

 下载:
下载:

-
The rise of antibiotic-resistant bacteria has become a serious challenge in healthcare, which is exacerbated by limited development of new antimicrobial agents and the increasing prevalence of multidrug-resistant (MDR) bacteria, often leading to treatment failure. Aeromonas spp., gram-negative bacteria commonly found in aquatic and terrestrial environments, are frequently implicated in both intestinal and extraintestinal infections[1]. Increasing numbers of MDR Aeromonas spp. have been reported, particularly among clinical isolates associated with extraintestinal infections and in seafood. Gram-negative bacteremia is empirically treated with carbapenems, and in accordance with the Sanford Guide to Antimicrobial Therapy, ciprofloxacin or levofloxacin is used as the first-line medication for Aeromonas infection. However, carbapenem-resistant and ciprofloxacin-resistant Aeromonas strains have been emerging worldwide[2]. For example, recent studies in China, Korea, and the USA reported the rates of ciprofloxacin resistance among clinical Aeromonas isolates as 16.5%, 10.1%, and 7.3%, and those of imipenem resistance as 12.8%, 9.8%, and 37%, respectively[2-4]. Aeromonas is a natural reservoir for various classes of β-lactamases genes, including blaTEM, blaCTX, blaOXA, blaNDM, blaVIM, blaKPC, and blaCphA[3,5], with blaCphA as the most prevalent[3]. Aeromonas isolates also harbor various plasmid replicons, with ColE, IncR, and IncU being the most frequently detected[5].
This study was aimed to genetically characterize MDR Aeromonas strains and analyze the antibiotic resistance genes (ARGs) and plasmids in these strains. A 10-year retrospective study of 60 Aeromonas strains was conducted at Beijing Friendship Hospital, the Second Affiliated Hospital of Capital Medical University, Beijing. Seventeen Aeromonas strains were collected from Feb 2015 to Dec 2017[6], and 43 strains were isolated from Jan 2018 to June 2025 (Supplementary Table S1). These strains were genetically analyzed and compared. Six ciprofloxacin-resistant Aeromonas spp. were chosen for further genomic analysis.
Genomic DNA was extracted using the Wizard Genomic DNA Extraction Kit. The extracted gDNA was split into two portions: one was randomly fragmented into a 300 bp insert library and sequenced on an Illumina NovaSeq 6000 platform (PE150), and the other was sheared using g-TUBEs, selected for size, and used to construct an SMRTbell library, which was sequenced on a PacBio Sequel II system. Gene prediction was performed using Prodigal v2.6.3, and genome annotation was carried out using the Rapid Annotation using Subsystem Technology (RAST) server.
Average nucleotide identity (ANI) was analysed, and strains with an ANI value > 95% were considered to be the same species. Potential ARGs were predicted using the Comprehensive Antibiotic Resistance Database (CARD), and the BLAST+ parameters were an E-value of 1 × 10-5, sequence identity ≥90%, and length coverage ≥90%. Open reading frames (ORF) were predicted and annotated using RAST and the Basic Local Alignment Search Tool (BLAST) from the National Center for Biotechnology Information (NCBI). Plasmid replicons were identified using MOB-suite and PlasmidFinder v2.1 with thresholds of ≥95% for identity and ≥60% for coverage. The structure of the exogenous insertion region of the plasmid was annotated using ISfinder, INTEGRALL, and the Transposon Registry. Inkscape v0.48.1 was used to draw circular maps of the plasmids. Protein three-dimensional structure was predicted using the SWISS-MODEL online platform. PyMOL v2.5.7 software was used for protein visualization and structural analysis.
Gene deletion was performed using the chloramphenicol-resistant suicide plasmid pRE112 and the primers listed in Supplementary Table S2. Briefly, PCR fragments containing the region flanking cphA were amplified, and then an overlapping fragment was generated using primers cphA112upF/cphA112upR and cphA112dnF/cphA112dnR. The fragments were then digested and ligated into the suicide plasmid pRE112. The resulting plasmid, pRE112-cphA, was conjugated into A. hydrophila BJ054 from E. coli SM10 λpir. Sucrose (15% W/V) plates containing chloramphenicol (30 μg/ml) and without salt were used to select for deletion mutants, which were then sequenced and verified. A cphA complementation strain was constructed using the isopropyl-thio-β-D-galactoside (IPTG)‐inducible expression plasmid pSRKTc (which carries tetracycline resistance). The cphA fragment was amplified using the primers cphAF/cphAR, digested, and ligated into pSRKTc. The resulting construct, pSRK-cphA, was conjugated into A. hydrophila BJ054ΔcphA from E. coli SM10 λpir.
The complete genome and plasmid sequences of the strains were deposited in GenBank under the following accession numbers: BJ014 chromosome and its plasmids, CP180559–CP180563; BJ017, JBKGFS000000000; BJ018 chromosome and its plasmids, CP180564–CP180568; BJ042, CP180569; BJ093, CP180570; BJ054, JBKGFS000000000; and CphA-18, WP_411637240.1.
