RIPK1 was initially identified through a yeast two-hybrid screening as a Fas-interacting protein and an adaptor protein in the TNF receptor 1 (TNFR1) signaling complex. There is a serine/threonine kinase domain and a death domain which can bind to the Fas, TNFR1, Tumor necrosis factor-related apoptosis-inducing ligand receptor 1 (TRAILR1), and TRAILR2 in the RIPK1. RIPK1 is a critical adaptor protein for TNFR1-mediated signaling to NF-κB activation. The conjugation of RIPK1 with the Fas or TNFR1 activated mitogen activated protein kinases (MAPK, including JNK, extracellular signal-regulated kinase ERK, and p38), apoptosis and necroptosis signaling pathways. LPS was recognized by TLR4 and activated the NF-κB1 and PI3K/AKT, apoptosis and necroptosis pathways[95]. One study indicated that gallic acid attenuated LPS-induced CASP3 and RIPK1 activation[96], similar to our finding. In the present study, the mRNA expressions of RIPK1, TLR4, NF-κB1, Fas, TNFα, and p38 upregulated after LPS induction; whereas quercein inhibited the abovementioned signaling pathways. Quercetin could alleviate the mRNA and protein expressions of CASP3, and mRNA expression of RIPK1.
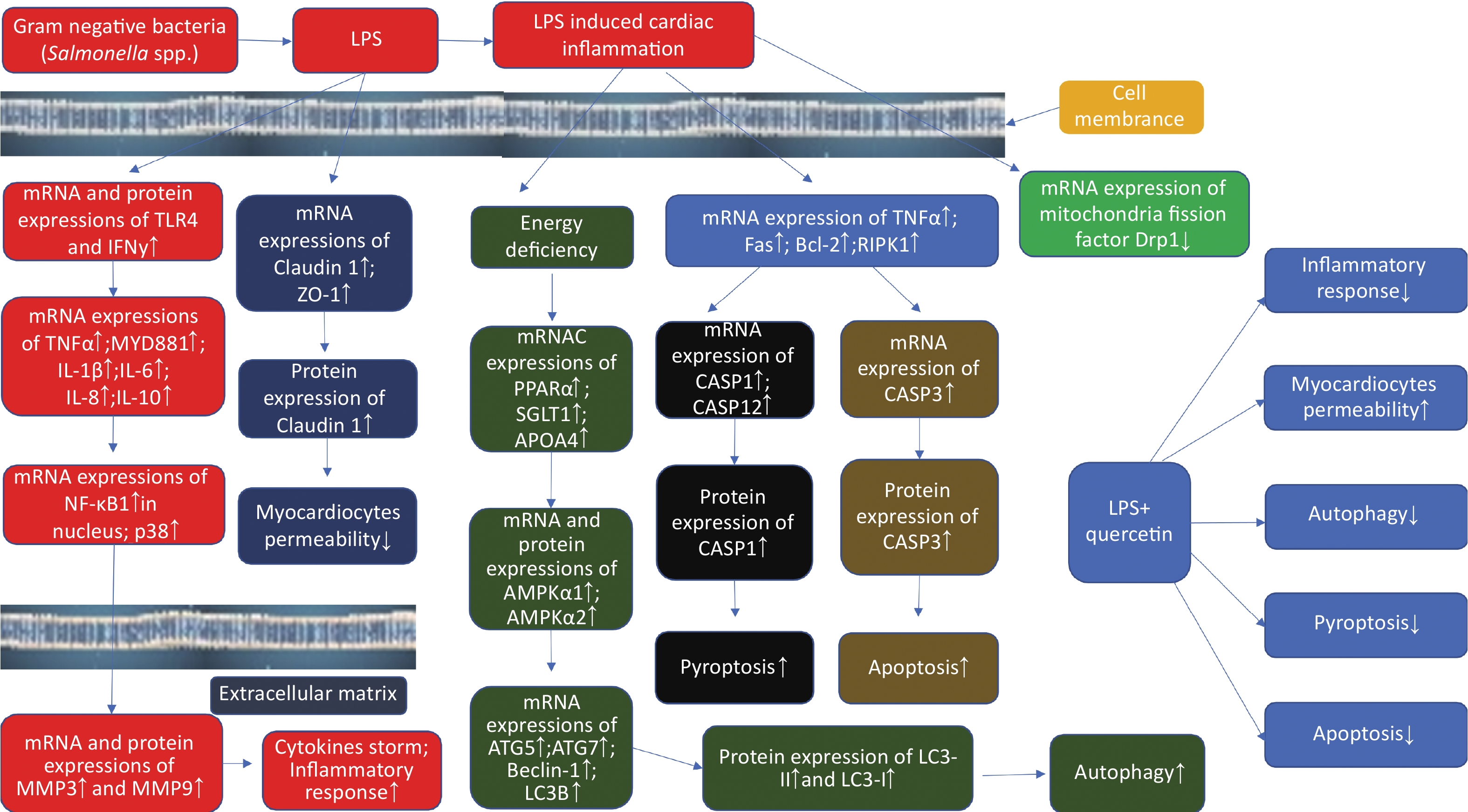
In conclusion, LPS induced cardiac inflammation in the present study; caused cardiac inflammatory cell infiltration; and increased autophagy, apoptosis, and pyroptosis; upregulated myocardiocytes permeability; however, quercetin alleviated cardiac inflammation induced by LPS by modulating myocardiocytes permeability, autophagy, and programmed cell death (Supplementary Figure S3, available in www.besjournal.com).
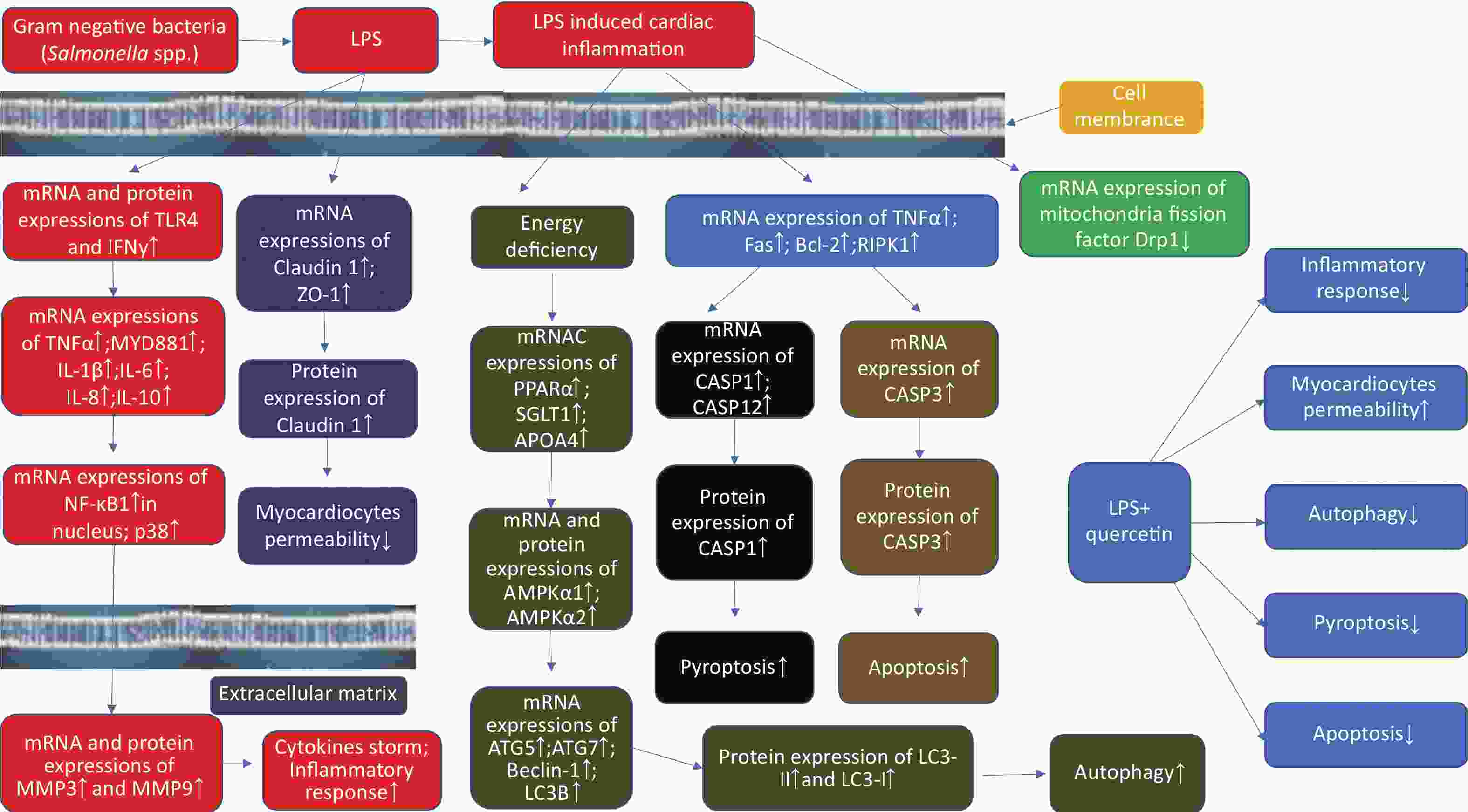
Figure S3. The schematic figure on quercetin attenuate cardiac inflammation, autophagy, pyroptosis, and apoptosis after LPS induction in chicken embryos.
-
The study was approved by the University Animal Ethics Committee (JXAULL-2022002).
doi: 10.3967/bes2024.006
Quercetin Alleviates Lipopolysaccharide-Induced Cardiac Inflammation via Inhibiting Autophagy and Programmed Cell Death
-
Abstract:
Objective The aim of this study is to explore the potential modulatory role of quercetin against Endotoxin or lipopolysaccharide (LPS) induced septic cardiac dysfunction. Methods Specific pathogen-free chicken embryos (n = 120) were allocated untreated control, phosphate buffer solution (PBS) vehicle, PBS with ethanol vehicle, LPS (500 ng/egg), LPS with quercetin treatment (10, 20, or 40 nmol/egg, respectively), Quercetin groups (10, 20, or 40 nmol/egg). Fifteen-day-old embryonated eggs were inoculated with abovementioned solutions via the allantoic cavity. At embryonic day 19, the hearts of the embryos were collected for histopathological examination, RNA extraction, real-time polymerase chain reaction, immunohistochemical investigations, and Western blotting. Results They demonstrated that the heart presented inflammatory responses after LPS induction. The LPS-induced higher mRNA expressions of inflammation-related factors (TLR4, TNFα, MYD88, NF-κB1, IFNγ, IL-1β, IL-8, IL-6, IL-10, p38, MMP3, and MMP9) were blocked by quercetin with three dosages. Quercetin significantly decreased immunopositivity to TLR4 and MMP9 in the treatment group when compared with the LPS group. Quercetin significantly decreased protein expressions of TLR4, IFNγ, MMP3, and MMP9 when compared with the LPS group. Quercetin treatment prevented LPS-induced increase in the mRNA expression of Claudin 1 and ZO-1, and significantly decreased protein expression of claudin 1 when compared with the LPS group. Quercetin significantly downregulated autophagy-related gene expressions (PPARα, SGLT1, APOA4, AMPKα1, AMPKα2, ATG5, ATG7, Beclin-1, and LC3B) and programmed cell death (Fas, Bcl-2, CASP1, CASP12, CASP3, and RIPK1) after LPS induction. Quercetin significantly decreased immunopositivity to APOA4, AMPKα2, and LC3-II/LC3-I in the treatment group when compared with the LPS group. Quercetin significantly decreased protein expressions of AMPKα1, LC3-I, and LC3-II. Quercetin significantly decreased the protein expression to CASP1 and CASP3 by immunohistochemical investigation or Western blotting in treatment group when compared with LPS group. Conclusion Quercetin alleviates cardiac inflammation induced by LPS through modulating autophagy, programmed cell death, and myocardiocytes permeability. -
Key words:
- Quercetin /
- Lipopolysaccharide /
- Inflammation /
- Autophagy /
- Programmed cell death /
- Myocardiocytes permeability
The authors declare no conflict of interest.
注释:1) AUTHOR CONTRIBUTIONS: 2) CONFLICTS OF INTEREST: -
Figure 1. The histopathologic changes of the hearts induced by LPS and quercetin in chicken embryos A, control group; B, PBS group; C, PBS + ethanol group; D–F, 10 nmol, 20 nmol or 40 nmol quercetin; G, treatment group (500 ng/egg); H–J, 500 ng LPS +(10 nmol, 20 nmol, or 40 nmol). Scale bar: 40 μm. HB: Heterophilc banded cells, arrow.
S1. Quercetin alleviates the mRNA expressions of inflammatory associated factors induced by LPS in the hearts of chicken embryos. Data are presented as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; #Indicates the significant differences between the LPS group and the (LPS + Q) group; ★Indicates the significant differences between the (PBS + ethanol) group and the (LPS + Q) group; ●Indicates the significant differences between the (PBS + ethanol) group and the Q (quercetin) group.