Strains BJ014, BJ017, BJ018, and BJ054 were identified as Aeromonas hydrophila, whereas strains BJ042 and BJ093 were identified as Aeromonas caviae (Supplementary Figure S1). The Type Genome Server (TYGS) results were consistent with those of the ANI analysis. Complete genomic sequencing of BJ014 and BJ018 revealed that each contained four circular plasmids tentatively named pBJ014-1/2/3/4 and pBJ018-1/2/3/4, respectively. No plasmids were found in strains BJ017, BJ054, BJ042, or BJ093.
Antimicrobial testing reports of the 43 strains isolated in 2018–2025 were compared with those of the 17 strains isolated in 2014–2017 (Table 1), which showed that of the 43 more recent isolates, 12 (27.9%) exhibited resistance to imipenem, and seven (16.3%) exhibited resistance to ciprofloxacin, indicating that the ciprofloxacin resistance rate had decreased, whereas the rate of imipenem resistance had increased (Table 1). Two strains were resistant to both imipenem and ciprofloxacin, and these were isolated during 2021–2022.
Table 1. Antibiotic susceptibilities of 60 Aeromonas spp. isolates collected from extraintestinal infections at Beijing Friendship Hospital, the Second Affiliated Hospital of Capital Medical University, Beijing in 2014–2025.
Antibiotic Total (n = 60) 2014–2017 (n = 17) 2018–2025 (n = 43) R1 [n (%)] I [n (%)] S [n (%)] R [n (%)] I [n (%)] S [n (%)] R [n (%)] I [n (%)] S [n (%)] Imipenema 13 (21.7) 4 (6.7) 43 (71.7) 1 (5.9) 2 (11.8) 14 (82.4) 12 (27.9) 2 (4.7) 29 (67.4) Cefepimea 8 (4.3) 6 (2.6) 46 (93.0) 4 (23.5) 3 (17.6) 10 (58.8) 4 (9.3) 3 (7.0) 36 (83.7) Ciprofloxacina 13 (21.7) 0 (0.0) 47 (78.3) 6 (35.3) 0 (0.0) 11 (64.7) 7 (16.3) 0 (0.0) 36 (83.7) Trimethoprim- Sulfamethoxazolea 13 (21.7) 0 (0.0) 47 (78.3) 5 (29.4) 0 (0.0) 12 (70.6) 8 (18.6) 0 (0.0) 35 (81.4) Note. 1 R, Resistant; I, Intermediate; S, Sensitive.
a Breakpoints are based on the CLSIM45-A3 standards for Aeromonas spp.
b Other breakpoints are based on the CLSIM100- ED33 criteria for Enterobacteriaceae.Six strains in the genomic analysis were MDR (Supplementary Table S3) but were sensitive to imipenem; three A. hydrophila strains, BJ017, BJ018, and BJ054, were resistant to ertapenem; A. hydrophila strain BJ014 had intermediate resistance to ertapenem; and A. caviae strains BJ042 and BJ093 were sensitive to ertapenem.
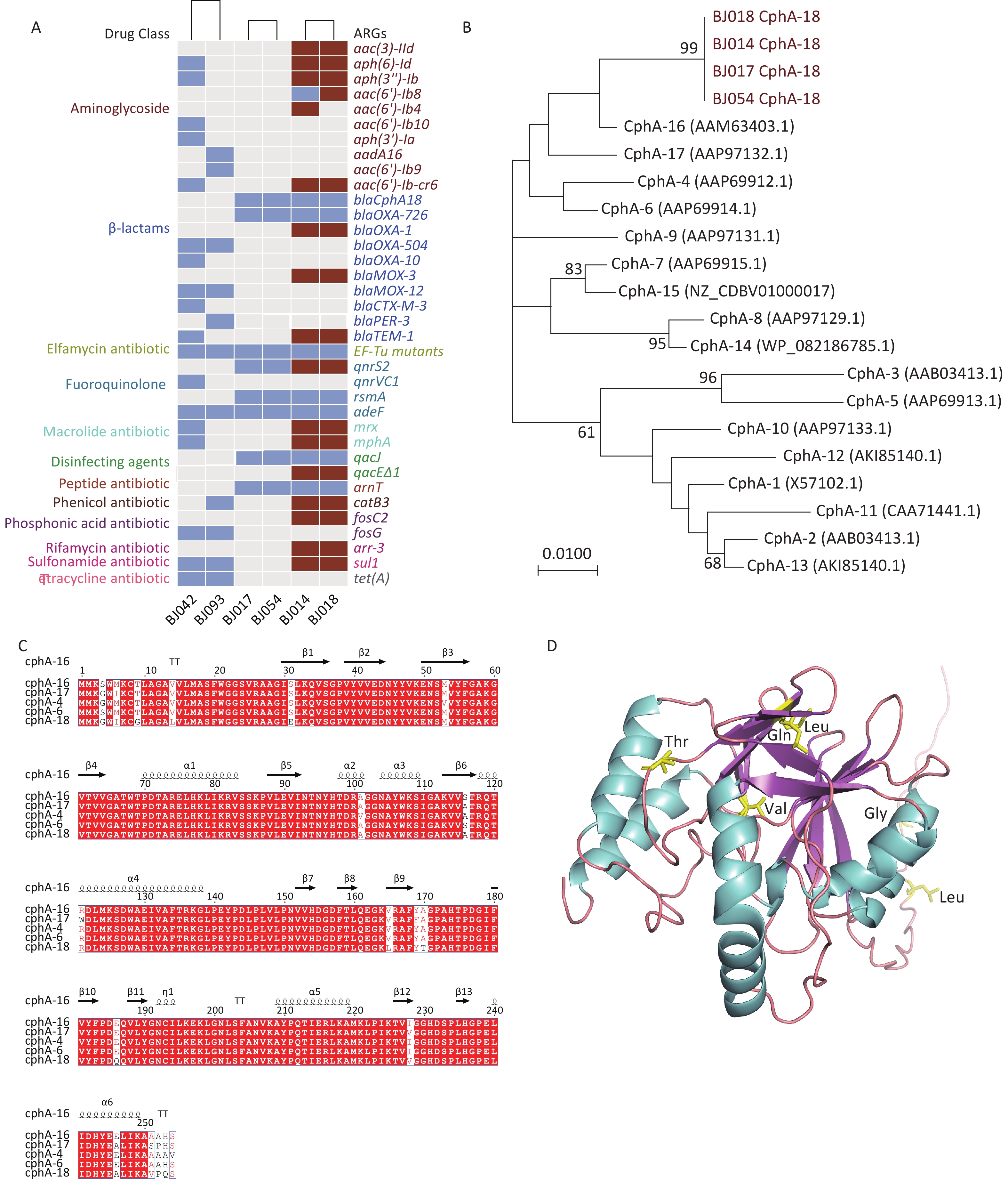
The strains were a rich reservoir of ARGs, with 36 non-duplicate ARGs detected (Figure 1A). The ARGs in strains BJ017, BJ054, BJ042, and BJ093 were all located on the chromosome (represented as blue rectangles in Figure 1A), whereas some ARGS in strains BJ014 and BJ018 were located on the chromosome, while others were located on plasmids (red rectangles in Figure 1A). Strain BJ014 harbored the highest number of ARGs. A. caviae strains BJ042 and BJ093 harbored the tetracycline resistance gene tet (A) and were insensitive to tetracycline and doxycycline. All six strains were resistant to ciprofloxacin. The known resistance amino acid substitutions Ser83Ile on gyrA and Ser87Ile on parC were detected in these strains as well as the quinolone resistance gene adeF. In addition, A. caviae strain BJ042 carried qnrVC1, and all A. hydrophila