Figure 2. Quercetin alleviates the protein expression of TLR4, MMP9, Claudin 1, IFNγ, and MMP3 induced by LPS in the hearts of chicken embryos by immunohistochemistry and Western blotting. A, E, I: PBS group of TLR4, MMP9, Claudin 1, respectively; B, F, J: LPS group (500 ng LPS/egg) of TLR4, MMP9, Claudin 1, respectively; C, G, K, treatment group (500 ng LPS/egg + 40 nmol quercetin/egg) of TLR4, MMP9, Claudin 1, respectively; D, H, L, relative immunopositive area of TLR4, MMP9, Claudin 1, respectively. Immunopositivity to TLR4, MMP9, Claudin 1 (arrow, brown to yellow). Scale bar: 50 μm. M, the protein expression of TLR4, IFNγ, MMP3 and MMP9; N, relative protein expression of MMP9; O, relative protein expression of MMP3; P, relative protein expression of IFNγ; Q, relative protein expression of TLR4. Data were expressed as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; **P < 0.01; ***P < 0.001. #indicates the significant differences between the LPS group and the (LPS + Q) group; Q, Quercetin; #P < 0.05; ##P < 0.01; ###P < 0.001.
S2. Qercetin ameliorates LPS-induced cardiac autophagy and programmed cell death in chicken embryos. The abbreviation and annotation is the same as Table 1 and Figure 2 Data are presented as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; #Indicates the significant differences between the LPS group and the (LPS + Q) group; ★Indicates the significant differences between the (PBS + ethanol) group and the (LPS + Q) group; ●Indicates the significant differences between the (PBS + ethanol) group and the Q (quercetin) group.
Figure 3. Quercetin alleviates the protein expression of AMPKα1, AMPKα2, APOA4, LC3-II, and LC3-I induced by LPS in the hearts of chicken embryos by immunohistochemistry and Western blotting. A, E, I: PBS group of AMPKα2, APOA4, and LC3-II/I, respectively; B, F, J: LPS group (500 ng LPS/egg) of AMPKα2, APOA4, and LC3-II/I, respectively; C, G, K, treatment group (500 ng LPS/egg + 40 nmol quercetin/egg) of AMPKα2, APOA4, and LC3-II/I, respectively; D, H, L, relative immunopositive area of AMPKα2, APOA4, and LC3-II/I, respectively. Immunopositivity to AMPKα2, APOA4, and LC3-II/I (arrow, brown to yellow). Scale bar: 50 μm. M, the protein expression of AMPKα1, LC3-I, and LC3-II; N, relative protein expression of LC3-II; O, relative protein expression of LC3-I; P, ratio of relative protein expression of LC3-II/LC3-I; Q, relative protein expression of AMPKα1. Data were expressed as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; **P < 0.01; ***P < 0.001. #Indicates the significant differences between the LPS group and the (LPS + Q) group; Q, Quercetin; #P < 0.05; ##P < 0.01; ###P < 0.001.
Figure 4. Quercetin alleviates the protein expression of CASP1 and CASP3 induced by LPS in the hearts of chicken embryos by immunohistochemistry and Western blotting. A, E: PBS group of CASP1 and CASP3, respectively; B, F: LPS group (500 ng LPS/egg) of CASP1 and CASP3, respectively; C, G, treatment group (500 ng LPS/egg + 40 nmol quercetin/egg) of CASP1 and CASP3, respectively; D, H, relative immunopositive area of CASP1 and CASP3, respectively. Immunopositivity to CASP1 and CASP3 (arrow, brown to yellow). Scale bar: 50 μm. I, the protein expression of CASP1 and CASP3; J, relative protein expression of CASP1; K, relative protein expression of CASP3. Data were expressed as the mean ± SD. *Indicates the significant differences between the PBS group and the LPS group; **P < 0.01; ***P < 0.001. #Indicates the significant differences between the LPS group and the (LPS + Q) group; Q, Quercetin; #P < 0.05; ##P < 0.01.
S3. The schematic figure on quercetin attenuate cardiac inflammation, autophagy, pyroptosis, and apoptosis after LPS induction in chicken embryos.
Table 1. Primer used in real-time quantitative polymerase chain reaction
Genes name Primer sequence (5’–3’) Gene bank ID APOA4 F:GAGGTACTTCACTGAGCTGGG R:TGAGCTGCTTGGTGATCTCAG NM_204938 AMPKα1 F:CGGCAGATAAACAGAAGCACGAG; R:CGATTCAGGATCTTCACTGCAAC NM_001039603 AMPKα2 F:CATGGACGTGTTGAAGAGGCAG; R:TTCTCTGGTTTCAGGTCCCTGTGG NM_001039605 ATG5 F:CCCATCCCTGGTCCGTAAC; R:CGGCGGCGTATACGAAGTA NM_001006409 ATG7 F:TGCAGTTTGCTCCCTTCAGT R:TGGGAAACCTGATGGATCGC NM_001396468 Bcl-2 F: TGGCTGCTTTACTCTTGGGG; R:TATCTCGCGGTTGTCGTAGC NM_205339 Beclin-1 F:CCGCTGAAGCTCGATACCTC R:TTCTGGCTGGTGGGATGAAC NM_001006332.1 CASP1 F:CACTTCCACTTCGGATGGCT; R:CCACGAGACAGTATCAGGCG XM_015295935 CASP3 F:ACCGAGATACCGGACTGTCA; R:GCCATGGCTTAGCAACACAC NM_204725 CASP12 F:AATAGTGGGCATCTGGGTCA; R:CGGTGTGATTTAGACCCGTAAGAC [97] Claudin 1 