strains carried qnrS2 and rsmA. The antibiotic resistance gene qnrVC, which was first reported in Vibrio spp. and was subsequently detected in Klebsiella and Enterobacter strains isolated from the coastal waters of Brazil[7], was also detected in A. caviae strain BJ042 (specifically qnrVC1), indicating its widespread, cross-species transmission.
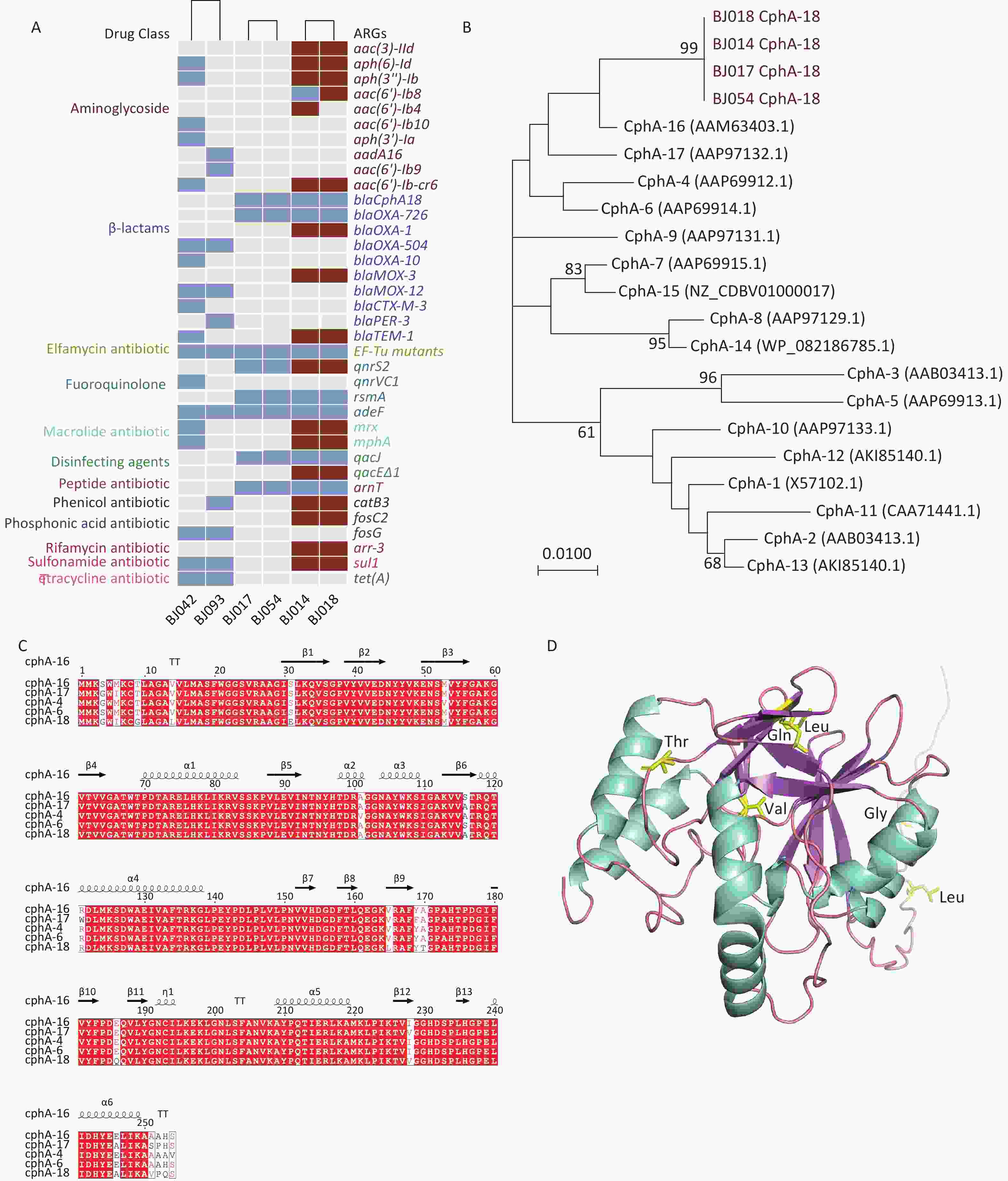
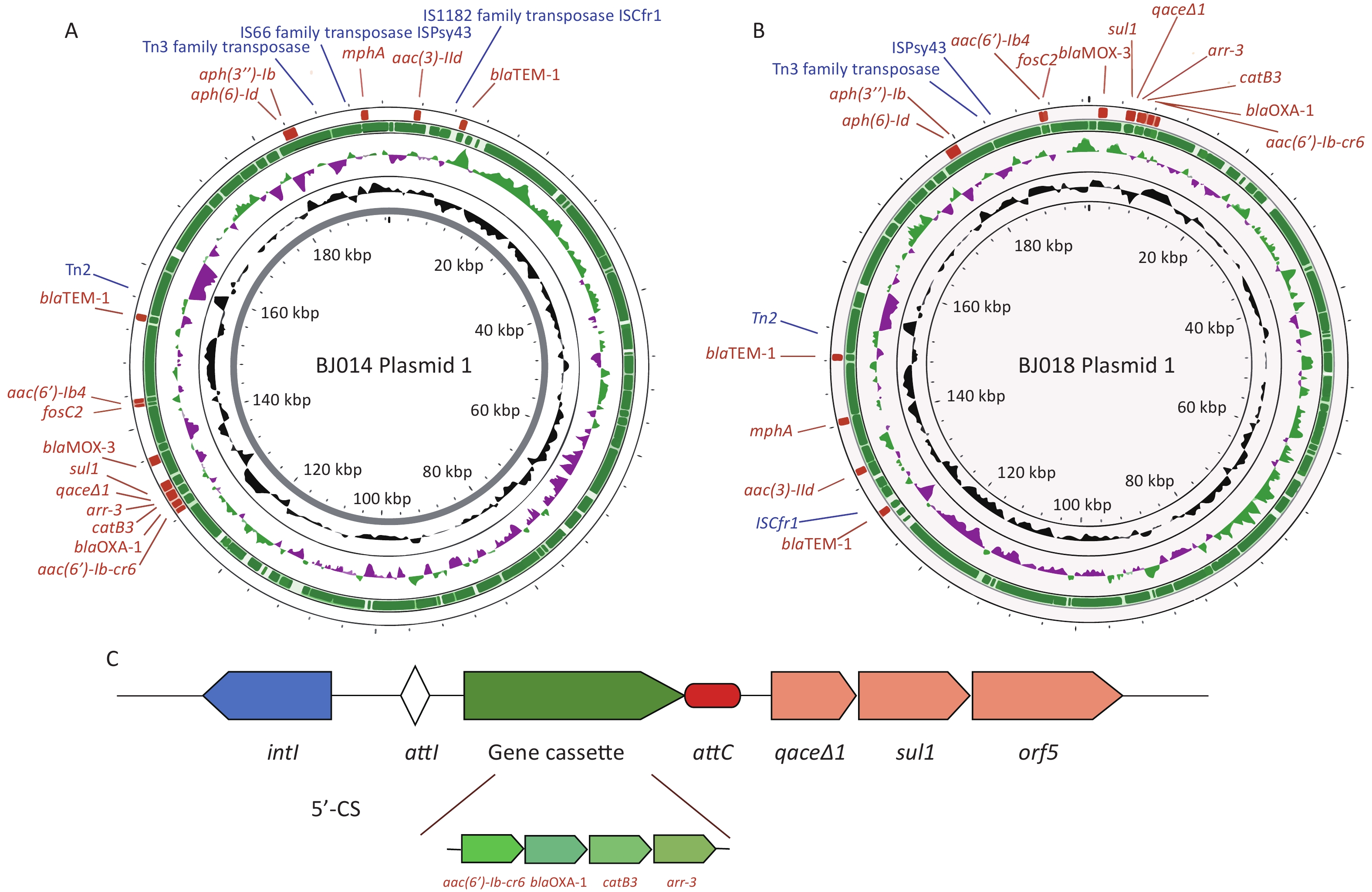
Figure 1. (A) The antibiotic resistance genes of six clinical Aeromonas strains. Maroon rectangles represent genes located on plasmids, and blue rectangles represent genes located in chromosomes. (B) Phylogenetic analysis of CphA enzymes. (C) Amino acid alignment of CphA-18 with four other CphA variants. (D) The predicted protein structure of CphA-18. Helices are shown in teal, sheets are shown in purple, and loops are shown in pink.
A. caviae strain BJ042 carried the β-lactamase genes blaOXA-504, blaOXA-10, blaMOX-12, and blaCTX-M-3, while A. caviae strain BJ093 carried blaOXA-504, blaMOX-12, and blaPER-3, and both showed resistance to ceftriaxone and cefepime. All A. hydrophila strains harbored blaCphA-18 and blaOXA-726; and BJ014 and BJ018 also harbored blaOXA-1, blaMOX-3, and blaTEM-1.
CphA—a metallo-β-lactamase (MBL) classified under subclass B2—is characterized by its narrow specificity, especially against carbapenems[8]. A gene encoding a new variant of the CphA enzyme, cphA-18, was identified on the chromosomes of four A. hydrophila strains (Figure 1B). The new variant harbored all the characteristic conserved structural elements of CphA. Among previously reported CphA enzymes, CphA-18 was the closest to CphA-16 (Figure 1C). Compared with CphA-16, CphA-18 has seven amino acid substitutions: T9G, V14L, V165L, A170T, E186Q, I228V, and E246A. The predicted protein structure is shown in Figure 1D.
Studies have shown that site-directed mutants of CphA have altered activity spectra[8]. Some strains that carry cphA are sensitive to carbapenems[3]. To assess whether cphA-18 confers resistance to carbapenem antibiotics, a cphA deletion strain (BJ054ΔcphA) and a cphA complementation strain (BJ054ΔcphA-pSRKcphA) were constructed from A. hydrophila strain BJ054. Mutant BJ054ΔcphA showed a 4-fold decrease in resistance to imipenem compared with wild-type BJ054, with a MIC of 0.25 µg/mL (Supplementary Table S4); a 4- to 8-fold decrease in resistance to ertapenem, with MICs of 0.5–1 µg/mL; and no change in resistance to meropenem, with a MIC of 0.06 µg/mL. The complemented mutant, BJ054ΔcphA-pSRKcphA, exhibited a 2-fold increase in resistance to imipenem compared with BJ054ΔcphA, with a MIC of 0.5 µg/mL; a 2- to 8-fold increase in resistance to ertapenem, with MICs of 2–4 µg/mL; and no change in resistance to meropenem, with a MIC of 0.06 µg/mL. This showed that CphA-18 has its own activity spectrum. The addition of pSRKcphA to BJ054ΔcphA increased the MIC of imipenem and ertapenem from 0.25 to 0.5 µg/mL and from 0.5–1 to 2–4 µg/mL, respectively; however, the increased levels were lower than those of BJ054 at 1 µg/mL and 4 µg/mL, respectively. This may be because of the relatively lower efficiency of the pSRK promoter in Aeromonas.