F:CTGGGTCTGGTTGGTGTGTT; R:CGAGCCACTCTGTTGCCATA NM_001013611 Fas F:GTCAGTGCTGCACGAAATGT; R:AACCTCCAAACCGAGTGCTT NM_001199487 Drp1 F: GGCAGTCACAGCAGCTAACA; R:GCATCCATGAGATCCAGCTT NM_001079722 GAPDH F: GAGAAACCAGCCAAGTATGATG; R: CACAGGAGACAACCTGGTCC NM_204305 IFNγ F:CTGACAAGTCAAAGCCGCAC; R:CTTCACGCCATCAGGAAGGT NM_205149 IL-1β F:GCTCAACATTGCGCTGTACC; R:AGGCGGTAGAAGATGAAGCG FJ537850 IL-6 F:ACGAGGAGAAATGCCTGACG; R:CTTCAGATTGGCGAGGAGGG NM_204628 IL-8 F:TGCCAGTGCATTAGCACTCA; R:TTGGCGTCAGCTTCACATCT HM179639 IL-10 F:TGCGAGAAGAGGAGCAAAGC R:AACTCCCCCATGGCTTTGTAG AJ621254 LC3B F:CTTCTTCCTCCTGGTGAACG; R:GCACTCCGAAAGTCTCCTGA NM_001031461 MMP9 F:ACACAGACTCTATGCTGCCTG; R:GAGAGTAGGGCGGGGAAAAT NM_204667 MMP3 F:ATCAGGCTCTACAGTGGTG; R:ATGGGATACATCAAGGCAC XM_025152201 MYD88 F:TTAGTCTTTCCCCAGGGGCT; R:GCCAGTCTTGTCCAGAACCA NM_001030962 NF-κB1 F:TCAACGCAGGACCTAAAGACAT; R:GCAGATAGCCAAGTTCAGGATG NM_001396396 Occludin F:TACATCATGGGCGTCAACCC R:CCAGATCTTACTGCGCGTCT NM_205128 PPARα F:AACGGAGTTCCAATCGCTGG R:GGATGCTGGTGAAAGGGTGT XM_025150258 RIPK1 F:GATCCATTTGCGAAGCTGCC R:CTTAGGCTAATGGCGCTGGT NM_204402 SGLT1 F:GGCCAGGGCTTACTATTGGT R:AGCTTCAGGTATCCGCACAT AJ236903 TLR4 F:GGCTCAACCTCACGTTGGTA; R:AGTCCGTTCTGAAATGCCGT KP410249 ZO-1 F:TATGAAGATCGTGCGCCTCC; R:GAGGTCTGCCATCGTAGCTC XM_015278977 TNFα F:CCCATCCCTGGTCCGTAAC; R:CGGCGGCGTATACGAAGTA MF000729 Note. APOA4: apolipoprotein 4; AMPKα1: Protein kinase AMP-activated catalytic subunit alpha 1; AMPKα2: Protein kinase AMP-activated catalytic subunit alpha 2; ATG5: Autophagy related gene 5; ATG7: Autophagy related gene 7; Bcl-2: B cell CLL/lymphoma 2; CASP1: Caspase 1; CASP3: Caspase 3; CASP12: Caspase 12; Drp1: dynamin 1 like; GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; Fas: Fas cell surface death receptor; IFNγ: Interferon gamma; IL-8: Interleukin-8; IL-1β: Interleukin-1β; IL-6: Interleukin-6; IL-10: Interleukin-10; LC3B: Microtubule associated protein 1 light chain 3 beta; MMP3: Matrix metallopeptidase 3; MMP9: Matrix metallopeptidase 9; MYD88: Myeloid differentiation primary response 88; NF-κB1: Nuclear factor kappa B subunit 1; RIPK1: receptor interacting serine/threonine kinase 1; PPARα: Peroxisome proliferator-activated receptor alpha; SGLT1: Sodium glucose co-transporter1; TLR4: Toll like receptor 4; ZO-1: Zonula occludens-1; TNFα: Tumor necrosis factor alpha.  下载: 导出CSV
下载: 导出CSV
-
[1] Schlapbach LJ, Kissoon N, Alhawsawi A, et al. World Sepsis Day: a global agenda to target a leading cause of morbidity and mortality. Am J Physiol Lung Cell Mol Physiol, 2020; 319, L518−22. doi: 10.1152/ajplung.00369.2020 [2] Balayan S, Chauhan N, Chandra R, et al. Recent advances in developing biosensing based platforms for neonatal sepsis. Biosens Bioelectron, 2020; 169, 112552. doi: 10.1016/j.bios.2020.112552 [3] McNamara JF, Harris PNA, Chatfield MD, et al. Long term sepsis readmission, mortality and cause of death following Gram negative bloodstream infection: a propensity matched observational linkage study. Int J Infect Dis, 2022; 114, 34−44. doi: 10.1016/j.ijid.2021.10.047 [4] Chousalkar K, Gast R, Martelli F, et al. Review of egg-related salmonellosis and reduction strategies in United States, Australia, United Kingdom and New Zealand. Crit Rev Microbiol, 2018; 44, 290−303. doi: 10.1080/1040841X.2017.1368998 [5] Chroni A, Rallidis L, Vassou D, et al. Identification and characterization of a rare variant in apolipoprotein A-IV, p. (V336M), and evaluation of HDL functionality in a Greek cohort with extreme HDL cholesterol levels. Arch Biochem Biophys, 2020; 696, 108655. doi: 10.1016/j.abb.2020.108655 [6] Akkoyun DÇ, Akyuz A, Doğan M, et al. Quercetin inhibits heart injury in lipopolysaccharide-induced endotoxemic model by suppressing the effects of reactive oxygen species. Anal Quant Cytopathol Histopathol, 2016; 38, 183−8. [7] Wei XQ, Meng XL, Yuan YX, et al. Quercetin exerts cardiovascular protective effects in LPS-induced dysfunction in vivo by regulating inflammatory cytokine expression, NF-κB phosphorylation, and caspase activity. Mol Cell Biochem, 2018; 446, 43−52. doi: 10.1007/s11010-018-3271-6 [8] Liu G, Wu KJ, Zhang L, et al. Metformin attenuated endotoxin-induced acute myocarditis via activating AMPK. Int Immunopharmacol, 2017; 47, 166−72. doi: 10.1016/j.intimp.2017.04.002 [9] Panaro MA, Acquafredda A, Cavallo P, et al. Inflammatory responses in embryonal cardiomyocytes exposed to LPS challenge: an in vitro model of deciphering the effects of LPS on the heart. Curr Pharm Des, 2010; 16, 754−65. doi: 10.2174/138161210790883516 [10] Shen L, Weber CR, Turner JR. The tight junction protein