Infections caused by cphA-carrying A. hydrophila have been reported worldwide[9]. Our sequencing detected cphA-1 genes in A. aquariorum strains BJ001, BJ005, and BJ065, and a cphA-2 gene in A. veronii strain BJ005, which was collected in our previous study and was found to be resistant to imipenem[6] (data not shown). CphA-18 was also found in strain CN18A183, which was isolated from a stool sample in Anhui, China in 2018, (sequence deposited in 2025), indicating widespread transmission of cphA-18. Our analysis showed that plasmids pBJ014-1 and pBJ018-1 from Aeromonas hydrophila strains BJ014 and BJ018, respectively, are very similar, and their genetic characteristics are listed in Supplementary Table S5. Sequencing of pBJ014-1 and pBJ018-1 showed that they are circular, with a length of 196,803 bp and an average G+C content of 53.6% (Figure 2A and B). A total of 191 and 192 coding sequences were predicted, respectively, and 62.3% of the predicted sequences encoded hypothetical proteins. We then used PlasmidFinder 2.0 to identify the replicon (rep) sequence of pBJ014 and pBJ018; however, no hits were found even when the threshold for minimum percent identity was set at 50%. Hence, these plasmids were identified as non-typeable. The modular structure of pBJ014-1 and pBJ018-1 is closely related to that of pQZ124-211 (a 203-kb plasmid of A. hydrophila), with 92% query coverage and 99.8% nucleotide identity (Supplementary Table S6). The plasmid harbors several resistance genes and mobile elements. Annotation revealed that blaOXA-1 was located in a class 1 integron with the structure aac(6')-Ib-cr6-blaOXA-1-catB3-arr-3-qacE∆1-sul1 led by intI1 (Figure 2C). BLASTn analysis revealed that the blaOXA-1 genetic environment was similar to that of blaOXA-1 in plasmids from Klebsiella pneumoniae (CP093457.1), E. coli (CP125007.1), Salmonella enterica (CP094271.1), and Citrobacter freundii (CP104961.1). An MDR region downstream of the blaOXA-1-bearing integron—which includes the blaMOX-3, fosC2, aac(6')-Ib4, and blaTEM-1 genes in pBJ014-1 and the blaMOX-3, fosC2, aac(6')-Ib4, aph(3'')-Ib, and aph(6)-Id genes in pBJ018-1—was clustered and flanked by mobile elements.
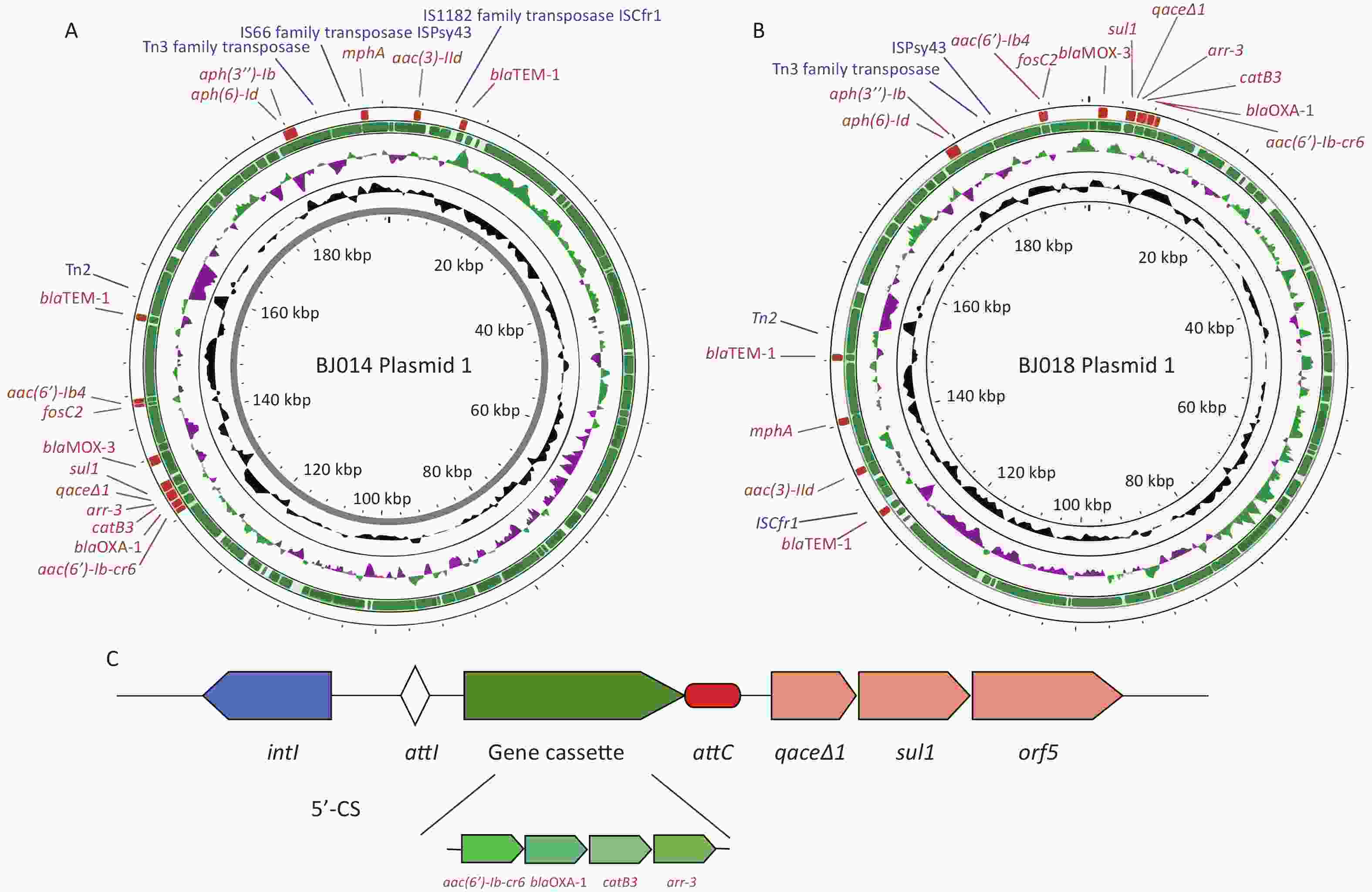
Figure 2. (A & B) Maps of the complete sequences of pBJ014-1 (A) and pBJ018-1 (B). (C) The genetic structure of the class 1 integron present in pBJ014-1 and pBJ018-1.