complex undergoes rapid and continuous molecular remodeling at steady state. J Cell Biol, 2008; 181, 683−95. doi: 10.1083/jcb.200711165 [11] Mokhtari B, Badalzadeh R. The potentials of distinct functions of autophagy to be targeted for attenuation of myocardial ischemia/reperfusion injury in preclinical studies: an up-to-date review. J Physiol Biochem, 2021; 77, 377−404. doi: 10.1007/s13105-021-00824-x [12] Larsen KE, Sulzer D. Autophagy in neurons: a review. Histol Histopathol, 2002; 17, 897−908. [13] Zhang C, Syed TW, Liu RJ, et al. Role of endoplasmic reticulum stress, autophagy, and inflammation in cardiovascular disease. Front Cardiovasc Med, 2017; 4, 29. doi: 10.3389/fcvm.2017.00029 [14] Mechesso AF, Quah YX, Park SC. Ginsenoside Rg3 reduces the adhesion, invasion, and intracellular survival of Salmonella enterica serovar Typhimurium. J Ginseng Res, 2021; 45, 75−85. doi: 10.1016/j.jgr.2019.09.002 [15] Chiu B, Jantuan E, Shen F, et al. Autophagy-inflammasome interplay in heart failure: a systematic review on basics, pathways, and therapeutic perspectives. Ann Clin Lab Sci, 2017; 47, 243−52. [16] Ashrafizadeh M, Tavakol S, Ahmadi Z, et al. Therapeutic effects of kaempferol affecting autophagy and endoplasmic reticulum stress. Phytother Res, 2020; 34, 911−23. doi: 10.1002/ptr.6577 [17] Mohammadinejad R, Ahmadi Z, Tavakol S, et al. Berberine as a potential autophagy modulator. J Cell Physiol, 2019; 234, 14914−26. doi: 10.1002/jcp.28325 [18] Nagata S, Nakano H. Apoptotic and non-apoptotic cell death. Springer. 2017. [19] Mughal W, Dhingra R, Kirshenbaum LA. Striking a balance: autophagy, apoptosis, and necrosis in a normal and failing heart. Curr Hypertens Rep, 2012; 14, 540−7. doi: 10.1007/s11906-012-0304-5 [20] Cong L, Bai YP, Guo ZG. The crosstalk among autophagy, apoptosis, and pyroptosis in cardiovascular disease. Front Cardiovasc Med, 2022; 9, 997469. doi: 10.3389/fcvm.2022.997469 [21] Dong Y, Chen HW, Gao JL, et al. Molecular machinery and interplay of apoptosis and autophagy in coronary heart disease. J Mol Cell Cardiol, 2019; 136, 27−41. doi: 10.1016/j.yjmcc.2019.09.001 [22] Zheng XT, Chen WW, Gong FC, et al. The role and mechanism of pyroptosis and potential therapeutic targets in sepsis: a review. Front Immunol, 2021; 12, 711939. doi: 10.3389/fimmu.2021.711939 [23] Wei SQ, Feng M, Zhang SD. Molecular characteristics of cell pyroptosis and its inhibitors: a review of activation, regulation, and inhibitors. Int J Mol Sci, 2022; 23, 16115. doi: 10.3390/ijms232416115 [24] Jia Y, Li DZ, Yu J, et al. Potential diabetic cardiomyopathy therapies targeting pyroptosis: a mini review. Front Cardiovasc Med, 2022; 9, 985020. doi: 10.3389/fcvm.2022.985020 [25] Zhang P, Zang MR, Sang ZZ, et al. Vitamin C alleviates LPS-induced myocardial injury by inhibiting pyroptosis via the ROS-AKT/mTOR signalling pathway. BMC Cardiovasc Disord, 2022; 22, 561. doi: 10.1186/s12872-022-03014-9 [26] Li Q, Zhang MM, Zhao Y, et al. Irisin protects against LPS-stressed cardiac damage through inhibiting inflammation, apoptosis, and pyroptosis. SHOCK, 2021; 56, 1009−18. doi: 10.1097/SHK.0000000000001775 [27] Ouyang CH, You JY, Xie ZL. The interplay between autophagy and apoptosis in the diabetic heart. J Mol Cell Cardiol, 2014; 71, 71−80. doi: 10.1016/j.yjmcc.2013.10.014 [28] Yu JH, Hu GL, Cao HB, et al. Quercetin ameliorates lipopolysaccharide-induced duodenal inflammation through modulating autophagy, programmed cell death and intestinal mucosal barrier function in chicken embryos. Animals, 2022; 12, 3524. doi: 10.3390/ani12243524 [29] Afrin S, Gasparrini M, Forbes-Hernandez TY, et al. Protective effects of Manuka honey on LPS-treated RAW 264.7 macrophages. Part 1: Enhancement of cellular viability, regulation of cellular apoptosis and improvement of mitochondrial functionality. Food Chem Toxicol, 2018; 121, 203−13. doi: 10.1016/j.fct.2018.09.001 [30] Häkkinen SH, Kärenlampi SO, Heinonen IM, et al. Content of the flavonols quercetin, myricetin, and kaempferol in 25 edible berries. J Agric Food Chem, 1999; 47, 2274−9. doi: 10.1021/jf9811065 [31] Gasparrini M, Afrin S, Forbes-Hernández TY, et al. Protective effects of Manuka honey on LPS-treated RAW 264.7 macrophages. Part 2: Control of oxidative stress induced damage, increase of antioxidant enzyme activities and attenuation of inflammation. Food Chem Toxicol, 2018; 120, 578−87. doi: 10.1016/j.fct.2018.08.001 [32] Gasparrini M, Forbes-Hernandez TY, Giampieri F, et al. Anti-inflammatory effect of strawberry extract against LPS-induced stress in RAW 264.7 macrophages. Food Chem Toxicol, 2017; 102, 1−10. doi: 10.1016/j.fct.2017.01.018 [33] Ulusoy HG, Sanlier N. A minireview of quercetin: from its metabolism to possible mechanisms of its biological activities. Crit Rev Food Sci Nutr, 2020; 60, 3290−303. doi: 10.1080/10408398.2019.1683810 [34] Lotfi N, Yousefi Z, Golabi M, et al. The potential anti-cancer effects of quercetin on blood, prostate and lung cancers: an update. Front Immunol, 2023; 14, 1077531. doi: 10.3389/fimmu.2023.1077531 [35] Paskeh MDA, Entezari M, Clark C, et al. Targeted regulation of autophagy using nanoparticles: new insight into cancer therapy. Biochim Biophys Acta Mol Basis Dis, 2022; 1868, 166326. doi: 10.1016/j.bbadis.2021.166326 [36] Ashrafizadeh M, Paskeh MDA, Mirzaei S, et al. Targeting autophagy in prostate cancer: preclinical and clinical evidence for therapeutic response. J Exp Clin Cancer Res, 2022; 41, 105. doi: 10.1186/s13046-022-02293-6 [37] Liu J, Wang SC, Zhang QJ, et al. Selenomethionine alleviates LPS-induced chicken myocardial inflammation by regulating the miR-128-3p-p38 MAPK axis and oxidative stress. Metallomics, 2020; 12, 54−64. doi: 10.1039/c9mt00216b [38] Frantz S, Kobzik L, Kim YD, et al. Toll4 (TLR4) expression in cardiac myocytes in normal and failing myocardium. J Clin Invest, 1999; 104, 271−80. doi: 10.1172/JCI6709 [39] Zanoni I, Bodio C, Broggi A, et al. Similarities and differences of innate immune responses elicited by smooth and rough LPS. Immunol Lett, 2012; 142, 41−7. doi: 10.1016/j.imlet.2011.12.002 [40] Kogut M, He HQ, Kaiser P. Lipopolysaccharide binding protein/CD14/TLR4-dependent recognition of salmonella LPS induces the functional activation of chicken heterophils and up-regulation of pro-inflammatory cytokine and chemokine gene expression in these cells. Anim Biotechnol, 2005; 16, 165−81. doi: 10.1080/10495390500264896 [41] Panaro MA, Cianciulli A, Gagliardi N, et al. CD14 major role during lipopolysaccharide-induced inflammation in chick embryo cardiomyocytes. FEMS Immunol Med Microbiol, 2008; 53, 35−45. doi: 10.1111/j.1574-695X.2008.00397.x [42] Rossetti C, Peri F. The role of toll-like receptor 4 in infectious and non Infectious Inflammation. Springer. 2021. [43] De Vicente LG, Muñoz VR, Pinto AP, et al. TLR4 deletion increases basal energy expenditure and attenuates heart apoptosis and ER stress but mitigates the training-induced cardiac function and performance improvement. Life Sci, 2021; 285, 119988. doi: 10.1016/j.lfs.2021.119988 [44] Karnati HK, Pasupuleti SR, Kandi R, et al. TLR-4 signalling pathway: MyD88 independent pathway up-regulation in chicken breeds upon LPS treatment. Vet Res Commun, 2015; 39, 73−8. doi: 10.1007/s11259-014-9621-2 [45] Wang XP, Guo DQ, Li WL, et al. Danshen (Salvia miltiorrhiza) restricts MD2/TLR4-MyD88 complex formation and signalling in acute myocardial infarction-induced heart failure. J Cell Mol Med, 2020; 24, 10677−92. doi: 10.1111/jcmm.15688 [46] Zhang LL, Zhang C, Peng JP. Application of nanopore sequencing technology in the clinical diagnosis of infectious diseases. Biomed Environ Sci, 2022; 35, 381−92. [47] Kogut MH, Rothwell L, Kaiser P. Priming by recombinant chicken interleukin-2 induces selective expression of IL-8 and IL-18 mRNA in chicken heterophils during receptor-mediated phagocytosis of opsonized and nonopsonized Salmonella enterica serovar enteritidis. Mol Immunol, 2003; 40, 603−10. doi: 10.1016/j.molimm.2003.08.002 [48] Akseh S, Nemati M, Zamani-Gharehchamani E, et al. Amnion membrane proteins attenuate LPS-induced inflammation and apoptosis by inhibiting TLR4/NF-κB pathway and repressing MicroRNA-155 in rat H9c2 cells. Immunopharmacol Immunotoxicol, 2021; 43, 487−94. doi: 10.1080/08923973.2021.1945086 [49] Handley SA, Miller VL. General and specific host responses to bacterial infection in Peyer's patches: a role for stromelysin-1 (matrix metalloproteinase-3) during Salmonella enterica infection. Mol Microbiol, 2007; 64, 94−110. doi: 10.1111/j.1365-2958.2007.05635.x [50] Lalu MM, Csont T, Schulz R. Matrix metalloproteinase activities are altered in the heart and plasma during endotoxemia. Crit Care Med, 2004; 32, 1332−7. doi: 10.1097/01.CCM.0000127778.16609.EC [51] Iriti M, Kubina R, Cochis A, et al. Rutin, a quercetin glycoside, restores chemosensitivity in human breast cancer cells. Phytother Res, 2017; 31, 1529−38. doi: 10.1002/ptr.5878 [52] Liu XC, Zheng L, Liu M, et al. Protective effects of rutin on lipopolysaccharide-induced heart injury in mice. J Toxicol Sci, 2018; 43, 329−37. doi: 10.2131/jts.43.329 [53] Shu J, Gu YW, Jin