The ability of Aeromonas to exchange and gain plasmids is well known, and Aeromonas has an open pan-genome with extensive variability[10]. Our characterization of pBJ014-1 and pBJ018-1 (Figure. 2A-B) revealed not only enrichment of ARGs but also co-localization of these ARGs with diverse mobile genetic elements (MGEs), which is a feature that links these plasmids to the broader landscape of horizontal gene transfer (HGT) mediated by integrative and conjugative elements (ICEs) or genomic islands (GIs). Both plasmids carried Tn3 family transposases, IS66/IS1182 family transposases, and ISCfr1 insertion sequences (ISs) (Figure 2A-B), which are canonical components of ICEs/GIs. For instance, blaTEM-1 (present in both plasmids) is flanked by Tn3 transposase and insertion sequence (IS) elements (Figure 2A-B): Tn3 family transposons are frequently captured by ICEs to form stable ARG-ICE composites, enabling ARG retention in chromosomes (via ICE integration) and interspecies transfer (via ICE conjugation). Notably, pBJ014-1 harbors a dense cluster of ARGs (blaMOX-3, sul1, arr-3, etc.) adjacent to multiple IS elements, while pBJ018-1 carries aac(6')-lb4 and fosC2 alongside ISPsy43. Such ARG-MGE arrays are hallmarks of resistance-associated GIs, which can cycle between the chromosomal (ICEs/GIs) and extrachromosomal (plasmid) states. This dual localization enhances their dissemination potential; ICEs/GIs facilitate cross-species HGT via conjugation, whereas plasmids drive rapid intraspecies spread within microbial communities. All four A. hydrophila strains were isolated from the same thoracic surgery department, indicating that they might have exchanged and gained ARGs from each other or from other MDR bacterial species.
In summary, in this 10-year retrospective study of 60 clinical antibiotic-resistant Aeromonas strains, we elucidated the genetic characteristics of six MDR Aeromonas spp., identified a novel CphA variant, and confirmed the presence of plasmid-mediated ARGs. Ongoing monitoring is required to curb the spread of ARGs and resistant bacteria.
doi: 10.3967/bes2026.068
Genetic Characterization of Multidrug-resistant Aeromonas Isolates from a General Hospital in China and Identification of a New CphA Variant
-
The authors declare that they received financial support for the research, authorship, and publication of this article. This work was supported by the Capital Fund for Health Improvement and Research (No. 2016-4-1101) and the Beijing Natural Science Foundation (No. 7164249).
The authors declare that there are no conflicts of interest.
Ethical approval for the study (2017-P2-095-01 and BFHHZS20250216) was obtained from and the requirement for informed consent was waived by the Medical Ethics Committee of Beijing Friendship Hospital, Capital Medical University, Beijing, China.
Conceptualization and Resources: Yanyan Zhou, Liyan Ma, and Li Yu; Data Analysis: Yanyan Zhou, Keyi Yu, and Mengyu Shi; Funding Acquisition: Yanyan Zhou and Li Yu; Investigation and Methodology: Keyi Yu, Ming Liu, Zhenzhou Huang, Zhenpeng Li, and Xiaoli Du; Project Administration and Visualization: Liyan Ma, Duochun Wang, and Li Yu; Supervision and Validation: Liyan Ma, Duochun Wang, and Li Yu; Writing–Original Draft Preparation & Editing: Yanyan Zhou, Keyi Yu, and Ming Liu.
&These authors contributed equally to this work.
注释:1) Funding: 2) Competing Interests: 3) Ethics: 4) Authors’ Contributions: -
Figure 1. (A) The antibiotic resistance genes of six clinical Aeromonas strains. Maroon rectangles represent genes located on plasmids, and blue rectangles represent genes located in chromosomes. (B) Phylogenetic analysis of CphA enzymes. (C) Amino acid alignment of CphA-18 with four other CphA variants. (D) The predicted protein structure of CphA-18. Helices are shown in teal, sheets are shown in purple, and loops are shown in pink.