L, et al. Matrix metalloproteinase 3 regulates angiotensin II-induced myocardial fibrosis cell viability, migration and apoptosis. Mol Med Rep, 2021; 23, 151. [54] Collins MM, Ryan AK. Manipulating claudin expression in avian embryos. Methods Mol Biol, 2011; 762, 195−212. [55] Simard A, Di Pietro E, Ryan AK. Gene expression pattern of Claudin-1 during chick embryogenesis. Gene Expr Patterns, 2005; 5, 553−60. doi: 10.1016/j.modgep.2004.10.009 [56] Simard A, Di Pietro E, Young CR, et al. Alterations in heart looping induced by overexpression of the tight junction protein Claudin-1 are dependent on its C-terminal cytoplasmic tail. Mech Dev, 2006; 123, 210−27. doi: 10.1016/j.mod.2005.12.004 [57] Kostin S. Zonula occludens-1 and connexin 43 expression in the failing human heart. J Cell Mol Med, 2007; 11, 892−5. doi: 10.1111/j.1582-4934.2007.00063.x [58] El Refaey M, Coles S, Musa H, et al. Altered expression of zonula occludens-1 affects cardiac Na+ channels and increases susceptibility to ventricular arrhythmias. Cells, 2022; 11, 665. doi: 10.3390/cells11040665 [59] Zhang JL, Vincent KP, Peter AK, et al. Cardiomyocyte expression of ZO-1 is essential for normal atrioventricular conduction but does not alter ventricular function. Circ Res, 2020; 127, 284−97. doi: 10.1161/CIRCRESAHA.119.315539 [60] Chen Y, Liu YQ, Dorn II GW. Mitochondrial fusion is essential for organelle function and cardiac homeostasis. Circ Res, 2011; 109, 1327−31. doi: 10.1161/CIRCRESAHA.111.258723 [61] Yu WC, Mei X, Zhang Q, et al. Yap overexpression attenuates septic cardiomyopathy by inhibiting DRP1-related mitochondrial fission and activating the ERK signaling pathway. J Recept Signal Transduct, 2019; 39, 175−86. doi: 10.1080/10799893.2019.1641822 [62] Bullon P, Cordero MD, Quiles JL, et al. Mitochondrial dysfunction promoted by Porphyromonas gingivalis lipopolysaccharide as a possible link between cardiovascular disease and periodontitis. Free Radic Biol Med, 2011; 50, 1336−43. doi: 10.1016/j.freeradbiomed.2011.02.018 [63] Xu T, Dong Q, Luo YX, et al. Porphyromonas gingivalis infection promotes mitochondrial dysfunction through Drp1-dependent mitochondrial fission in endothelial cells. Int J Oral Sci, 2021; 13, 28. doi: 10.1038/s41368-021-00134-4 [64] Tong MM, Zablocki D, Sadoshima J. The role of Drp1 in mitophagy and cell death in the heart. J Mol Cell Cardiol, 2020; 142, 138−45. doi: 10.1016/j.yjmcc.2020.04.015 [65] Zhao QC, Yan S, Lu J, et al. Drp1 regulates transcription of ribosomal protein genes in embryonic hearts. J Cell Sci, 2022; 135, jcs258956. doi: 10.1242/jcs.258956 [66] Sharp WW, Fang YH, Han M, et al. Dynamin-related protein 1 (Drp1)-mediated diastolic dysfunction in myocardial ischemia-reperfusion injury: therapeutic benefits of Drp1 inhibition to reduce mitochondrial fission. FASEB J, 2014; 28, 316−26. doi: 10.1096/fj.12-226225 [67] Ikeda Y, Shirakabe A, Maejima Y, et al. Endogenous Drp1 mediates mitochondrial autophagy and protects the heart against energy stress. Circ Res, 2015; 116, 264−78. doi: 10.1161/CIRCRESAHA.116.303356 [68] Ren J, Sowers JR, Zhang YM. Autophagy and cardiometabolic diseases. Academic Press. 2018. [69] Chang X, He YH, Wang L, et al. Puerarin alleviates LPS-induced H9C2 cell injury by inducing mitochondrial autophagy. J Cardiovasc Pharmacol, 2022; 80, 600−8. [70] Ben-Shaul V, Lomnitski L, Nyska A, et al. The effect of natural antioxidants, NAO and apocynin, on oxidative stress in the rat heart following LPS challenge. Toxicol Lett, 2001; 123, 1−10. [71] Sul OJ, Ra SW. Quercetin prevents LPS-induced oxidative stress and inflammation by modulating NOX2/ROS/NF-kB in lung epithelial cells. Molecules, 2021; 26, 6949. doi: 10.3390/molecules26226949 [72] Lushnikova EL, Nepomnyashchikh LM, Pichigin VI, et al. Expression of mRNA of apolipoprotein E, apolipoprotein A-IV, and matricellular proteins in the myocardium and intensity of fibroplastic processes during experimental hypercholesterolemia. Bull Exp Biol Med, 2013; 156, 271−5. doi: 10.1007/s10517-013-2328-5 [73] Zhang WQ, Liu XH, Zhou JT, et al. Apolipoprotein A-IV restrains fat accumulation in skeletal and myocardial muscles by inhibiting lipogenesis and activating PI3K-AKT signalling. Arch Physiol Biochem, 2023, 1-11. [74] Wang Y, Zhang ZZ, Wu Y, et al. Quercetin postconditioning attenuates myocardial ischemia/reperfusion injury in rats through the PI3K/Akt pathway. Braz J Med Biol Res, 2013; 46, 861−7. doi: 10.1590/1414-431X20133036 [75] Ravindranath TM, Goto M, Bakr S, et al. LPS-induced changes in myocardial markers in neonatal