Figure 2. (A & B) Maps of the complete sequences of pBJ014-1 (A) and pBJ018-1 (B). (C) The genetic structure of the class 1 integron present in pBJ014-1 and pBJ018-1.
Table 1. Antibiotic susceptibilities of 60 Aeromonas spp. isolates collected from extraintestinal infections at Beijing Friendship Hospital, the Second Affiliated Hospital of Capital Medical University, Beijing in 2014–2025.
Antibiotic Total (n = 60) 2014–2017 (n = 17) 2018–2025 (n = 43) R1 [n (%)] I [n (%)] S [n (%)] R [n (%)] I [n (%)] S [n (%)] R [n (%)] I [n (%)] S [n (%)] Imipenema 13 (21.7) 4 (6.7) 43 (71.7) 1 (5.9) 2 (11.8) 14 (82.4) 12 (27.9) 2 (4.7) 29 (67.4) Cefepimea 8 (4.3) 6 (2.6) 46 (93.0) 4 (23.5) 3 (17.6) 10 (58.8) 4 (9.3) 3 (7.0) 36 (83.7) Ciprofloxacina 13 (21.7) 0 (0.0) 47 (78.3) 6 (35.3) 0 (0.0) 11 (64.7) 7 (16.3) 0 (0.0) 36 (83.7) Trimethoprim- Sulfamethoxazolea 13 (21.7) 0 (0.0) 47 (78.3) 5 (29.4) 0 (0.0) 12 (70.6) 8 (18.6) 0 (0.0) 35 (81.4) Note. 1 R, Resistant; I, Intermediate; S, Sensitive.
a Breakpoints are based on the CLSIM45-A3 standards for Aeromonas spp.
b Other breakpoints are based on the CLSIM100- ED33 criteria for Enterobacteriaceae. 下载: 导出CSV
下载: 导出CSV
-
[1] Fernández-Bravo A, Figueras MJ. An update on the genus Aeromonas: taxonomy, epidemiology, and pathogenicity. Microorganisms, 2020; 8, 129. doi: 10.3390/microorganisms8010129 [2] Pineda-Reyes R, Neil BH, Orndorff J, et al. Clinical presentation, antimicrobial resistance, and treatment outcomes of Aeromonas human infections: a 14-year retrospective study and comparative genomics of 2 isolates from fatal cases. Clin Infect Dis, 2024; 79, 1144−52. doi: 10.1093/cid/ciae272 [3] Zhang DQ, Li WT, Hu X, et al. Requiring reconsideration of differences of Aeromonas infections between extra-intestinal and intestinal in hospitalized patients. Infect Drug Resist, 2023; 16, 487−97. doi: 10.2147/IDR.S393347. eCollection 2023 [4] Rhee JY, Jung DS, Peck KR. Clinical and therapeutic implications of Aeromonas bacteremia: 14 years nation-wide experiences in Korea. Infect Chemother, 2016; 48, 274−84. doi: 10.3947/ic.2016.48.4.274 [5] Drk S, Puljko A, Dželalija M, et al. Characterization of third generation cephalosporin- and carbapenem-resistant Aeromonas isolates from municipal and hospital wastewater. Antibiotics (Basel), 2023; 12, 513. doi: 10.3390/antibiotics12030513 [6] Zhou YY, Yu L, Nan Z, et al. Taxonomy, virulence genes and antimicrobial resistance of Aeromonas isolated from extra-intestinal and intestinal infections. BMC Infect Dis, 2019; 19, 158. doi: 10.1186/s12879-019-3766-0 [7] Kraychete GB, Botelho LAB, Monteiro-Dias PV, et al. qnrVC occurs in different genetic contexts in Klebsiella and Enterobacter strains isolated from Brazilian coastal waters. J Glob Antimicrob Resist, 2022; 31, 38−44. doi: 10.1016/j.jgar.2022.08.004 [8] Bottoni C, Perilli M, Marcoccia F, et al. Kinetic studies on CphA mutants reveal the role of the P158-P172 loop in activity versus carbapenems. Antimicrob Agents Chemother, 2016; 60, 3123−6. doi: 10.1128/AAC.01703-15 [9] Hilt EE, Fitzwater SP, Ward K, et al. Carbapenem resistant Aeromonas hydrophila carrying blacphA7 isolated from two solid organ transplant patients. Front Cell Infect Microbiol, 2020; 10, 563482. doi: 10.3389/fcimb.2020.563482 [10] Bello-López JM, Cabrero-Martínez OA, Ibáñez-Cervantes G, et al. Horizontal gene transfer and its association with antibiotic resistance in the genus Aeromonas spp. Microorganisms, 2019; 7, 363. doi: 10.3390/microorganisms7090363 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 37
- HTML全文浏览量: 13
- PDF下载量: 0
- 被引次数: 0

 Quick Links
Quick Links