rats. Biol Neonate, 2003; 84, 319−24. doi: 10.1159/000073641 [76] Lado-Abeal J, Martinez-Sánchez N, Cocho JA, et al. Lipopolysaccharide (LPS)-induced septic shock causes profound changes in myocardial energy metabolites in pigs. Metabolomics, 2018; 14, 131. doi: 10.1007/s11306-018-1433-x [77] Yamaguchi O. Autophagy in the Heart. Circ J, 2019; 83, 697−704. doi: 10.1253/circj.CJ-18-1065 [78] Drosatos K, Pollak NM, Pol CJ, et al. Cardiac myocyte KLF5 regulates Ppara expression and cardiac function. Circ Res, 2016; 118, 241−53. doi: 10.1161/CIRCRESAHA.115.306383 [79] Kumari R, Ray AG, Mukherjee D, et al. Downregulation of PTEN promotes autophagy via concurrent reduction in apoptosis in cardiac hypertrophy in PPAR α-/- mice. Front Cardiovasc Med, 2022; 9, 798639. doi: 10.3389/fcvm.2022.798639 [80] Saikia R, Joseph J. AMPK: a key regulator of energy stress and calcium-induced autophagy. J Mol Med, 2021; 99, 1539−51. doi: 10.1007/s00109-021-02125-8 [81] Eisenberg-Lerner A, Bialik S, Simon HU, et al. Life and death partners: apoptosis, autophagy and the cross-talk between them. Cell Death Differ, 2009; 16, 966−75. doi: 10.1038/cdd.2009.33 [82] Kajstura J, Mansukhani M, Cheng W, et al. Programmed cell death and expression of the protooncogene bcl-2 in myocytes during postnatal maturation of the heart. Exp Cell Res, 1995; 219, 110−21. doi: 10.1006/excr.1995.1211 [83] Grzegorzewska AK, Hrabia A, Paczoska-Eliasiewicz HE. Localization of apoptotic and proliferating cells and mRNA expression of caspases and Bcl-2 in gonads of chicken embryos. Acta Histochem, 2014; 116, 795−802. doi: 10.1016/j.acthis.2014.01.012 [84] Sun YX, Cai Y, Qian SH, et al. Beclin-1 improves mitochondria-associated membranes in the heart during endotoxemia. FASEB Bioadv, 2021; 3, 123−35. doi: 10.1096/fba.2020-00039 [85] Sun YX, Yao X, Zhang QJ, et al. Beclin-1-dependent autophagy protects the heart during sepsis. Circulation, 2018; 138, 2247−62. doi: 10.1161/CIRCULATIONAHA.117.032821 [86] Maejima Y, Isobe M, Sadoshima J. Regulation of autophagy by Beclin 1 in the heart. J Mol Cell Cardiol, 2016; 95, 19−25. doi: 10.1016/j.yjmcc.2015.10.032 [87] Haq S, Grondin J, Banskota S, et al. Autophagy: roles in intestinal mucosal homeostasis and inflammation. J Biomed Sci, 2019; 26, 19. doi: 10.1186/s12929-019-0512-2 [88] Liao ZH, Dai ZK, Cai CY, et al. Knockout of Atg5 inhibits proliferation and promotes apoptosis of DF-1 cells. In Vitro Cell Dev Biol Anim, 2019; 55, 341−8. doi: 10.1007/s11626-019-00342-7 [89] Zhu CM, Zhang SY, Liu D, et al. A novel gene prognostic signature based on differential DNA methylation in breast cancer. Front Genet, 2021; 12, 742578. doi: 10.3389/fgene.2021.742578 [90] Liu L, Chen ML, Lin K, et al. Inhibiting caspase-12 mediated inflammasome activation protects against oxygen-glucose deprivation injury in primary astrocytes. Int J Med Sci, 2020; 17, 1936−45. doi: 10.7150/ijms.44330 [91] Nakagawa T, Zhu H, Morishima N, et al. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-β. Nature, 2000; 403, 98−103. doi: 10.1038/47513 [92] Morishima N, Nakanishi K, Takenouchi H, et al. An endoplasmic reticulum stress-specific caspase cascade in apoptosis. Cytochrome c-independent activation of caspase-9 by caspase-12. J Biol Chem, 2002; 277, 34287−94. doi: 10.1074/jbc.M204973200 [93] Salvamoser R, Brinkmann K, O'Reilly LA, et al. Characterisation of mice lacking the inflammatory caspases-1/11/12 reveals no contribution of caspase-12 to cell death and sepsis. Cell Death Differ, 2019; 26, 1124−37. doi: 10.1038/s41418-018-0188-2 [94] Liu HH, Wang SJ, Gong LJ, et al. SIRT6 ameliorates LPS-induced apoptosis and tight junction injury in ARDS through the ERK1/2 pathway and autophagy. Int J Med Sci, 2023; 20, 581−94. doi: 10.7150/ijms.80920 [95] Wang JY, Luan YY, Fan EK, et al. TBK1/IKKε negatively regulate lps-induced neutrophil necroptosis and lung inflammation. SHOCK, 2021; 55, 338−48. doi: 10.1097/SHK.0000000000001632 [96] Liu YL, Hsu CC, Huang HJ, et al. Gallic acid attenuated LPS-induced neuroinflammation: protein aggregation and necroptosis. Mol Neurobiol, 2020; 57, 96−104. doi: 10.1007/s12035-019-01759-7 [97] Wang YC, Jiang L, Li YF, et al. Excessive selenium supplementation induced oxidative stress and endoplasmic reticulum stress in chicken spleen. Biol Trace Elem Res, 2016; 172, 481−7. doi: 10.1007/s12011-015-0596-9 -
 23031+Supplementary Materials.pdf
23031+Supplementary Materials.pdf

-

 点击查看大图
点击查看大图
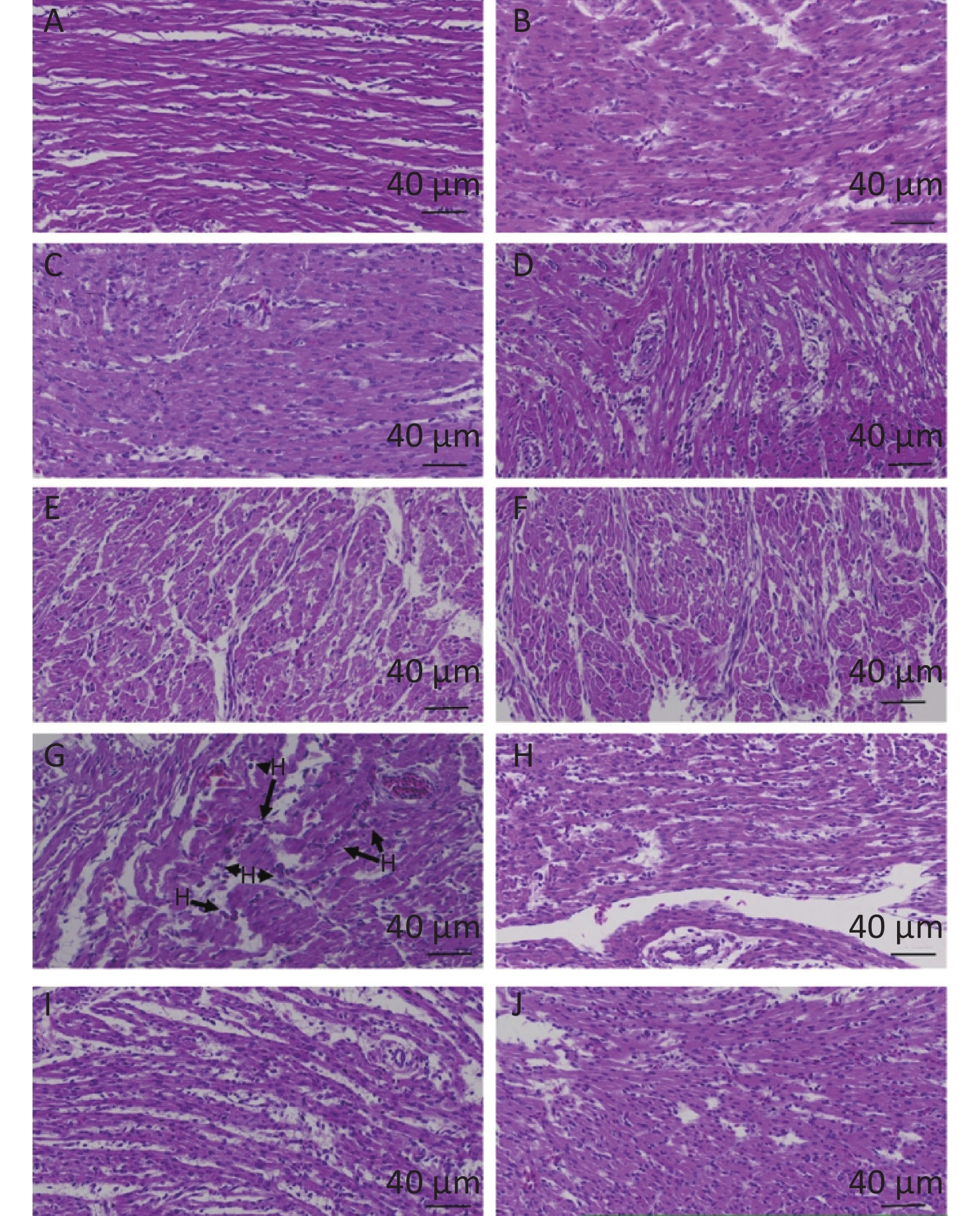
计量
- 文章访问数: 470
- HTML全文浏览量: 190
- PDF下载量: 24
- 被引次数: 0

 Quick Links
Quick Links